
Differences in the Pro/Antioxidative Status and Cellular Stress Response in Elderly Women after 6 Weeks of Exercise Training Supported by 1000 mg of Vitamin C Supplementation

 ,
, 
 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Diet Analysis
2.3. Training
2.4. Supplementation
2.5. Determination of Maximum Oxygen Uptake
2.6. RNA Isolation
2.7. Reverse Transcription and qRT-PCR
2.8. Assessment of Plasma Vitamin C Level
2.9. Biochemical Analysis
2.10. Statistical Analysis
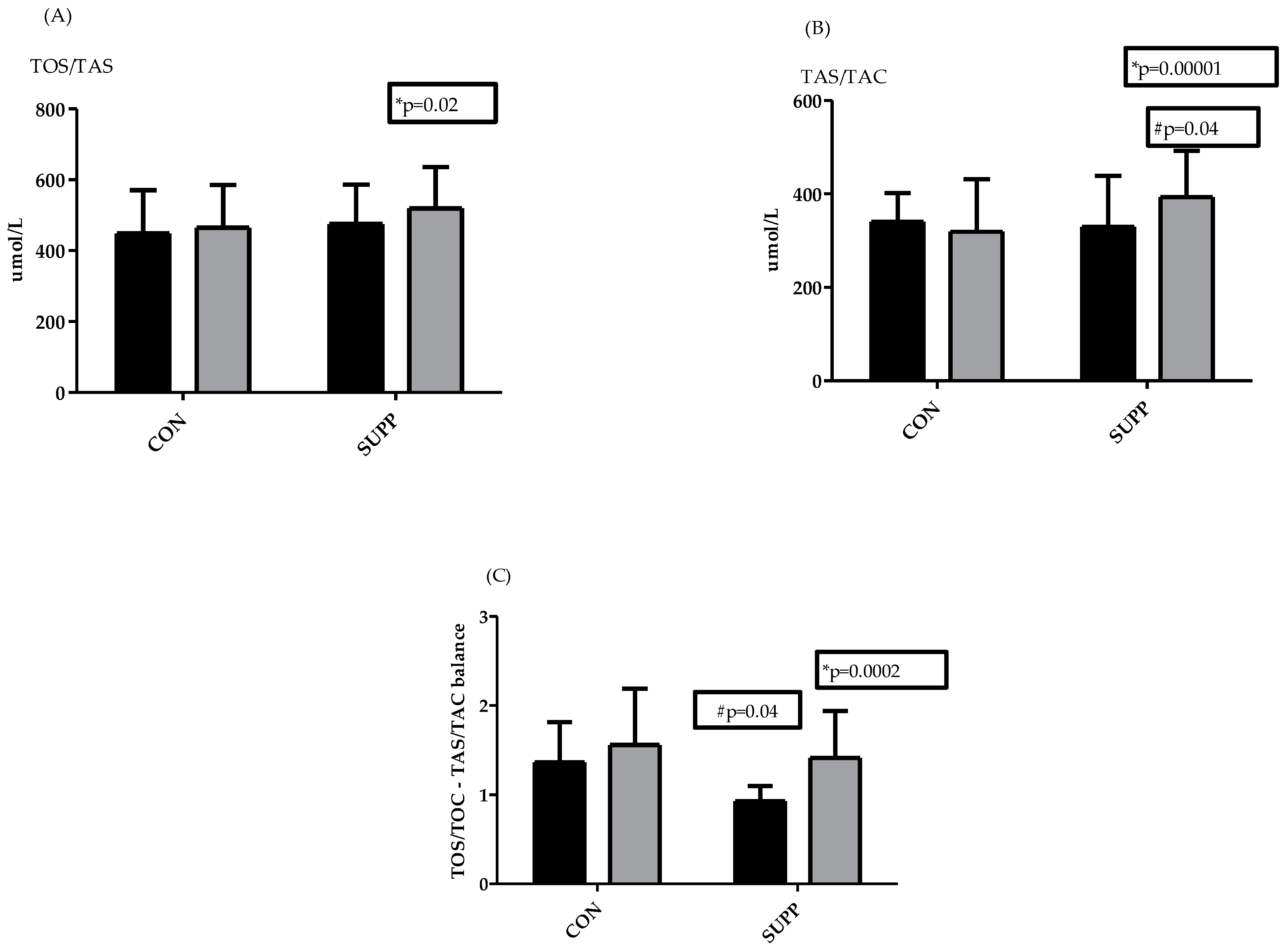
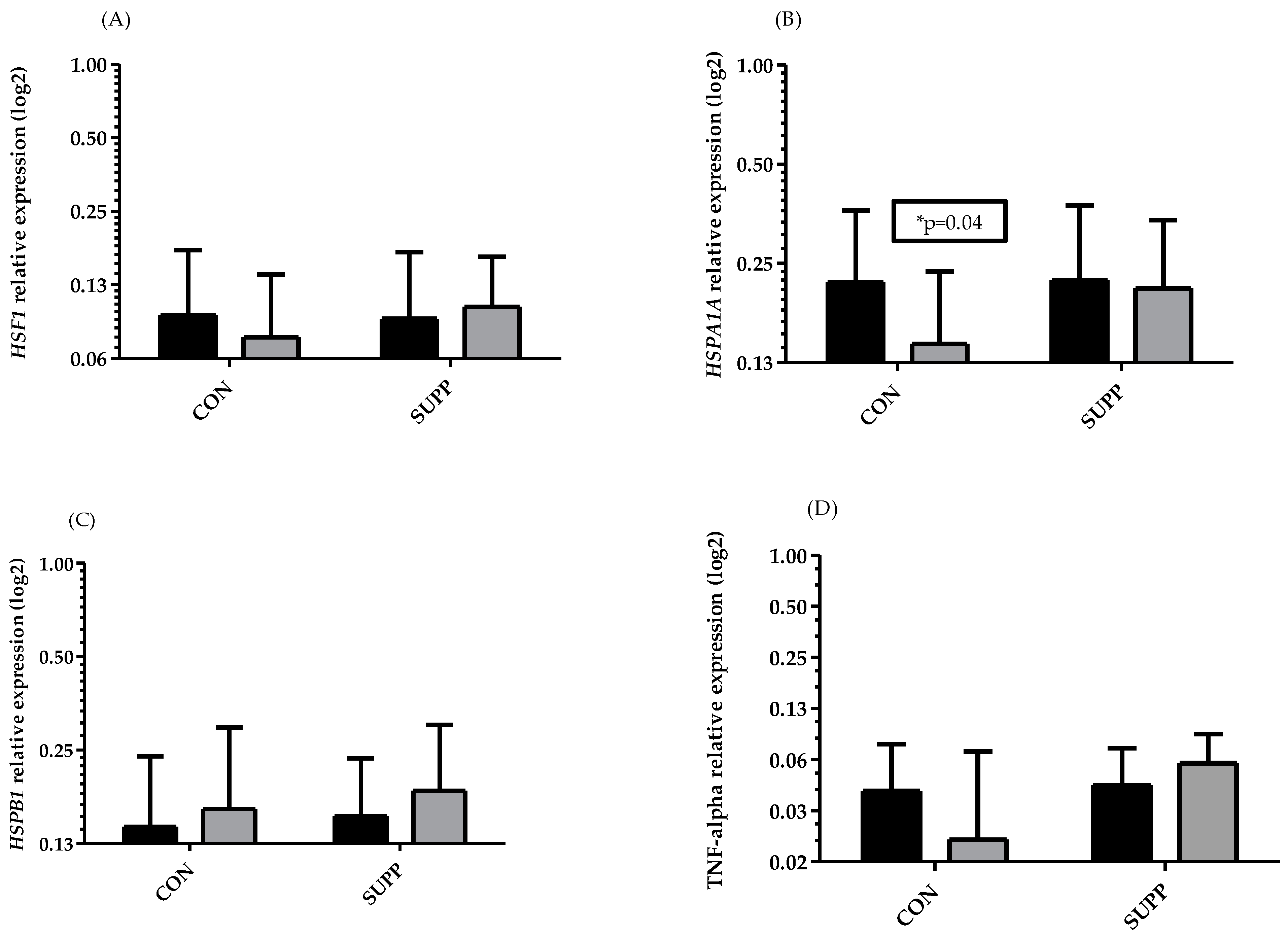
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balsano, C.; Alisi, A. Antioxidant Effects of Natural Bioactive Compounds. Curr. Pharm. Des. 2009, 15, 3063–3073. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Wan, R.; Woo, H.; Cheung, Y.; Fong, C.; Cheung, C.; Lam, B.; Janus, E.; Woo, J.; Lam, K.S.L. Dietary Intake of Anti-Oxidant Vitamins A, C, and E Is Inversely Associated with Adverse Cardiovascular Prospective Study. Nutrients 2018, 10, 1664. [Google Scholar] [CrossRef] [PubMed]
- Gaman, A.M.; Buga, A.; Gaman, M.; Popa-wagner, A. The Role of Oxidative Stress and the Effects of Antioxidants on the Incidence of Infectious Complications of Chronic Lymphocytic Leukemia. Oxid. Med. Cell. Longev. 2014, 2014, 158135. [Google Scholar] [CrossRef]
- Tsuma-kaneko, M.; Sawanobori, M.; Kawakami, S.; Uno, T. Iron removal enhances vitamin C-induced apoptosis and growth inhibition of K-562 leukemic cells. Sci. Rep. 2018, 8, 17377. [Google Scholar] [CrossRef]
- Żychowska, M.; Jastrzębski, Z.; Chruściński, G.; Michałowska-Sawczyn, M.; Nowak-Zalewska, M. Vitamin C, A and E supplementation decreases the expression of HSPA1A and HSPB1 genes in the leukocytes of young polish figure skaters during a 10-day training camp. J. Int. Soc. Sports Nutr. 2015, 12, 9. [Google Scholar] [CrossRef]
- Taylor, P.; Frei, B.; Birlouez-aragon, I.; Lykkesfeldt, J.; Frei, B.; Birlouez-aragon, I.; Lykkesfeldt, J. Authors’ Perspective: What is the Optimum Intake of Vitamin C in Humans? Authors’ Perspective: What is the Optimum Intake of Vitamin C in Humans? Crit. Rev. Food Sci. Nutr. 2012, 52, 815–829. [Google Scholar] [CrossRef]
