
Pathophysiological Mechanisms in Non-Alcoholic Fatty Liver Disease: From Drivers to Targets

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Molecular Mechanisms of Pathogenesis
2.1. Lipid Metabolism
2.1.1. Fatty Acids
2.1.2. Compound Lipids
2.1.3. Cholesterol
2.1.4. Evidence from Patients
2.2. Carbohydrates Metabolism Disruption
2.2.1. Glucose Metabolism
2.2.2. Fructose Metabolism
2.3. Immunologic System Distrubances
2.3.1. Triggers of Inflammation
Lipotoxicity
Adipose Tissue-Derived Mediators
Gut-Derived Mediators
2.3.2. Role of Innate and Adaptive Immunity Components
Hepatocytes and Sinusoidal Endothelial Cells
Macrophages
Neutrophils
Natural Killer and Innate Lymphoid Cells
B Lymphocytes
Conventional T Lymphocytes
Innate-like T Cells
3. Therapeutic Strategies According to the Disturbed Underlying Mechanism
3.1. Modulators of Lipid and Carbohydrate Metabolism
3.1.1. Non-Pharmacological Treatments
3.1.2. Peroxisome Proliferator-Activated Receptor Agonists
3.1.3. Fibroblast Growth Factor Analogues
3.1.4. De Novo Lipogenesis Inhibition
3.1.5. Treatments Targeting Cholesterol Metabolism
3.2. Antihyperglycemic Drugs
3.2.1. Metformin and Dipeptidyl Peptidase 4 Inhibitors
3.2.2. Glucagon-like Peptide-1 Receptor Agonists
3.2.3. Sodium Glucose Co-Transporter 2 Inhibitors
3.3. Immune-Related Targets
3.3.1. TLR4 Inhibitors
3.3.2. CCR2/CCR5 Antagonists
3.4. Microbiome-Targeted Therapy
3.4.1. Prebiotics, Probiotics, and Synbiotics
3.4.2. Fecal Microbiota Transplantation
3.4.3. Potential Intervention with Bacteriophages

{kind=link}
| Identifier | Design | Drug (Mechanism) | Patients (n) | Patients’ Profile | Drug Administration | Primary Outcome | Main Outcomes | Refs | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lipid Profile | Glucidic Profile | Immunologic Profile | Microbiotic Profile | Dose | Period | |||||||
| Drugs targeting lipids and carbohydrates metabolism | ||||||||||||
| NCT00063622 (PIVENS) | Phase III | Pioglitazone (PPARγ agonist) | Non-diabetic patients with biopsy-proven NASH (247) | TGs: 165 ± 93 mg/dL TChol: 196 ± 39 mg/dL in all patients at baseline | Glucose: 94 ± 13 mg/dL HOMA-IR: 5.2 ± 4.3 in all patients at baseline | Hepatic inflammation | NA | Pioglitazone: 30 mg | Once daily | 1.Improvement in NAFLD activity defined by change in standardized scoring of liver biopsies at 96 weeks | Significant improvement of histological features, excluding fibrosis, in comparison to PBO group. 47% of the subjects reached NASH resolution | [196] |
| PBO | Once daily | |||||||||||
| NCT00994682 (UTHSCSA NASH Trial) | Phase IV | Pioglitazone (PPARγ agonist) | Biopsy-proven NASH and pre-diabetes or T2DM patients (101) | FFA: 0.49 ± 0.18 mmol/L TGs: 224 ± 171 mg/dL TChol: 187 ± 46 In pioglitazone group | T2DM: 48% Glucose: 124 ± 29 HbA1c: 5.7 ± 0.5 (in non-T2DM patients) and 7.1 ± 0.9 (in T2DM patients) Insulin: 15 ± 11 µU/mL in pioglitazone group | Hepatic inflammation Neutrophils count: ≥1500/mm3 Platelets: ≥ 100,000/mm3 in all patients | NA | Pioglitazone: 30 mg (if tolerated 45 mg) | Once daily, 8 weeks | 1. ≥2-point reduction in NAS (in at least two different histological categories) without worsening of fibrosis at 18 months | 58% of patients assigned to pioglitazone achieved the primary outcome, whereas 51% had NASH resolution. Pioglitazone improved individual histological scores, including the fibrosis score, and insulin sensitivity. | [197] |
| PBO | Once daily | |||||||||||
| Open label pioglitazone (all patients) | Once daily for an additional 18 months | |||||||||||
| NCT00492700 (FLIRT 2) | Phase II (Extension phase) | Rosiglitazone (PPARγ agonist) | Biopsy-proven NASH with increased transaminase values (53) | TGs: 1.5 ± 1.1 mmol/L HDL: 1.2 ± 0.5 in treated group | T2DM: 24% Glucose: 5.5 ± 2 mmol/L Insulin: 13.5 ± 8.5 µUI/L HOMA: 3.3 ± 3.4 in treated group | Hepatic inflammation | NA | Rosiglitazone: 8 mg (4 mg the 1st month) | Once daily | 1.Reduction in steatosis ≥30% | No improvement in the NAS score and histological features after 2 additional years of rosiglitazone administration | [195] |
| PBO | Once daily | |||||||||||
| NCT02704403 (RESOLVE-IT) | Phase III | Elafibranor (Dual PPARα/δ agonists) | Biopsy-proven NASH patients with BMI ≤ 45 kg/m2 (2157) | NA | 49.6% T2DM | Hepatic inflammation | NA | Elafibranor: 120 mg | Once daily | 1.Resolution of NASH without worsening of fibrosis. 2. Long-term outcome composed of all-cause mortality, cirrhosis, and liver-related clinical outcomes | Terminated, not accomplished | NA |
| PBO | Once daily | |||||||||||
| NCT03008070 (NATIVE) | Phase II | Lanifibranor (Pan-PPAR agonists) | Biopsy-proven NASH patients with BMI < 45 kg/m2 (247) | Tchol: 1.2 ± 0.3 mmol/L TGs: 2.0 ± 0.9 mmol/L | HbA1c: ≤8.5% Glucose: <10 mmol/L Insulin: 246.9 ± 260.7 pmol/L | Hepatic inflammation | NA | Lanifibranor: 800 mg | Once daily | 1. SAF-A decrease of at least 2 points with no worsening of the CRN-F | SAF-A decrease of at least 2 points with no worsening of CRN-F for 1200 mg dose RR = 1.82 (95% CI 1.24, 2.4) | [203] |
| Lanifibranor: 1200 mg | Once daily | |||||||||||
| PBO | Once daily | |||||||||||
| NCT02413372 | Phase II | Pegbelfermin (FGF21 analogue) | Biopsy-proven NASH with BMI ≥ 25 kg/m2 (184) | TGs: 187 ± 55 mg/dL HDL-C: 45 ± 12 mg/dL LDL-C: 120 ± 36 mg/dL | HbA1c: 6.2 ± 1.1% | Hepatic inflammation | NA | Pegbelfermin: 10 mg | Daily | 1. Mean change in percent hepatic fat fraction by MRI | Significant decrease in absolute hepatic fat fraction in 10 mg/day and 20 mg/week groups compared with PBO group | [206] |
| Pegbelfermin: 20 mg | Weekly | |||||||||||
| PBO | Daily | |||||||||||
