
Multiple Head Rotations Result in Persistent Gait Alterations in Piglets
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Accreditation
2.3. Acclimation
2.4. Design of Animal Experiments Based on On-Field Head Impact Measurements in Soccer
2.5. Head Rotation Methodology
2.6. Gait Assessment
2.7. Data Processing
3. Statistics and Analysis
3.1. Healthy Reference Range
3.2. Experimental Groups
4. Results
4.1. Overview
4.2. Gait Parameters
4.2.1. Number of Stances
4.2.2. Gait Time
4.2.3. Gait Velocity
4.2.4. Cycle Time
4.2.5. Cycles per Minute (Cadence)
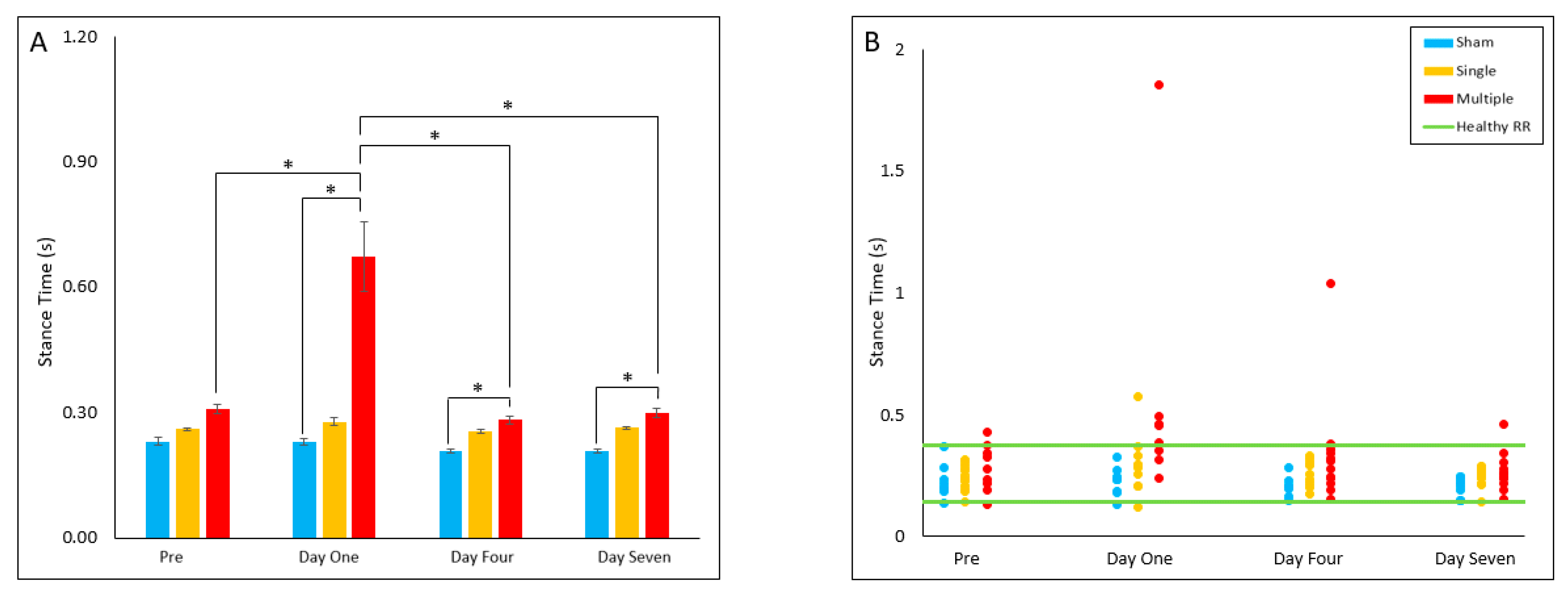
4.2.6. Stance Time
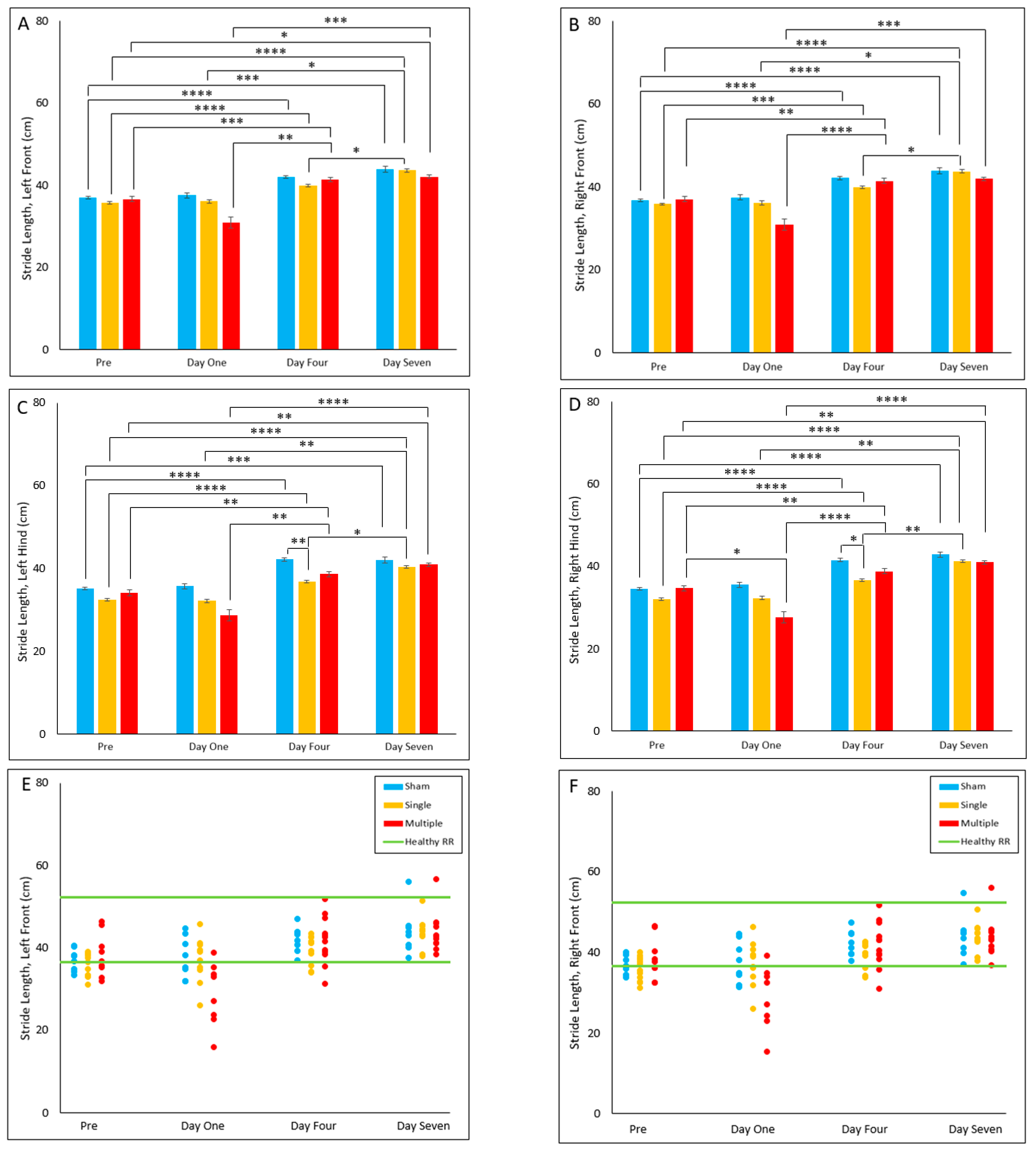
4.2.7. Stride Length
5. Discussion
5.1. General Summary
5.2. Relationship with Previous Pediatric Studies
5.3. Limitations and Future Work
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, P.; Li, R.; Schwebel, D.C.; Zhu, M.; Hu, G. Traumatic brain injury mortality among U.S. children and adolescents ages 0–19 years, 1999–2017. J. Saf. Res. 2020, 72, 93–100. [Google Scholar] [CrossRef]
- Dewan, M.C.; Mummareddy, N.; Wellons, J.C., 3rd; Bonfield, C.M. Epidemiology of Global Pediatric Traumatic Brain Injury: Qualitative Review. World Neurosurg. 2016, 91, 497–509.e1. [Google Scholar] [CrossRef]
- Schneier, A.J.; Shields, B.J.; Hostetler, S.G.; Xiang, H.; Smith, G.A. Incidence of Pediatric Traumatic Brain Injury and Associated Hospital Resource Utilization in the United States. Pediatrics 2006, 118, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Faul, M.; Xu, L.; Wald, M.M.; Coronado, V.; Dellinger, A.M. Traumatic brain injury in the United States: National estimates of prevalence and incidence, 2002–2006. Inj. Prev. 2010, 16, A268. [Google Scholar] [CrossRef]
- Prince, C.; Bruhns, M.E. Evaluation and Treatment of Mild Traumatic Brain Injury: The Role of Neuropsychology. Brain Sci. 2017, 7, 105. [Google Scholar] [CrossRef]
- Popernack, M.L.; Gray, N.; Reuter-Rice, K. Moderate-to-Severe Traumatic Brain Injury in Children: Complications and Rehabilitation Strategies. J. Pediatr. Health Care 2015, 29, e1–e7. [Google Scholar] [CrossRef]
- Kuhtz-Buschbeck, J.P.; Hoppe, B.; Gölge, M.; Dreesmann, M.; Damm-Stünitz, U.; Ritz, A. Sensorimotor recovery in children after traumatic brain injury: Analyses of gait, gross motor, and fine motor skills. Dev. Med. Child Neurol. 2003, 45, 821–828. [Google Scholar] [CrossRef]
- Schaaf, P.J.V.; Kriel, R.L.; Krach, L.; Luxenberg, M.G. Late improvements in mobility after acquired brain injuries in children. Pediatr. Neurol. 1997, 16, 306–310. [Google Scholar] [CrossRef]