- Żychowska, M.; Grzybkowska, A.; Wiech, M.; Urbański, R.; Pilch, W.; Piotrowska, A.; Czerwińska-Ledwig, O.; Antosiewicz, J. Exercise training and vitamin c supplementation affects ferritin mrna in leukocytes without affecting prooxidative/antioxidative balance in elderly women. Int. J. Mol. Sci. 2020, 21, 6469. [Google Scholar] [CrossRef]
- Stadtman, R. Ascorbic acid and oxidative inactivation of proteines. Am. J. Clin. Nutr. 1991, 54, 125S–128S. [Google Scholar] [CrossRef]
- Żychowska, M.; Grzybkowska, A.; Zasada, M.; Piotrowska, A.; Dworakowska, D.; Czerwińska-Ledwig, O.; Pilch, W.; Antosiewicz, J. Effect of six weeks 1000 mg/day vitamin C supplementation and healthy training in elderly women on genes expression associated with the immune response—A randomized controlled trial. J. Int. Soc. Sports Nutr. 2021, 18, 19. [Google Scholar] [CrossRef]
- Pilch, W.; Piotrowska, A.; Wyrostek, J.; Czerwińska-Ledwig, O.; Ziemann, E.; Antosiewicz, J.; Zasada, M.; Kulesa-Mrowiecka, M.; Żychowska, M. Different changes in adipokines, lipid profile, and TNF-αlpha levels between 10 and 20 whole body cryostimulation sessions in individuals with I and II degrees of obesity. Biomedicines 2022, 10, 269. [Google Scholar] [CrossRef]
- Ziemann, E.; Zembro, A.; Kasperska, A.; Antosiewicz, J. Exercise Training-Induced Changes in Inflammatory Mediators and Heat Shock Proteins in Young Tennis Players. J. Sports Sci. Med. 2013, 12, 282–289. [Google Scholar]
- Mankowski, R.; Anton, S.; Buford, T.; Leeuwenburgh, C. Dietary Antioxidants as Modifiers of Physiologic Adaptations to Exercise. Med. Sci. Sports Exerc. 2015, 47, 1857–1868. [Google Scholar] [CrossRef] [PubMed]