| NCT03976401 | Phase II | Efruxifermin (Fc-FGF21 fusion protein) | Biopsy-proven NASH with BMI ≥ 25 kg/m2 and confirmation of ≥10% liver fat content on MRI-PDFF (80) | TGs: 180.0 ± 99.0 mg/dL TChol: 175.7 ± 45.1 mg/dL Apo B: 94.7 ± 27.8 mg/dL Apo C3: 9.4 ± 4.9 mg/dL | HbA1c: 6.23 ± 1.2% Glucose: 134.8 ± 66.2 mg/dL HOMA-IR: 14.1 ± 12.3 | Hepatic inflammation | NA | Efruxifermin: 28 mg | Once weekly | 1. Change from baseline in hepatic fat fraction assessed by MRI-PDFF | All efruxifermin-treated patients achieved ≥30%, and 88% achieved ≥50%, relative reduction in liver fat | [211] |
| Efruxifermin: 50 mg | Once weekly | |||||||||||
| Efruxifermin: 70 mg | Once weekly | |||||||||||
| PBO | Once weekly | |||||||||||
| NCT03298464 | Phase I | NGM313 (β-Klotho/FGFR1 agonist) | Insulin resistant and obese patients (i.e., BMI: 30–43 kg/m2) with both increased liver fat and normal ECG readings (25) | NA | NA | NA | NA | NGM313: 240 mg | Single dose | 1. Evaluation of whole body insulin sensitivity measured as insulin sensitivity index (M and Si) following intravenous insulin administration | Significant reductions in LFC (measured by MRI-PDFF), HbA1c, TGs, and LDL-C; and an increase in HDL-C, in NGM313-treated group | [212] |
| Pioglitazone: 45 mg | Daily | |||||||||||
| NCT02856555 | Phase II | Firsocostat (ACC inhibitor) | MRI-PDFF ≥8% Liver stiffness by MRE >2.5 kPa (126) | TGs: 160 (125, 201) mg/dL Tchol: 179 (152, 203) mg/dL | HbA1c: 6.5 (5.8, 7.8) % Glucose: 117 (97, 149) mg/dL HOMA-IR: 8.7 (5.3, 13.0) Insulin: 26.1 (17.0, 48.9) µIU/ml | NA | NA | Firsocostat: 5 mg | Once daily | 1.Percentage of participants experiencing treatment-emergent adverse events | AEs were experienced by 71% of patients receiving GS-0976, and by 62% with PBO | [213] |
| Firsocostat: 20 mg | Once daily | |||||||||||
| PBO | Once daily | |||||||||||
| NCT03776175 | Phase II | PF-05221304 (ACC inhibitor) PF-06865571 (DGAT2 inhibitor) | Metabolic syndrome (99) | TGs: 175.3 ± 66.8 mg/dL Tchol: 194.5 ± 36.6 mg/dL Apo C3: 13.9 ± 7.0 mg/dL | HbA1c: 5.8 ± 1% Glucose ≥100 mg/dL | NA | NA | PF-05221304: 15 mg | Twice daily | 1. Percent change from baseline in whole liver PDFF | PF-05221304 and PF-06865571 co-administration lowered steatosis by −44.6%, which was numerically greater than the reduction with PF-06865571 alone, but similar to that with PF-05221304 monotherapy | [214] |
| PF-06865571: 300 mg | Twice daily | |||||||||||
| PF-05221304: 15 mg + PF-06865571: 300 mg | Twice daily | |||||||||||
| PBO | Twice daily | |||||||||||
| NCT03938246 (FASCINATE-1) | Phase II | TVB-2640 (FASN Inhibitor) | Biopsy-proven NASH/overweight/obese/diabetic/ALT ≥ 30 U/L in patients with BMI ≤ 40 kg/m2 and ≥8% liver fat content on MRI-PDFF. (142) | TGs: 163 (124, 262) mg/dL Tchol: 189 (167, 225) mg/dL Apo B: 104 (89, 124) mg/dL | HbA1c: 5.8 (5.5, 6.4) % Insulin: 22 (14, 32) µU/mL HOMA-IR: 5.0 (3.7, 7.8) Glucose: 98 (80, 124) mg/dL | Hepatic inflammation | NA | TVB-2640: 25 mg | Once daily | 1. Change in hepatic fat fraction from baseline in subjects with NASH by MRI PDFF 2. Safety of TVB-2640, including changes in liver enzymes by monitoring AEs | TVB-2640 treatment resulted in significant relative and absolute reductions of liver fat compared to PBO in a dose-dependent manner | [215] |
| TVB-2640: 50 mg | Once daily | |||||||||||
| PBO | Once daily | |||||||||||
| NCT02279524 (ARREST) | Phase II | Aramchol (SCD1 inhibitor) | Biopsy-proven NASH and BMI between 25 kg/m2 to 40 kg/m2 (247) | TGs: 1.92 ± 1.6 mmol/L Tchol: 4.88 ± 1.1 mmol/L | Glucose: 6.94 ± 2.4 mmol/L HbA1c: 6.65 ± 1.0% HOMA-IR: 9.6 ± 6.5 U | Hepatic inflammation | NA | Aramchol: 400 mg | Once daily | 1. Change from baseline in mean liver fat | NASH resolution without worsening fibrosis was achieved in Aramchol 600 mg: OR = 4.74 (95% CI 0.99, 22.7) | [216] |
| Aramchol: 600 mg | Once daily | |||||||||||
| PBO | Once daily | |||||||||||
| NCT01265498 (FLINT) | Phase II | Obeticholic acid (FXR agonist) | Biopsy-proven NASH | TGs: 2.2 ± 1.5 mmol/l TChol 4.9 ± 1.2 mmol/l | Glucose: 6.5 ± 1.8 mmol/L Insulin: 201 ± 226 pmol/L HbA1c: 48 ± 12 mmol/mol HOMA-IR: 61 ± 74 | Hepatic inflammation | NA | OCA: 25 mg | Once daily | 1. Improvement in liver histology, defined as a decrease in NAFLD activity score by at least 2 points without worsening of fibrosis | OCA met primary outcome in 46% of patients compared with 21% of patients in the PBO group | [223] |
| PBO | Once daily | |||||||||||
| NCT03449446 (ATLAS) | Phase II | Cilofexor (FXR agonist) Firsocostat (ACC1/ACC2 inhibitor) Selonsertib (Authopagy inhibitor) | Advanced fibrosis (395) | TGs: 123 (101, 191) mg/dL Tchol: 177 (150, 208) mg/dL Total BAs: 7.1 (4.9, 11.6) µmol/L | Glucose: 111 (97, 138) mg/dL HbA1c: 6.0 (5.6, 6.6) % HOMA-IR: 6.2 (3.8, 8.3) | Hepatic inflammation | NA | Cilofexor:30 mg | Once daily | 1. Percentage of participants experiencing treatment-emergent adverse events 2. Percentage of participants experiencing treatment-emergent laboratory abnormalities 3. Percentage of participants who achieved a ≥1-Stage improvement in fibrosis without worsening of NASH | 1. Thirteen patients (3%) discontinued treatment due to an AE, with similar rates between treatment groups 2. Grade 3 and 4 laboratory abnormalities were observed in 0–13% of patients across groups 3. Differences between the treatment arms and PBO (11%) did not reach statistical significance | [225] |
| Firsocostat: 20 mg | Once daily | |||||||||||
| Selonsertib: 18 mg | Once daily | |||||||||||
| Selonsertib: 18 mg + Firsocostat: 20 mg | Once daily | |||||||||||
| Selonsertib: 18 mg + Cilofexor:30 mg | Once daily | |||||||||||
| Firsocostat: 20 mg + Cilofexor:30 mg | Once daily | |||||||||||
| PBO | Once daily | |||||||||||
| Antihyperglycemic drugs | ||||||||||||