- Kuhtz-Buschbeck, J.P.; Stolze, H.; Ritz, A. Analyses of gait, reaching, and grasping in children after traumatic brain injury. Arch. Phys. Med. Rehabil. 2003, 84, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Katz-Leurer, M.; Rotem, H.; Keren, O.; Meyer, S. The effect of variable gait modes on walking parameters among children post severe traumatic brain injury and typically developed controls. NeuroRehabilitation 2011, 29, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Howell, D.; Osternig, L.R.; Koester, M.C.; Chou, L.-S. The effect of cognitive task complexity on gait stability in adolescents following concussion. Exp. Brain Res. 2014, 232, 1773–1782. [Google Scholar] [CrossRef]
- Sambasivan, K.; Grilli, L.; Gagnon, I. Balance and mobility in clinically recovered children and adolescents after a mild traumatic brain injury. J. Pediatr. Rehabil. Med. 2015, 8, 335–344. [Google Scholar] [CrossRef]
- Berkner, J.; Meehan, W.P.; Master, C.L.; Howell, D.R. Gait and Quiet-Stance Performance Among Adolescents After Concussion-Symptom Resolution. J. Athl. Train. 2017, 52, 1089–1095. [Google Scholar] [CrossRef]
- Howell, D.R.; Kirkwood, M.W.; Provance, A.; Iverson, G.L.; Meehan, W.P. Using concurrent gait and cognitive assessments to identify impairments after concussion: A narrative review. Concussion 2018, 3, CNC54. [Google Scholar] [CrossRef]
- Stephens, J.; Salorio, C.; Denckla, M.; Mostofsky, S.; Suskauer, S. Subtle Motor Findings During Recovery from Pediatric Traumatic Brain Injury: A Preliminary Report. J. Mot. Behav. 2016, 49, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Lasry, O.; Liu, E.Y.; Powell, G.A.; Ruel-Laliberté, J.; Marcoux, J.; Buckeridge, D.L. Epidemiology of recurrent traumatic brain injury in the general population. Neurology 2017, 89, 2198–2209. [Google Scholar] [CrossRef]
- Sariaslan, A.; Sharp, D.J.; D’Onofrio, B.M.; Larsson, H.; Fazel, S. Long-Term Outcomes Associated with Traumatic Brain Injury in Childhood and Adolescence: A Nationwide Swedish Cohort Study of a Wide Range of Medical and Social Outcomes. PLoS Med. 2016, 13, e1002103. [Google Scholar] [CrossRef] [PubMed]
- Swaine, B.R.; Tremblay, C.; Platt, R.W.; Grimard, G.; Zhang, X.; Pless, I.B. Previous Head Injury Is a Risk Factor for Subsequent Head Injury in Children: A Longitudinal Cohort Study. Pediatrics 2007, 119, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Prins, M.; Giza, C. Repeat traumatic brain injury in the developing brain. Int. J. Dev. Neurosci. 2012, 30, 185–190. [Google Scholar] [CrossRef]