- Bjørnsen, T.; Salvesen, S.; Berntsen, S.; Hetlelid, K.J.; Stea, T.H.; Lohne-Seiler, H.; Rohde, G.; Haraldstad, K.; Raastad, T.; Køpp, U.; et al. Vitamin C and E supplementation blunts increases in total lean body mass in elderly men after strength training. Scand. J. Med. Sci. Sports 2016, 26, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Close, G.L.; Ashton, T.; Cable, T.; Doran, D.; Holloway, C.; McArdle, F.; MacLaren, D.P.M.; Bjørnsen, T.; Salvesen, S.; Berntsen, S.; et al. Ascorbic acid supplementation does not attenuate post-exercise muscle soreness following muscle-damaging exercise but may delay the recovery process. Br. J. Nutr. 2006, 1, 976–981. [Google Scholar] [CrossRef]
- Gomez-Cabrera, M.C.; Domenech, E.; Romagnoli, M.; Arduini, A.; Borras, C.; Pallardo, F.V.; Sastre, J.; Viña, J. Oral administration of vitamin C decreases muscle mitochondrial biogenesis and hampers training-induced adaptations in endurance performance. Am. J. Clin. Nutr. 2008, 87, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Dutra, M.T.; Martins, W.R.; Ribeiro, A.L.A.; Bottaro, M. The Effects of Strength Training Combined with Vitamin C and E Supplementation on Skeletal Muscle Mass and Strength: A Systematic Review and Meta-Analysis. J. Sports Med. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Bryant, R.J.; Ryder, J.; Martino, P.; Kim, J.; Craig, B.W. Effects of vitamin E and C supplementation either alone or in combination on exercise-induced lipid peroxidation in trained cyclists. J. Strength Cond. Res. 2003, 17, 792–800. [Google Scholar] [CrossRef]
- Lira, V.A.; Benton, C.R.; Yan, Z.; Bonen, A. PGC-1α regulation by exercise training and its influences on muscle function and insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E145–E161. [Google Scholar] [CrossRef] [PubMed]
- Clifford, T.; Jeffries, O.; Stevenson, E.J.; Davies, K.A.B. The effects of vitamin C and E on exercise-induced physiological adaptations: A systematic review and Meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 3669–3679. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.; Moon, Y.; Sohn, S.; Jang, I. Effects of Cyclic Heat Stress or Vitamin C Supplementation during Cyclic Heat Stress on HSP70, Inflammatory Cytokines, and the Antioxidant Defense System in Sprague Dawley Rats. Exp. Anim. 2012, 61, 543–553. [Google Scholar] [CrossRef]