| NCT01237119 (LEAN) | Phase II | Liraglutide (GLP1-RA) | Biopsy proven NASH patients with BMI ≥ 25 kg/m² (52) | Hyperlipidaemia: 9 (35%) in liraglutide group. Other parameters in liraglutide group: Esterifi ed fatty acids: 967 ± 535 μmol/L; ADIPO-IR: 22.2 ± 12.7 U; TChol: 4.5 ± 1.1 mmol/L; TGs: 1.9 ± 1.1 mmol/L | HbA1c ≤ 9% T2DM: 9 (35%) in liraglutide group; Other parameters in liraglutide group: Glucose: 6.0 ± 1.7 mmol/L; Insulin: 166 ± 80 pmol/L HOMA-IR: 6.7 ± 4.7 U | Hepatic inflammation | NA | Liraglutide: 1.8 mg | Once daily | 1. Liver histological improvement | Resolution of NASH with no worsening fibrosis in 39% in liraglutide group vs. 9% in the PBO group. Progression of fibrosis 9% vs. 36%. Reduction of body weight, and BMI improvement of physical activity, and no severe AEs. | [232] |
| PBO | Once daily | |||||||||||
| NCT02970942 | Phase II | Semaglutide (GLP1-RA) | Biopsy proven NASH with fibrosis stage 1, 2 or 3 patients with BMI > 25 kg/m² (320). | NA | HbA1c ≤ 10% Parameters in highest dose semaglutide group: T2DM: 49 ± 60% | Hepatic inflammation | NA | Semaglutide: 0.1 mg | Once daily | 1. Percentage of participants with NASH resolution without worsening of fibrosis after 72 weeks | Percentage of patients in whom NASH resolution was achieved with no worsening of fibrosis: 40% in the 0.1-mg group, 36% in the 0.2-mg group, 59% in the 0.4-mg group (p < 0.001), and 17% in the PBO group. An improvement in fibrosis stage occurred in 43% of the patients in the 0.4-mg group, and in 33% of the patients in the PBO group. Mean percent weight loss was 13% in the 0.4-mg group, and 1% in the PBO group. | [233] |
| Semaglutide: 0.2 mg | Once daily | |||||||||||
| Semaglutide: 0.4 mg | Once daily | |||||||||||
| PBO | Once daily | |||||||||||
| NCT03131687 | Phase II | Tirzepatide (GLP1-RA with gastric inhibitory polypeptide receptor activity) | Patients with T2DM with BMI ≥ 23 and <50 kg/m² (316) | Adiponectine: 5.1 ± 0.5 mg/L in highest dose of tirzepatide group | HbA1c: ≥7% and ≤10.5% Glucose: 164.8 ± 48.6 mg/dL in highest tirzepatide group | NA | NA | Tirzepatide: 1 mg | Once weekly | 1. Change from baseline to week 26 in HbA1c Bayesian dose response. | Decreases with tizepartide were significant compared to PBO for K-18 (10 mg) and pro-C3 (15 mg) and with dulaglutide for ALT (10 and 15 mg). Adiponectine significantly increased from baseline with tizepartide compared to PBO (10 and 15 mg). | [244] |
| Tirzepatide: 5 mg | Once weekly | |||||||||||
| Tirzepatide: 10 mg | Once weekly | |||||||||||
| Tirzepatide: 15 mg | Once weekly | |||||||||||
| Dulaglutide: 1.5 mg | Once weekly | |||||||||||
| PBO | Once weekly | |||||||||||
| NCT03235050 | Phase II | Cotadutide (GLP1-RA with glucagon activity) and liraglutide (GLP1-RA) | Patients with treated T2DM BMI ≥ 25 kg/m² (834) | TGs: <1000 mg/dL | HbA1c ≥ 7% and ≤10.5% | NA | NA | Cotadutide: 100 μg | Once daily | 1. Change in HbA1c 2. Percent change in body weight | Cotagutide 300 μg yielded greater reduction in body weight and ALT levels vs. liraglutide. The improvement in FLI and NFS with cotadutide may indicate reduced liver fat and fibrosis, respectively. | [245] |
| Cotadutide: 200 μg | Once daily | |||||||||||
| Cotadutide: 300 μg | Once daily | |||||||||||
| Liraglutide_ 1.8 mg | Once daily | |||||||||||
| PBO | Once daily | |||||||||||
| NCT02686476 (E-LIFT) | NA | Empagliflozin (SGLT2i) | Patients with T2DM of age ≥ 20 years and NAFLD (100) | Parameters in the Empaglifozin group at baseline: TGs: 201 ± 124) mg/dL; HDL: 42 ± 12 mg/dL; LDL: 112 ± 35 mg/dL | HbA1c > 7.0% and <10.0% Glucose in the Emplaglifozin group at baseline: 173 ± 44 mg/dL | NA | NA | Standard care of T2DM * + Empagliflozin: 10 mg | Once daily | 1. To evaluate the change in liver fat content at baseline and 3 months. | Mean MRI-PDFF difference between the empagliflozin and control groups −4.0% (p < 0.001). | [253] |
| NCT02964715 | Phase IV | Empagliflozin (SGLT2i) | Obese patients with T2DM, biopsy proven NASH and BMI < 45 kg/m² (25) | Dyslipidemia 8 (88.9%); TGs: 1.6 (1.3–2.4) mmol/L; TChol: 4.4 (3.5–4.7) mmol/L | HbA1c: >6.5% | Hepatic inflammation | NA | Any anti-diabetic agent except SGLT2 inhibitors, TZDs, DPP4 inhibitors and GLP1RAs + Empagliflozin: 25 mg | Daily for 6 months | 1. Change in histological grade as evaluated with non-alcoholic Steatohepatitis Clinical Research Network scoring system 2. Change in serum FGF21 | Empaglifozin resulted in signifcantly greater improvements in steatosis (67% vs. 26%, p = 0.025), ballooning (78% vs. 34%, p = 0.024), and fbrosis (44% vs. 6%, p = 0.008) compared with PBO. | [261] |
| Drugs targeting immunologic system | ||||||||||||
| NCT02442687 | Phase II | JKB-121 (TLR-4 antagonist) | Biopsy-proven NASH patients with BMI > 25 kg/m2 (65) | NA | HbA1c: ≤9% | Hepatic inflammation | NA | JKB-121: 5 mg | Twice daily | 1. Reduction in liver fat content by MRI-PDFF change from baseline to week 24. 2. Reduction in liver fat content by MRI-PDFF change from baseline to week 12. | Not accomplished | [263] |
| JKB-121: 10 mg | Twice daily | |||||||||||
| PBO | Twice daily | |||||||||||
| NCT02316717 | Phase ll | IMM-124E (anti-LPS polyclonal antibodies) | Biopsy proven NASH patients with BMI > 25 kg/m2 (133) | NA | HbA1c: <9% | Hepatic inflammation | NA | IMM-124E:600 mg | Three times daily | 1. Incidence of AEs per arm/group. 2. Mean change from baseline in percentage fat content of the liver. measured by MRI at week 24. 3. Number of patients with treatment-related AEs. 4. Number of grade 3–5 AEs. | NA | NA |
| IMM-124E:1200 mg | Three times daily | |||||||||||
| PBO | Three times daily | |||||||||||