- Joyce, T.; Gossman, W.; Huecker, M.R. Pediatric Abusive Head Trauma. In Statpearls; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Hung, K.-L. Pediatric abusive head trauma. Biomed. J. 2020, 43, 240–250. [Google Scholar] [CrossRef]
- Slobounov, S.; Slobounov, E.; Sebastianelli, W.; Cao, C.; Newell, K. Differential rate of recovery in athletes after first and second concussion episodes. Neurosurgery 2007, 61, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Gaetz, D.G.M.; Goodman, D.; Weinberg, H. Electrophysiological evidence for the cumulative effects of concussion. Brain Inj. 2000, 14, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Wall, S.E.; Williams, W.H.; Cartwright-Hatton, S.; Kelly, T.P.; Murray, J.; Murray, M.; Owen, A.; Turner, M. Neuropsychological dysfunction following repeat concussions in jockeys. J. Neurol. Neurosurg. Psychiatry 2006, 77, 518–520. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.N.B.; Joseph, B.; Brelsfoard, J.M.; Saatman, K.E. Repeated Closed Head Injury in Mice Results in Sustained Motor and Memory Deficits and Chronic Cellular Changes. PLoS ONE 2016, 11, e0159442. [Google Scholar] [CrossRef]
- Mountney, A.; Boutté, A.M.; Cartagena, C.M.; Flerlage, W.F.; Johnson, W.D.; Rho, C.; Lu, X.-C.; Yarnell, A.; Marcsisin, S.; Sousa, J.; et al. Functional and Molecular Correlates after Single and Repeated Rat Closed-Head Concussion: Indices of Vulnerability after Brain Injury. J. Neurotrauma 2017, 34, 2768–2789. [Google Scholar] [CrossRef]
- Araki, T.; Yokota, H.; Morita, A. Pediatric Traumatic Brain Injury: Characteristic Features, Diagnosis, and Management. Neurol. Med. Chir. 2017, 57, 82–93. [Google Scholar] [CrossRef]
- Coats, B.; Margulies, S.S. Material Properties of Human Infant Skull and Suture at High Rates. J. Neurotrauma 2006, 23, 1222–1232. [Google Scholar] [CrossRef]
- Duhaime, A.-C.; Margulies, S.S.; Durham, S.R.; O’Rourke, M.M.; Golden, J.A.; Marwaha, S.; Raghupathi, R. Maturation-dependent response of the piglet brain to scaled cortical impact. J. Neurosurg. 2000, 93, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Duberstein, K.J.; Platt, S.R.; Holmes, S.P.; Dove, C.R.; Howerth, E.W.; Kent, M.; Stice, S.L.; Hill, W.D.; Hess, D.C.; West, F.D. Gait analysis in a pre- and post-ischemic stroke biomedical pig model. Physiol. Behav. 2014, 125, 8–16. [Google Scholar] [CrossRef]
- Xiong, Y.; Mahmood, A.; Chopp, M. Animal models of traumatic brain injury. Nat. Rev. Neurosci. 2013, 14, 128–142. [Google Scholar] [CrossRef]