- Janus, P.; Pakuła-Cis, M.; Kalinowska-Herok, M.; Kashchak, N.; Szołtysek, K.; Pigłowski, W.; Widlak, W.; Kimmel, M.; Widlak, P. NF-κB signaling pathway is inhibited by heat shock independently of active transcription factor HSF1 and increased levels of inducible heat shock proteins. Genes Cells 2011, 16, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Bae, S.; Kim, Y.; Cho, C.; Kim, S.J.; Kim, Y.; Lee, S.; Kim, H.; Hwang, Y.; Kang, J.S.; et al. Vitamin C prevents stress-induced damage on the heart caused by the death of cardiomyocytes, through down-regulation of the excessive production of catecholamine, TNF-α, and ROS production in Gulo (−/−)Vit C-Insufficient mice. Free Radic. Biol. Med. 2013, 65, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Gamaley, I.; Klyubin, I. Roles of Reactive Oxygen Species: Signaling and Regulation of Cellular Functions. Int. Rev. Cytol. 1999, 188, 203–255. [Google Scholar] [CrossRef]
- Arya, R.; Mallik, M.; Lakhotia, S. Heat shock genes—Integrating cell survival and death. J. Biosci. 2007, 32, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lormes, W.; Baur, C.; Altenburg, D.; Lehmann, M.; Steinacker, J.M. Human Skeletal Muscle HSP70 Response to Physical Training Depends on Exercise Intensity. Int. J. Sports Med. 2000, 21, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, G.M.; Kaur, P.; Asea, A. Role of Human and Mouse HspB1 in Metastasis. Curr. Mol. Med. 2012, 12, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Fehrenbach, E. Multifarious microarray-based gene expression patterns in response to exercise. J. Appl. Physiol. 1985 2007, 102, 7–8. [Google Scholar] [CrossRef]
- Acunzo, J.; Katsogiannou, M.; Rocchi, P. The International Journal of Biochemistry Small heat shock proteins HSP27 (HspB1), B-crystallin (HspB5) and HSP22 (HspB8) as regulators of cell death. Int. J. Biochem. Cell Biol. 2012, 44, 1622–1631. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, M. Normy Żywienia Dla Populacji Polski; Instytut Żywności i Żywienia: Warszawa, Poland, 2017; ISBN 9788386060894.