| NCT02217475 (CENTAUR study) | Phase ll | CVC (dual CCR2/CCR5 antagonist) | Biopsy-proven NASH and liver fibrosis patients with mean BMI 33.9 ± 6.5 kg/m2 (289) | Biomarkers in all patients at baseline TGs: 177.4 ± 130.8 mg/dLTChol: 190.2 ± 48.1 mg/dL | HbA1c: 6.54 ± 1.27% | Hepatic inflammation biomarkers at baseline in CVC group (median (min, max)): hs-CRP: 2.35 (0.2, 24.0) mg/L; IL-1β: 0.090 (0.00, 2.69) pg/mL; IL-6: 4.30 (1.4, 475.6) pg/mL; sCD14: 1731.0 (138, 3601) μg/L; sCD163: 615.0 (263, 1486) μg/L; CCL2: 499.00 (166.1, 1497.4) pg/mL; CCL4: 90.80 (2.6, 2432.9) pg/mL | NA | CVC: 150 mg | Once daily | 1. Number of participant with hepatic histological improvement in NAS by ≥2 points with at least 1-point reduction in either lobular inflammation or hepatocellular ballooning, and no concurrent worsening of fibrosis at year | Improvement in fibrosis by ≥1 stage (NASH CRN system) and no worsening of steatohepatitis (no worsening of lobular inflammation or hepatocellular ballooning grade): OR = 2.201 (95% CI 1.113, 4.352) | [265] |
| PBO | Once daily | |||||||||||
| NCT03028740 (AURORA Study) | Phase III | CVC (dual CCR2/CCR5 antagonist) | Biopsy- proven NASH and stage 2 or 3 liver fibrosis patients (1779) | NA | HbA1c: ≤10% | Hepatic inflammation | NA | CVC: 150 mg | Once daily For year 1 or years 1&2 | 1. Superiority of CVC compared to PBO on liver histology at month 12 relative to the screening biopsy. 2. Superiority of CVC compared to PBO on the composite endpoint of histopathologic progression to cirrhosis, liver-related clinical outcomes, and all-cause mortality. | Not accomplished (study early terminated) | NA |
| PBO | Once daily for year 2 | |||||||||||
| Drugs targeting microbiota | ||||||||||||
| NCT03434860 | NA | Symbiter (14 strains from Bifidobacterium, Lactobacillus, Lactococus, Propionibacterium and Acetobacter) | T2DM patients of age 18–65 with NAFLD and BMI ≥ 25 kg/m2 (58) | TGs: 2.57 ± 1.03 mmol/L TChol: 6.28 ± 0.89 mmol/L | NA | Hepatic inflammation | NA | Symbiter: 10 g | Once daily | 1. Changes in LFC 2. Changes in liver stiffness | Decrease of LFC, serum insulin and leptin, and IR compared with PBO | [267] |
| IRCT201410052394N13 | Phase II | Orafti (L. acidophilus, L. casei, L. rhamnosus, B. breve and B. longum) | Patients of age 20–60 with NAFLD and BMI ≥ 25 kg/m2 (89) | NA | Glucose: 89 ± 17 mg/dL Insulin: 11.42 ± 4.5 µU/mL HOMA-IR: 2.58 ± 1.35 | Hepatic inflammation | NA | Orafti: 8 g | Twice daily | 1. Modulation of glycemic parameters | Decrease of LFC, serum insulin and leptin, and IR compared with PBO | [268] |
| NCT01791959 | Phase II | Protexin (L. acidophilus, L. bulgaricus, L. casei, L. rhamnosus, B. breve, B. longum and S. thermophilus) | Patients of age > 18 with NAFLD determined by steatosis and ALT > 60 U/L (52) | NA | Glucose: 99.6 ± 24.2 mg/dL Insulin: 11.2 ± 3.4 µU/mL HOMA-IR: 2.8 ± 1 | Hepatic inflammation | NA | Protexin: NA | Twice daily | 1. Modulation of hepatic fibrosis, liver enzymes, and inflammatory markers | Significant decrease of serum ALT compared with PBO | [269] |
| IRCT201111082709N22 | Phase II | Protexin (L. acidophilus, L. bulgaricus, L. casei, L. rhamnosus, B. breve, B. longum and S. thermophilus) + vitamin E | Patients of age 25–64 with NAFLD determined by steatosis and persistently elevated ALT > 30 mg/dL (60) | TGs: 162.56 ± 18.83 mg/dL TChol: 167.3 ± 18.79 mg/dL Apo B: 85.7 ± 18.2 mg/dL | Glucose: 98.63 ± 7.14 mg/dL Insulin: 1.77 ± 0.53 µU/mL HOMA-IR: 0.63 ± 0.2 µU/mL | Hepatic inflammation | NA | Protexin + vitamin E: 400 IU | Twice daily | 1. Modulation of liver enzymes, leptin, lipid profile, and IR | Significantive decrease of serum ALT, leptin, plasma glucose, IR, TG, cholesterol, and LDL compared with PBO | [270] |
| IRCT2017020932417N2 | NA | Synbiotic yogurt with B. animalis | Patients of age > 18 with NAFLD determined by ultrasound and grade 1–3 fatty liver (102) | TGs: 165.7 ± 60.9 mg/dL TChol: 195.3 ± 34.7 mg/dL | Glucose: 89 ± 17 mg/dL Insulin: 11.42 ± 4.5 µU/mL HOMA-IR: 2.58 ± 1.35 | Hepatic inflammation | NA | NA | Three times a day | 1. Hepatic steatosis and modulation of liver enzymes | Reduction of steatosis and decrease of serum ALT, AST, and GGT compared with PBO | [271] |
| NCT01680640 (INSYTE) | NA | Actilight with B. animalis | Patients with NAFLD (104) | TGs: 1.8 (1.1) mmol/L TChol: 4.9 (1.2) mmol/L | Glucose: 6.2 (2.5) mmol/L Insulin: 13.5 (7.9) µU/mL | Hepatic inflammation | Higher proportions of Bifidobacterium and Faecalibacterium, and lower proportions of Oscillibacter and Allistipes in fecal samples | NA | Twice daily | 1. Modulation of liver fat content, biomarkers of liver fibrosis 2. Composition of fecal microbiome | The administration of a synbiotic altered fecal microbiome, but did not reduce liver fat content or markers of liver fibrosis compared with PBO | [272] |
| NA | NA | Familact (L. acidophilus, L. bulgaricus, L. casei, L. rhamnosus, B. breve, B. longum and S. thermophilus) + sitagliptin | Patients of age 18–60 with NAFLD determined by ultrasound and BMI 25–30 kg/m2 (138) | TGs: 203.84 ± 47.40 mg/dL TChol: 205.84 ± 29.29 mg/dL | Glucose: 103.25 ± 3.63 mg/dL | Hepatic inflammation | NA | Familact: 500 mg | Once daily | 1. Effect of sitagliptin | Significant reduction of BMI, plasma glucose, ALT, AST, cholesterol, and TGs compared with PBO | [273] |
| IRCT2013122811763N15 | NA | Familact (L. acidophilus, L. bulgaricus, L. casei, L. rhamnosus, B. breve, B. longum and S. thermophilus) | Patients of age 18–60 with NAFLD determined by ultrasound (80) | NA | NA | Hepatic inflammation | NA | Familact: 500 mg | Once daily | 1. Effects of symbiotic on C-reactive protein and liver enzymes | Significantly reduced steatosis on abdominal ultrasound compared with PBO | [274] |
| IRCT201301223140N6 | Phase II | Synbiotic (L. acidophilus and B. longum) | Patients of age 20–60 with high levels of AST and ALT and NAFLD determined by ultrasound (75) | NA | NA | NA | NA | Familact: 250 mg | Twice daily | 1. Supplementation with probiotics and/or prebiotics on liver function | The treatment lowered ALT and BMI compared with PBO | [275] |