- Povlishock, J.T.; Hayes, R.L.; Michel, M.E.; McIntosh, T.K. Workshop on Animal Models of Traumatic Brain Injury. J. Neurotrauma 1994, 11, 723–732. [Google Scholar] [CrossRef]
- Grate, L.L.; A Golden, J.; Hoopes, P.; Hunter, J.V.; Duhaime, A.-C. Traumatic brain injury in piglets of different ages: Techniques for lesion analysis using histology and magnetic resonance imaging. J. Neurosci. Methods 2003, 123, 201–206. [Google Scholar] [CrossRef]
- Lind, N.M.; Moustgaard, A.; Jelsing, J.; Vajta, G.; Cumming, P.; Hansen, A.K. The use of pigs in neuroscience: Modeling brain disorders. Neurosci. Biobehav. Rev. 2007, 31, 728–751. [Google Scholar] [CrossRef] [PubMed]
- Duhaime, A.-C. Large Animal Models of Traumatic Injury to the Immature Brain. Dev. Neurosci. 2006, 28, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Kinder, H.; Baker, E.W.; Wang, S.; Fleischer, C.C.; Howerth, E.W.; Duberstein, K.J.; Mao, H.; Platt, S.R.; West, F.D. Traumatic Brain Injury Results in Dynamic Brain Structure Changes Leading to Acute and Chronic Motor Function Deficits in a Pediatric Piglet Model. J. Neurotrauma 2019, 36, 2930–2942. [Google Scholar] [CrossRef]
- Kinder, H.A.; Baker, E.W.; West, F.D. The pig as a preclinical traumatic brain injury model: Current models, functional outcome measures, and translational detection strategies. Neural Regen. Res. 2019, 14, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Baker, E.W.; Mandal, A.; Pidaparti, R.M.; West, F.D.; Kinder, H.A. Identification of predictive MRI and functional biomarkers in a pediatric piglet traumatic brain injury model. Neural Regen. Res. 2021, 16, 338–344. [Google Scholar] [CrossRef]
- Dickerson, J.W.T.; Dobbing, J. Prenatal and postnatal growth and development of the central nervous system of the pig. Proc. R. Soc. London. Ser. B Boil. Sci. 1967, 166, 384–395. [Google Scholar] [CrossRef]
- Ryan, M.C.; Sherman, P.; Rowland, L.; Wijtenburg, S.A.; Acheson, A.; Fieremans, E.; Veraart, J.; Novikov, D.; Hong, L.E.; Sladky, J.; et al. Miniature pig model of human adolescent brain white matter development. J. Neurosci. Methods 2017, 296, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Margulies, S.S.; Coats, B. Biomechanics of pediatric TBI. In Pediatric Traumatic Brain Injury: New Frontiers in Clinical and Translational Research; Yeates, K.O., Anderson, V., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 7–17. [Google Scholar]