- Szponar, L.; Wolnicka, K.; Rychlik, E. Album Fotografii Produktów I Potraw; Prace IŻŻ: Loreley, Germany, 2000; ISBN 83-86060-51-4.
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Robitaille, L.; Hoffer, L.J. A simple method for plasma total vitamin C analysis suitable for routine clinical laboratory use. Nutr. J. 2016, 15, 1–9. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Duarte, T.L.; Lunec, J. ReviewPart of the Series: From Dietary Antioxidants to Regulators in Cellular Signalling and Gene ExpressionReview: When is an antioxidant not an antioxidant? A review of novel actions and reactions of vitamin C. Free Radic. Res. 2005, 39, 671–686. [Google Scholar] [CrossRef] [PubMed]
- Aguiló, A.; Tauler, P.; Fuentespina, E.; Villa, G.; Córdova, A.; Tur, J.A.; Pons, A. Blood Iron Status in Endurance Athletes. Int. J. Sport Nutr. Exerc. Metab. 2004, 14, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Barańska, J.; Boguszewska, K.; Adamus-Grabicka, A.; Karwowski, B.T. Two Faces of Vitamin C—Antioxidative and Pro-Oxidative Agent. Nutrients 2020, 12, 1501. [Google Scholar] [CrossRef] [PubMed]
- Poljšak, B.; Gazdag, Z.; Jenko-Brinovec, Š.; Fujs, Š.; Pesti, M.; Bélagyi, J.; Plesničar, S.; Raspor, P. Pro-oxidative vs antioxidative properties of ascorbic acid in chromium(VI)-induced damage: Anin vivo andin vitro approach. J. Appl. Toxicol. 2005, 25, 535–548. [Google Scholar] [CrossRef]
- Bhat, S.H.; Azmi, A.S.; Hanif, S.; Hadi, S.M. Ascorbic acid mobilizes endogenous copper in human peripheral lymphocytes leading to oxidative DNA breakage: A putative mechanism for anticancer properties. Int. J. Biochem. Cell Biol. 2006, 38, 2074–2081. [Google Scholar] [CrossRef] [PubMed]
- Zoppi, C.C.; Hohl, R.; Silva, F.C.; Lazarim, F.L.; Antunes, J.M.F.; Stancanneli, M.; Macedo, D.V. Vitamin C and E Supplementation Effects in Professional Soccer Players Under Regular Training. J. Int. Soc. Sports Nutr. 2006, 3, 37–44. [Google Scholar] [CrossRef]
- Morton, J.P.; Kayani, A.C.; Mcardle, A.; Drust, B. The Exercise-Induced Stress Response of Skeletal Muscle, with Specific Emphasis on Humans. Sports Med. 2009, 39, 643–662. [Google Scholar] [CrossRef] [PubMed]
- Simar, D.; Malatesta, D.; Mas, E.; Delage, M.; Caillaud, C. Effect of an 8-weeks aerobic training program in elderly on oxidative stress and hsp72 expression in leukocytes. J. Nutr. Health Aging 2012, 16, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Cumming, K.T.; Raastad, T.; Sørstrøm, A.; Paronetto, M.P.; Mercatelli, N.; Ugelstad, I.; Caporossi, D. Vitamin C and E supplementation does not affect heat shock proteins or endogenous antioxidants in trained skeletal muscles during 12 weeks of strength training. BMC Nutr. 2017, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bunpo, P.; Anthony, T. Ascorbic acid supplementation does not alter oxidative stress markers in healthy volunteers engaged in a supervised exercise program. Appl. Physiol. Nutr. Metab. 