| NCT02496390 | Phase II | FMT | Patients of age > 18 with NAFLD determined by AASLD criteria (21) | TGs: 2.30 (1.43) mmol/L TChol: 4.68 (1.15) mmol/L Apo B: 1.13 (0.35) g/L | Glucose: 7.3 (1.8) mmol/L Insulin: 196 (177) pmol/L HOMA-IR: 3.5 (1.3) | NA | No changes | FMT: 2 g | Once | 1. Improvement of IR, hepatic proton density fat fraction, and intestinal permeability | Half of the patients with elevated small intestinal permeability at baseline had a signifi-cant reduction 6 weeks after allogenic transplant, coupled with an increase in GM diversity | [277] |
| NTR4339 | NA | FMT | Patients of age 21–69 with BMI > 25 kg/m2, suspicion of NAFLD (elevated liver enzymes, impaired glucose tolerance and severity of steatosis on ultrasound) (21) | TGs: 1.4 ± 0.5 mmol/L TChol: 6 ± 0.8 mmol/L | Glucose: 5.8 ± 0.7 mmol/L | Hepatic inflammation | Increase in Ruminococcus, Eubacterium hallii, Faecalibacterium, and Prevotella copri in allogenic FMT. Increase in Lachnospiraceae in autologous FMT. | NA | NA | 1. Modulation of GM composition through FMT | Allogenic FMT altered GM composition, and led to beneficial changes in plasma metabolites and genic expression involved in hepatic inflammation or lipid metabolism | [279] |
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ogawa, Y.; Imajo, K.; Honda, Y.; Kessoku, T.; Tomeno, W.; Kato, S.; Fujita, K.; Yoneda, M.; Saito, S.; Saigusa, Y.; et al. Palmitate-induced lipotoxicity is crucial for the pathogenesis of nonalcoholic fatty liver disease in cooperation with gut-derived endotoxin. Sci. Rep. 2018, 8, 11365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Mota, M.; Banini, B.A.; Cazanave, S.C.; Sanyal, A.J. Molecular mechanisms of lipotoxicity and glucotoxicity in nonalcoholic fatty liver disease. Metabolism 2016, 65, 1049–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasr, P.; Ignatova, S.; Kechagias, S.; Ekstedt, M. Natural history of nonalcoholic fatty liver disease: A prospective follow-up study with serial biopsies. Hepatol. Commun. 2018, 2, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paik, J.M.; Henry, L.; De Avila, L.; Younossi, E.; Racila, A.; Younossi, Z.M. Mortality related to nonalcoholic fatty liver disease is increasing in the United States. Hepatol. Commun. 2019, 3, 1459–1471. [Google Scholar] [CrossRef] [Green Version]
- Rosato, V.; Masarone, M.; Dallio, M.; Federico, A.; Aglitti, A.; Persico, M. NAFLD and extra-hepatic comorbidities: Current evidence on a multi-organ metabolic syndrome. Int. J. Environ. Res. Public Health 2019, 16, 3415. [Google Scholar] [CrossRef] [Green Version]
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wong, V.W.-S.; Dufour, J.-F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef]
- Lin, S.; Huang, J.; Wang, M.; Kumar, R.; Liu, Y.; Liu, S.; Wu, Y.; Wang, X.; Zhu, Y. Comparison of MAFLD and NAFLD diagnostic criteria in real world. Liver Int. 2020, 40, 2082–2089. [Google Scholar] [CrossRef]
- Yamamura, S.; Eslam, M.; Kawaguchi, T.; Tsutsumi, T.; Nakano, D.; Yoshinaga, S.; Takahashi, H.; Anzai, K.; George, J.; Torimura, T. MAFLD identifies patients with significant hepatic fibrosis better than NAFLD. Liver Int. 2020, 40, 3018–3030. [Google Scholar] [CrossRef]
- Juanola, O.; Martínez-López, S.; Francés, R.; Gómez-Hurtado, I. Non-alcoholic fatty liver disease: Metabolic, genetic, epigenetic and environmental risk factors. Int. J. Environ. Res. Public Health 2021, 18, 5227. [Google Scholar] [CrossRef]
- Dallio, M.; Sangineto, M.; Romeo, M.; Villani, R.; Romano, A.D.; Loguercio, C.; Serviddio, G.; Federico, A. Immunity as cornerstone of non-alcoholic fatty liver disease: The contribution of oxidative stress in the disease progression. Int. J. Mol. Sci. 2021, 22, 436. [Google Scholar] [CrossRef]
- Ashraf, N.U.; Sheikh, T.A. Endoplasmic reticulum stress and Oxidative stress in the pathogenesis of Non-alcoholic fatty liver disease. Free Radic. Res. 2015, 49, 1405–1418. [Google Scholar] [CrossRef]
- Fryk, E.; Olausson, J.; Mossberg, K.; Strindberg, L.; Schmelz, M.; Brogren, H.; Gan, L.-M.; Piazza, S.; Provenzani, A.; Becattini, B.; et al. Hyperinsulinemia and insulin resistance in the obese may develop as part of a homeostatic response to elevated free fatty acids: A mechanistic case-control and a population-based cohort study. EBioMedicine 2021, 65, 103264. [Google Scholar] [CrossRef]
- Kucuk, S.; Niven, J.; Caamano, J.; Jones, S.W.; Camacho-Muñoz, D.; Nicolaou, A.; Mauro, C. Unwrapping the mechanisms of ceramide and fatty acid-initiated signals leading to immune-inflammatory responses in obesity. Int. J. Biochem. Cell Biol. 2021, 135, 105972. [Google Scholar] [CrossRef]
- Zhan, Y.-T. Roles of liver innate immune cells in nonalcoholic fatty liver disease. World J. Gastroenterol. 2010, 16, 4652–4660. [Google Scholar] [CrossRef] [PubMed]
- Nunes, V.S.; Andrade, A.R.; Guedes, A.L.V.; Diniz, M.A.; Oliveira, C.P.; Cançado, E.L.R. Distinct phenotype of non-alcoholic fatty liver disease in patients with low levels of free copper and of ceruloplasmin. Arq. Gastroenterol. 2020, 57, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Olefsky, J.M. Inflammation and lipid signaling in the etiology of insulin resistance. Cell Metab. 2012, 15, 635–645. [Google Scholar] [CrossRef] [Green Version]