- Burrows, P.; Trefan, L.; Houston, R.; Hughes, J.; Pearson, G.; Edwards, R.J.; Hyde, P.; Maconochie, I.; Parslow, R.C.; Kemp, A.M. Head injury from falls in children younger than 6 years of age. Arch. Dis. Child. 2015, 100, 1032–1037. [Google Scholar] [CrossRef]
- Cullen, D.K.; Harris, J.P.; Browne, K.D.; Wolf, J.A.; Duda, J.E.; Meaney, D.F.; Margulies, S.S.; Smith, D.H. A Porcine Model of Traumatic Brain Injury via Head Rotational Acceleration. Methods Mol. Biol. 2016, 1462, 289–324. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Wang, Y.; Kim, S.; Hong, S.M.; Jeng, L.; Bilgen, M.; Liu, J. Assessing gait impairment following experimental traumatic brain injury in mice. J. Neurosci. Methods 2009, 176, 34–44. [Google Scholar] [CrossRef]
- Luo, J.; Nguyen, A.; Villeda, S.; Zhang, H.; Ding, Z.; Lindsey, D.; Bieri, G.; Castellano, J.M.; Beaupre, G.S.; Ewyss-Coray, T. Long-Term Cognitive Impairments and Pathological Alterations in a Mouse Model of Repetitive Mild Traumatic Brain Injury. Front. Neurol. 2014, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.; Grillakis, A.; Kline, A.; Ahmed, A.E.; Byrnes, K.R. Gait analysis in a rat model of traumatic brain injury. Behav. Brain Res. 2021, 405, 113210. [Google Scholar] [CrossRef] [PubMed]
- Abstracts from The 38th Annual National Neurotrauma Symposium July 11–14, 2021 Virtual Conference. J. Neurotrauma 2021, 38, A125. [CrossRef]
- Hajiaghamemar, M.; Seidi, M.; Margulies, S.S. Head Rotational Kinematics, Tissue Deformations, and Their Relationships to the Acute Traumatic Axonal Injury. J. Biomech. Eng. 2020, 142, 0310061–03100613. [Google Scholar] [CrossRef]
- Hajiaghamemar, M.; Seidi, M.; Patton, D.; Huber, C.; Arbogast, K.B.; Master, C.; Margulies, S.S. Using On-Field Human Head Kinematics to Guide Study Design for Animal-Model Based Traumatic Brain Injury. In Proceedings of the Biomedical Engineering Society Annual Meeting, Philadelphia, PA, USA, 16–19 October 2020. [Google Scholar]
- Gennarelli, T.A.; Thibault, L.E.; Graham, D.I. Diffuse Axonal Injury: An Important Form of Traumatic Brain Damage. Neurosci. 1998, 4, 202–215. [Google Scholar] [CrossRef]
- Ommaya, A.K.; Gennarelli, T.A. Cerebral Concussion and Traumatic Unconsciousness: Correlation of Experimental and Clinical Observations on Blunt Head Injuries. Brain 1974, 97, 633–654. [Google Scholar] [CrossRef]
- Wu, T.; Antona-Makoshi, J.; Alshareef, A.; Giudice, S.; Panzer, M.B. Investigation of Cross-Species Scaling Methods for Traumatic Brain Injury Using Finite Element Analysis. J. Neurotrauma 2020, 37, 410–422. [Google Scholar] [CrossRef]