2016, 41, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Tauler, P.; Aguiló, A.; Gimeno, I.; Fuentespina, E.; Tur, J.A.; Pons, A. Response of blood cell antioxidant enzyme defences to antioxidant diet supplementation and to intense exercise. Eur. J. Nutr. 2006, 45, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.M.W.; Pedersen, B.K. The anti-inflammatory effect of exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.S.; Beezhold, B.L.; Mostow, B.; Swan, P.D. Plasma Vitamin C Is Inversely Related to Body Mass Index and Waist Circumference but Not to Plasma Adiponectin in Nonsmoking Adults. J. Nutr. 2007, 137, 1757–1762. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Parameter | SUPP Group (n = 18) | dCohen | CON Group (n = 18) | dCohen | 2-Way ANOVA | ||||
|---|---|---|---|---|---|---|---|---|---|
| Baseline (x ± SD) | After (x ± SD) | Change (x ± SD) | Baseline (x ± SD) | After (x ± SD) | Change (x ± SD) | ||||
| Body mass (kg) | 71.1 ± 9.0 | 70.4 ± 10.4 | −0.7 ± 2.9 | 0.02 | 71.3 ± 0.4 | 68.8 * ± 10.5 | −2.4 ± 3.9 | 0.15 | # row factor (p < 0.0001) |
| VO2 max. (mL/kg/min) | 20.6.± 3.6 | 21.4 ± 3.9 | 0.9 ± 1.4 | 0.19 | 21.3 ± 3.8 | 21.4 ± 3.8 | 0.1 ± 0.75 | 0.00 | # interaction (p = 0.0006) # row factor (p < 0.0001) |
| Vit. C concent. (μmol/L) | 14.0 ± 3.9 | 18.0 * ± 7.8 | 3.9 ± 6.4 ↑ * | 0.44 | 14.5 ± 7.1 | 14.7 ± 3.9 | 0.2 ± 5.7 | 0.08 | # row factor (p < 0.0001) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żychowska, M.; Sadowska-Krępa, E.; Damiani, E.; Tiano, L.; Ziemann, E.; Nowak-Zaleska, A.; Lipińska, P.; Piotrowska, A.; Czerwińska-Ledwig, O.; Pilch, W.; et al. Differences in the Pro/Antioxidative Status and Cellular Stress Response in Elderly Women after 6 Weeks of Exercise Training Supported by 1000 mg of Vitamin C Supplementation. Biomedicines 2022, 10, 2641. https://doi.org/10.3390/biomedicines10102641
Żychowska M, Sadowska-Krępa E, Damiani E, Tiano L, Ziemann E, Nowak-Zaleska A, Lipińska P, Piotrowska A, Czerwińska-Ledwig O, Pilch W, et al. Differences in the Pro/Antioxidative Status and Cellular Stress Response in Elderly Women after 6 Weeks of Exercise Training Supported by 1000 mg of Vitamin C Supplementation. Biomedicines. 2022; 10(10):2641. https://doi.org/10.3390/biomedicines10102641
Chicago/Turabian StyleŻychowska, Małgorzata, Ewa Sadowska-Krępa, Elisabetta Damiani, Luca Tiano, Ewa Ziemann, Alicja Nowak-Zaleska, Patrycja Lipińska, Anna Piotrowska, Olga Czerwińska-Ledwig, Wanda Pilch, and et al. 2022. "Differences in the Pro/Antioxidative Status and Cellular Stress Response in Elderly Women after 6 Weeks of Exercise Training Supported by 1000 mg of Vitamin C Supplementation" Biomedicines 10, no. 10: 2641. https://doi.org/10.3390/biomedicines10102641
APA StyleŻychowska, M., Sadowska-Krępa, E., Damiani, E., Tiano, L., Ziemann, E., Nowak-Zaleska, A., Lipińska, P., Piotrowska, A., Czerwińska-Ledwig, O., Pilch, W., & Antosiewicz, J. (2022). Differences in the Pro/Antioxidative Status and Cellular Stress Response in Elderly Women after 6 Weeks of Exercise Training Supported by 1000 mg of Vitamin C Supplementation. Biomedicines, 10(10), 2641. https://doi.org/10.3390/biomedicines10102641