- European Association for the Study of the Liver; European Association for the Study of Diabetes; European Association for the Study of Obesity. EASL–EASD–EASO clinical practice guidelines for the management of non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1388–1402. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.D.; Targher, G. NAFLD: A multisystem disease. J. Hepatol. 2015, 62, S47–S64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, W.; Cheng, X.-F.; Liu, Y.; Lv, Q.-Z.; Liu, G.-L.; Zhang, J.-G.; Li, X.-Y. Potential nexus of non-alcoholic fatty liver disease and type 2 diabetes Mellitus: Insulin resistance between hepatic and peripheral tissues. Front. Pharmacol. 2019, 9, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannou, S.; Haslam, D.; McKeown, N.M.; Herman, M.A. Fructose metabolism and metabolic disease. J. Clin. Investig. 2018, 128, 545–555. [Google Scholar] [CrossRef]
- Bubici, C.; Papa, S.; Pham, C.G.; Zazzeroni, F.; Franzoso, G. The NF-kappaB-mediated control of ROS and JNK signaling. Histol. Histopathol. 2005, 21, 69–80. [Google Scholar]
- Kubes, P.; Jenne, C. Immune responses in the liver. Ann. Rev. Immunol. 2018, 36, 247–277. [Google Scholar] [CrossRef] [PubMed]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Genet. 2021, 19, 77–94. [Google Scholar] [CrossRef]
- Perakakis, N.; Stefanakis, K.; Mantzoros, C.S. The role of omics in the pathophysiology, diagnosis and treatment of non-alcoholic fatty liver disease. Metabolism 2020, 111, 154320. [Google Scholar] [CrossRef]
- Rada, P.; González-Rodríguez, Á.; García-Monzón, C.; Valverde, Á.M. Understanding lipotoxicity in NAFLD pathogenesis: Is CD36 a key driver? Cell Death Dis. 2020, 11, 802. [Google Scholar] [CrossRef]
- Jahn, D.; Kircher, S.; Hermanns, H.M.; Geier, A. Animal models of NAFLD from a hepatologist’s point of view. Biochim. Biophys. Acta—Mol. Basis Dis. 2019, 1865, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Esmaili, S.; Rogers, G.B.; Bugianesi, E.; Petta, S.; Marchesini, G.; Bayoumi, A.; Metwally, M.; Azardaryany, M.K.; Coulter, S.; et al. Lean NAFLD: A distinct entity shaped by differential metabolic adaptation. Hepatology 2020, 71, 1213–1227. [Google Scholar] [CrossRef]
- Iacobini, C.; Pugliese, G.; Fantauzzi, C.B.; Federici, M.; Menini, S. Metabolically healthy versus metabolically unhealthy obesity. Metabolism 2019, 92, 51–60. [Google Scholar] [CrossRef]
- Yu, S.J.; Kim, W.; Kim, D.; Yoon, J.-H.; Lee, K.; Kim, J.H.; Cho, E.J.; Lee, J.-H.; Kim, H.Y.; Kim, Y.J.; et al. Visceral obesity predicts significant fibrosis in patients with non-alcoholic fatty liver disease. Medicine 2015, 94, 2159. [Google Scholar] [CrossRef]
- Di Renzo, L.; Tyndall, E.; Gualtieri, P.; Carboni, C.; Valente, R.; Ciani, A.S.; Tonini, M.G.; De Lorenzo, A. Association of body composition and eating behavior in the normal weight obese syndrome. Eat. Weight Disord. 2016, 21, 99–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimaudo, S.; Pipitone, R.M.; Pennisi, G.; Celsa, C.; Cammà, C.; Di Marco, V.; Barcellona, M.R.; Boemi, R.; Enea, M.; Giannetti, A.; et al. Association between PNPLA3 rs738409 C>G variant and liver-related outcomes in patients with nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 2020, 18, 935–944.e3. [Google Scholar] [CrossRef]
- Alonso, C.; Fernández-Ramos, D.; Varela-Rey, M.; Martínez-Arranz, I.; Navasa, N.; Van Liempd, S.M.; Trueba, J.L.L.; Mayo, R.; Ilisso, C.P.; de Juan, V.G.; et al. Metabolomic identification of subtypes of nonalcoholic steatohepatitis. Gastroenterology 2017, 152, 1449–1461.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad Guerra, J.; Arias Loste, M.T.; Burgos Santamaria, D.; Ampuero, J.; Martinez Porras, J.L.; Iruzubieta, P.; Calleja Panero, J.L. Interim analysis from a 72 week, double-blind, placebo-controlled, multicenter, paired liver biopsy study of endoscopic sleeve gastroplasty in patients with non-alcoholic steatohepatitis (NASH). J. Hepatol. 2021, 75, 269. [Google Scholar]
- Ludwig, D.S.; Hu, F.B.; Tappy, L.; Brand-Miller, J. Dietary carbohydrates: Role of quality and quantity in chronic disease. BMJ 2018, 361, k2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GBD Diet Collaborators. Health effects of dietary risks in 195 countries, 1990–2017: A systematic analysis for the global burden of disease study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef] [Green Version]
- Mei, S.; Yang, X.; Guo, H.; Gu, H.; Zha, L.; Cai, J.; Li, X.; Liu, Z.; Bennett, B.J.; He, L.; et al. A small amount of dietary carbohydrate can promote the HFD-induced insulin resistance to a maximal level. PLoS ONE 2014, 9, e100875. [Google Scholar] [CrossRef] [Green Version]
- Glass, M.; Hunt, C.; Fuchs, M.; Su, G.L. Comorbidities and non-alcoholic fatty liver disease: The chicken, the egg, or both? Fed. Pract. 2019, 36, 64–71. [Google Scholar] [PubMed]
- Mantovani, A.; Byrne, C.D.; Bonora, E.; Targher, G. Nonalcoholic fatty liver disease and risk of incident type 2 diabetes: A meta-analysis. Diabetes Care 2018, 41, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Kwok, R.; Choi, K.C.; Wong, G.L.-H.; Zhang, Y.; Chan, H.L.-Y.; Luk, A.O.-Y.; Shu, S.S.-T.; Chan, A.W.-H.; Yeung, M.-W.; Chan, J.C.-N.; et al. Screening diabetic patients for non-alcoholic fatty liver disease with controlled attenuation parameter and liver stiffness measurements: A prospective cohort study. Gut 2016, 65, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, R.A.; Cuadrado, N.M.; Tafur, C.; Gómez, J.J.L.; De La Cuesta, S.G.; Sánchez, M.C.G.; Melero, B.A.; Román, D.A.D.L. Histopathological differences in patients with biopsy-proven non-alcoholic fatty liver disease with and without type 2 diabetes. Endocrinol. Diabetes Nutr. 2018, 65, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, P. Serum metabolomics-based steatohepatitis score for the noninvasive identification of patients with non-alcoholic steatohepatitis (NASH) in multiethnic, including type 2 diabetes mellitus population. J. Hepatol. 2021, 75. Available online: https://www.postersessiononline.eu/173580348_eu/congresos/ILC2021/aula/-PO_1518_ILC2021.pdf (accessed on 28 September 2021).