- Hajiaghamemar, M.; Margulies, S.S. Traumatic Brain Injury: Translating Head Kinematics Outcomes between Pig and Human; International Research Council on Biomechanics of Injury: Zurich, Switzerland, 2020; pp. 605–607. [Google Scholar]
- Patton, D.A.; Huber, C.M.; Douglas, E.C.; Seacrist, T.; Arbogast, K.B. Laboratory assessment of a head impact sensor for youth soccer ball heading impacts using an anthropomorphic test device. Proc. Inst. Mech. Eng. Part P J. Sports Eng. Technol. 2021, 17543371211063124. [Google Scholar] [CrossRef]
- Pusparum, M.; Ertaylan, G.; Thas, O. From Population to Subject-Specific Reference Intervals. In Proceedings of the International Conference on Computational Science—ICCS 2020, Amsterdam, The Netherlands, 3–5 June 2020; Krzhizhanovskaya, V.V., Závodszky, G., Lees, M.H., Dongarra, J.J., Sloot, P.M.A., Brissos, S., Teixeira, J., Eds.; Springer: New York, NY, USA, 2020; pp. 468–482. [Google Scholar] [CrossRef]
- Karunakaran, K.K.; Ehrenberg, N.; Cheng, J.; Bentley, K.; Nolan, K.J. Kinetic Gait Changes after Robotic Exoskeleton Training in Adolescents and Young Adults with Acquired Brain Injury. Appl. Bionics Biomech. 2020, 2020, e8845772. [Google Scholar] [CrossRef]
- Gagnon, I.; Swaine, B.; Friedman, D.; Forget, R. Children show decreased dynamic balance after mild traumatic brain injury. Arch. Phys. Med. Rehabil. 2004, 85, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Katz-Leurer, M.; Rotem, H.; Keren, O.; Meyer, S. Effect of concurrent cognitive tasks on gait features among children post-severe traumatic brain injury and typically-developed controls. Brain Inj. 2011, 25, 581–586. [Google Scholar] [CrossRef]
- Katz-Leurer, M.; Rotem, H.; Lewitus, H.; Keren, O.; Meyer, S. Relationship between balance abilities and gait characteristics in children with post-traumatic brain injury. Brain Inj. 2008, 22, 153–159. [Google Scholar] [CrossRef]
- Maki, B.E. Gait Changes in Older Adults: Predictors of Falls or Indicators of Fear? J. Am. Geriatr. Soc. 1997, 45, 313–320. [Google Scholar] [CrossRef]
- Fukuchi, C.A.; Fukuchi, R.K.; Duarte, M. Effects of walking speed on gait biomechanics in healthy participants: A systematic review and meta-analysis. Syst. Rev. 2019, 8, 153. [Google Scholar] [CrossRef]
- Fettrow, T.; Reimann, H.; Grenet, D.; Crenshaw, J.; Higginson, J.; Jeka, J. Walking Cadence Affects the Recruitment of the Medial-Lateral Balance Mechanisms. Front. Sports Act. Living 2019, 1, 40. [Google Scholar] [CrossRef] [PubMed]
- Katz-Leurer, M.; Rotem, H.; Keren, O.; Meyer, S. Balance abilities and gait characteristics in post-traumatic brain injury, cerebral palsy and typically developed children. Dev. Neurorehabilit. 2009, 12, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Katz-Leurer, M.; Rotem, H.; Keren, O.; Meyer, S. The relationship between step variability, muscle strength and functional walking performance in children with post-traumatic brain injury. Gait Posture 2009, 29, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Rahman, R.A.A.; Hanapiah, F.A.; Nikmat, A.W.; Ismail, N.A.; Manaf, H. Effects of Concurrent Tasks on Gait Performance in Children with Traumatic Brain Injury Versus Children With Typical Development. Ann. Rehabil. Med. 2021, 45, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Beretta, E.; Cimolin, V.; Picciinini, L.; Turconi, A.C.; Galbiati, S.; Crivellini, M.; Galli, M.; Strazzer, S. Assessment of gait recovery in children after traumatic brain injury. Brain Inj. 2009, 23, 751–759. [Google Scholar] [CrossRef]
- Howell, D.R.; Beasley, M.; Vopat, L.; Meehan, W.P. The Effect of Prior Concussion History on Dual-Task Gait following a Concussion. J. Neurotrauma 2017, 34, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Rahman, R.A.A.; Rafi, F.; Hanapiah, F.A.; Nikmat, A.W.; Ismail, N.A.; Manaf, H. Effect of Dual-Task Conditions on Gait Performance during Timed Up and Go Test in Children with Traumatic Brain Injury. Rehabil. Res. Pr. 2018, 2018, e2071726. [Google Scholar] [CrossRef]
- Coats, B.; Binenbaum, G.; Smith, C.; Peiffer, R.L.; Christian, C.W.; Duhaime, A.-C.; Margulies, S.S. Cyclic Head Rotations Produce Modest Brain Injury in Infant Piglets. J. Neurotrauma 2017, 34, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Prange, M.T.; Coats, B.; Duhaime, A.-C.; Margulies, S.S. Anthropomorphic simulations of falls, shakes, and inflicted impacts in infants. J. Neurosurg. 2003, 99, 143–150. [Google Scholar] [CrossRef]
- Ommaya, A.; Hirsch, A. Tolerances for cerebral concussion from head impact and whiplash in primates. J. Biomech. 1971, 4, 13–21. [Google Scholar] [CrossRef]
- Gupte, R.P.; Brooks, W.; Vukas, R.; Pierce, J.D.; Harris, J.L. Sex Differences in Traumatic Brain Injury: What We Know and What We Should Know. J. Neurotrauma 2019, 36, 3063–3091. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rotation Type | Angular Velocity (rad/s) | Angular Acceleration (rad/s²) | |
|---|---|---|---|
| Single | High | 104.5 ± 0.47 | 40,052 ± 1559 |
| Multiple | Median | 61.3 ± 0.18 | 15,010 ± 169 |
| High | 104.6 ± 0.41 | 38,368 ± 499 |
| Parameter Type | Parameter | Definition |
|---|---|---|