- Li, Y.; Wang, J.; Tang, Y.; Han, X.; Liu, B.; Hu, H.; Li, X.; Yang, K.; Yuan, J.; Miao, X.; et al. Bidirectional association between nonalcoholic fatty liver disease and type 2 diabetes in Chinese population: Evidence from the Dongfeng-Tongji cohort study. PLoS ONE 2017, 12, e0174291. [Google Scholar] [CrossRef]
- Forlani, G.; Giorda, C.; Manti, R.; Mazzella, N.; De Cosmo, S.; Rossi, M.C.; Nicolucci, A.; Di Bartolo, P.; Ceriello, A.; Guida, P.; et al. The burden of NAFLD and its characteristics in a nationwide population with type 2 diabetes. J. Diabetes Res. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lyu, K.; Zhang, D.; Song, J.D.; Li, X.; Perry, R.J.; Samuel, V.T.; Shulman, G.I. Short-term overnutrition induces white adipose tissue insulin resistance through sn-1,2-diacylglycerol—PKCε—insulin receptorT1160 phosphorylation. JCI Insight 2021, 6, e139946. [Google Scholar] [CrossRef]
- Lyu, K.; Zhang, Y.; Zhang, D.; Kahn, M.; Horst, K.W.; Rodrigues, M.R.; Gaspar, R.C.; Hirabara, S.M.; Luukkonen, P.K.; Lee, S.; et al. A membrane-bound diacylglycerol species induces PKCϵ-mediated hepatic insulin resistance. Cell Metab. 2020, 32, 654–664.e5. [Google Scholar] [CrossRef] [PubMed]
- Peck, B.; Huot, J.; Renzi, T.; Arthur, S.; Turner, M.J.; Marino, J.S. Mice lacking PKC-θ in skeletal muscle have reduced intramyocellular lipid accumulation and increased insulin responsiveness in skeletal muscle. Am. J. Physiol. Integr. Comp. Physiol. 2018, 314, R468–R477. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.; Samuel, V.T. The sweet path to metabolic demise: Fructose and lipid synthesis. Trends Endocrinol. Metab. 2016, 27, 719–730. [Google Scholar] [CrossRef] [Green Version]
- Kolczynska, K.; Loza-Valdes, A.; Hawro, I.; Sumara, G. Diacylglycerol-evoked activation of PKC and PKD isoforms in regulation of glucose and lipid metabolism: A review. Lipids Health Dis. 2020, 19, 1–15. [Google Scholar] [CrossRef]
- Santoleri, D.; Titchenell, P.M. Resolving the paradox of hepatic insulin resistance. CMGH 2019, 7, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Dentin, R.; Tomas-Cobos, L.; Foufelle, F.; Leopold, J.; Girard, J.; Postic, C.; Ferré, P. Glucose 6-phosphate, rather than xylulose 5-phosphate, is required for the activation of ChREBP in response to glucose in the liver. J. Hepatol. 2012, 56, 199–209. [Google Scholar] [CrossRef]
- Song, Z.; Yang, H.; Zhou, L.; Yang, F. Glucose-sensing transcription factor MondoA/ChREBP as targets for type 2 diabetes: Opportunities and challenges. Int. J. Mol. Sci. 2019, 20, 5132. [Google Scholar] [CrossRef] [Green Version]
- Kubota, N.; Kubota, T.; Kajiwara, E.; Iwamura, T.; Kumagai, H.; Watanabe, T.; Inoue, M.; Takamoto, I.; Sasako, T.; Kumagai, K.; et al. Differential hepatic distribution of insulin receptor substrates causes selective insulin resistance in diabetes and obesity. Nat. Commun. 2016, 7, 12977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, Y.; Kubota, N.; Yamauchi, T.; Kadowaki, T. Role of insulin resistance in MAFLD. Int. J. Mol. Sci. 2021, 22, 4156. [Google Scholar] [CrossRef]
- Gastaldelli, A.; Gaggini, M.; DeFronzo, R.A. Role of adipose tissue insulin resistance in the natural history of type 2 diabetes: Results from the san antonio metabolism study. Diabetes 2017, 66, 815–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeBosch, B.J.; Chen, Z.; Saben, J.L.; Finck, B.N.; Moley, K.H. Glucose transporter 8 (GLUT8) mediates fructose-induced de novo lipogenesis and macrosteatosis. J. Biol. Chem. 2014, 289, 10989–10998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.-S.; Krawczyk, S.A.; Doridot, L.; Fowler, A.J.; Wang, J.X.; Trauger, S.A.; Noh, H.-L.; Kang, H.J.; Meissen, J.K.; Blatnik, M.; et al. ChREBP regulates fructose-induced glucose production independently of insulin signaling. J. Clin. Investig. 2016, 126, 4372–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, A.; Rosato, V.; Masarone, M.; Torre, P.; Dallio, M.; Romeo, M.; Persico, M. The role of fructose in non-alcoholic steatohepatitis: Old relationship and new insights. Nutrients 2021, 13, 1314. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Torres, A.; Henry, R.A.; Trefely, S.; Wallace, M.; Lee, J.V.; Carrer, A.; Sengupta, A.; Campbell, S.L.; Kuo, Y.-M.; et al. ATP-citrate Lyase controls a glucose-to-acetate metabolic switch. Cell Rep. 2016, 17, 1037–1052. [Google Scholar] [CrossRef] [Green Version]
- Becares, N.; Gage, M.C.; Pineda-Torra, I. Posttranslational modifications of lipid-activated nuclear receptors: Focus on metabolism. Endocrinology 2016, 158, 213–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.; Lee, Y.; Shin, M.; Cho, S.; Zouboulis, C.; Kim, M.; Lee, D.; Chung, J. Histone deacetylase 1 reduces lipogenesis by suppressing SREBP1 transcription in human sebocyte cell line SZ95. Int. J. Mol. Sci. 2021, 22, 4477. [Google Scholar] [CrossRef]
- Chyau, C.-C.; Wang, H.-F.; Zhang, W.-J.; Chen, C.-C.; Huang, S.-H.; Chang, C.-C.; Peng, R.Y. Antrodan alleviates high-fat and high-fructose diet-induced fatty liver disease in C57BL/6 Mice Model via AMPK/Sirt1/SREBP-1c/PPARγ pathway. Int. J. Mol. Sci. 2020, 21, 360. [Google Scholar] [CrossRef] [Green Version]
- Luci, C.; Bourinet, M.; Leclère, P.S.; Anty, R.; Gual, P. Chronic inflammation in non-alcoholic steatohepatitis: Molecular mechanisms and therapeutic strategies. Front. Endocrinol. 2020, 11, 597648. [Google Scholar] [CrossRef]