| Single-Value Parameters | Number of Stances | Also known as ‘Number of Strikes’; how many total stances an animal makes during trial; stances in quadrupeds can involve 2- or 3-legged support |
| Gait Time | Time, in seconds, that it takes for animal to cross the gait mat; begins with contact of the first left or right front stance and ends with the time of contact of the last left or right front stance registered on the sensor | |
| Gait Velocity | Gait distance divided by gait time; centimeters per second | |
| Cycle Time | Average time, in seconds, to complete a gait cycle | |
| Cycles Per Minute | Also known as “cadence”; number of complete gait cycles per minute | |
| Individual Hoof Parameters (left front, right front, left hind, right hind) | Swing Time | Elapsed time between the last contact of a preceding hoof and first contact of the next step of that same hoof, in seconds |
| Stride Time | Elapsed time between the first contacts of two consecutive hoof falls, in seconds | |
| Stance Time | Average time from first contact to last contact of each hoof, in seconds | |
| Stride Length | Distance measured parallel to the line of progression, between the posterior heel points of two consecutive hoof falls, in centimeters | |
| Stride Velocity | Stride length divided by stride time for each hoof; centimeters per second |
| Pre-Injury | Day Seven | p-Value | |
|---|---|---|---|
| Sham | 6.34 ± 0.601 | 9.20 ± 0.728 | <0.001 |
| Single | 6.60 ± 0.221 | 8.90 ± 0.444 | <0.001 |
| Multiple | 7.11 ± 0.556 | 9.33 ± 0.625 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mull, M.; Aderibigbe, O.; Hajiaghamemar, M.; Oeur, R.A.; Margulies, S.S. Multiple Head Rotations Result in Persistent Gait Alterations in Piglets. Biomedicines 2022, 10, 2976. https://doi.org/10.3390/biomedicines10112976
Mull M, Aderibigbe O, Hajiaghamemar M, Oeur RA, Margulies SS. Multiple Head Rotations Result in Persistent Gait Alterations in Piglets. Biomedicines. 2022; 10(11):2976. https://doi.org/10.3390/biomedicines10112976
Chicago/Turabian StyleMull, Mackenzie, Oluwagbemisola Aderibigbe, Marzieh Hajiaghamemar, R. Anna Oeur, and Susan S Margulies. 2022. "Multiple Head Rotations Result in Persistent Gait Alterations in Piglets" Biomedicines 10, no. 11: 2976. https://doi.org/10.3390/biomedicines10112976
APA StyleMull, M., Aderibigbe, O., Hajiaghamemar, M., Oeur, R. A., & Margulies, S. S. (2022). Multiple Head Rotations Result in Persistent Gait Alterations in Piglets. Biomedicines, 10(11), 2976. https://doi.org/10.3390/biomedicines10112976