- Crispe, I.N. Hepatocytes as immunological agents. J. Immunol. 2016, 196, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Arrese, M.; Cabrera, D.; Kalergis, A.M.; Feldstein, A.E. Innate immunity and inflammation in NAFLD/NASH. Dig. Dis. Sci. 2016, 61, 1294–1303. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Zhang, X.-J.; Li, H. Role of innate immune signaling in non-alcoholic fatty liver disease. Trends Endocrinol. Metab. 2018, 29, 712–722. [Google Scholar] [CrossRef]
- Arias-Loste, M.T.; Fábrega, E.; López-Hoyos, M.; Crespo, J. The crosstalk between hypoxia and innate immunity in the development of obesity-related nonalcoholic fatty liver disease. BioMed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Sun, J.; Liu, L.; Du, X.; Liu, Y.; Yan, X.; Osoro, E.K.; Zhang, F.; Feng, L.; Liang, D.; et al. Anti-toll-like receptor 2 antibody ameliorates hepatic injury, inflammation, fibrosis and steatosis in obesity-related metabolic disorder rats via regulating MAPK and NF-κB pathways. Int. Immunopharmacol. 2020, 82, 106368. [Google Scholar] [CrossRef]
- Ahmed, L.A.; Salem, M.B.; Seif El-Din, S.H.; El-Lakkany, N.M.; Ahmed, H.O.; Nasr, S.M.; Hammam, O.A.; Botros, S.S.; Saleh, S. Gut microbiota modulation as a promising therapy with metformin in rats with non-alcoholic steatohepatitis: Role of LPS/TLR4 and autophagy pathways. Eur. J. Pharmacol. 2020, 887, 173461. [Google Scholar] [CrossRef]
- Garcia-Martinez, I.; Santoro, N.; Chen, Y.; Hoque, R.; Ouyang, X.; Caprio, S.; Shlomchik, M.J.; Coffman, R.L.; Candia, A.; Mehal, W.Z. Hepatocyte mitochondrial DNA drives nonalcoholic steatohepatitis by activation of TLR9. J. Clin. Investig. 2016, 126, 859–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mridha, A.R.; Haczeyni, F.; Yeh, M.M.; Haigh, W.G.; Ioannou, G.N.; Barn, V.; Ajamieh, H.; Adams, L.; Hamdorf, J.M.; Teoh, N.C.; et al. TLR9 is up-regulated in human and murine NASH: Pivotal role in inflammatory recruitment and cell survival. Clin. Sci. 2017, 131, 2145–2159. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyoshi, H.; Yasui, K.; Hara, T.; Taketani, H.; Ishiba, H.; Okajima, A.; Seko, Y.; Umemura, A.; Nishikawa, T.; Yamaguchi, K.; et al. Hepatic nucleotide binding oligomerization domain-like receptors pyrin domain-containing 3 inflammasomes are associated with the histologic severity of non-alcoholic fatty liver disease. Hepatol. Res. 2017, 47, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Kubes, P.; Mehal, W.Z. Sterile inflammation in the liver. Gastroenterology 2012, 143, 1158–1172. [Google Scholar] [CrossRef]
- Ibrahim, S.H.; Hirsova, P.; Gores, G.J. Non-alcoholic steatohepatitis pathogenesis: Sublethal hepatocyte injury as a driver of liver inflammation. Gut 2018, 67, 963–972. [Google Scholar] [CrossRef]
- Rangwala, F.; Guy, C.D.; Lu, J.; Suzuki, A.; Burchette, J.L.; Abdelmalek, M.F.; Chen, W.; Diehl, A.M. Increased production of sonic hedgehog by ballooned hepatocytes. J. Pathol. 2011, 224, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Kakazu, E.; Mauer, A.S.; Yin, M.; Malhi, H. Hepatocytes release ceramide-enriched pro-inflammatory extracellular vesicles in an IRE1α-dependent manner. J. Lipid Res. 2016, 57, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, A.N.; Suresh, D.; Santhekadur, P.K.; Suvarna, D.; Kumar, D.P. Extracellular vesicles as inflammatory drivers in NAFLD. Front. Immunol. 2021, 11, 627424. [Google Scholar] [CrossRef] [PubMed]
- Povero, D.; Eguchi, A.; Li, H.; Johnson, C.D.; Papouchado, B.G.; Wree, A.; Messer, K.; Feldstein, A.E. Circulating extracellular vesicles with specific proteome and liver MicroRNAs are potential biomarkers for liver injury in experimental fatty liver disease. PLoS ONE 2014, 9, e113651. [Google Scholar] [CrossRef] [Green Version]
- Kornek, M.; Lynch, M.; Mehta, S.H.; Lai, M.; Exley, M.; Afdhal, N.H.; Schuppan, D. Circulating microparticles as disease-specific biomarkers of severity of inflammation in patients with Hepatitis C or nonalcoholic steatohepatitis. Gastroenterology 2012, 143, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Ariel, E.F.; Feldstein, E.A.; Gores, G.J. Apoptosis in alcoholic and nonalcoholic steatohepatitis. Front. Biosci. 2005, 10, 3093–3099. [Google Scholar] [CrossRef] [Green Version]
- Canbay, A.; Feldstein, A.E.; Higuchi, H.; Werneburg, N.; Grambihler, A.; Bronk, S.F. Kupffer cell engulfment of apoptotic bodies stimulates death ligand and cytokine expression. Hepatology 2003, 38, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Tsurusaki, S.; Tsuchiya, Y.; Koumura, T.; Nakasone, M.; Sakamoto, T.; Matsuoka, M.; Imai, H.; Kok, C.Y.-Y.; Okochi, H.; Nakano, H.; et al. Hepatic ferroptosis plays an important role as the trigger for initiating inflammation in nonalcoholic steatohepatitis. Cell Death Dis. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Gaul, S.; Leszczynska, A.; Alegre, F.; Kaufmann, B.; Johnson, C.D.; Adams, L.A.; Wree, A.; Damm, G.; Seehofer, D.; Calvente, C.J.; et al. Hepatocyte pyroptosis and release of inflammasome particles induce stellate cell activation and liver fibrosis. J. Hepatol. 2021, 74, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Gautheron, J.; Gores, G.J.; Rodrigues, C.M. Lytic cell death in metabolic liver disease. J. Hepatol. 2020, 73, 394–408. [Google Scholar] [CrossRef] [PubMed]
- Reilly, S.M.; Saltiel, A.R. Adapting to obesity with adipose tissue inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic adipose tissue inflammation linking obesity to insulin resistance and Type 2 diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Du Plessis, J.; Van Pelt, J.; Korf, H.; Mathieu, C.; Van Der Schueren, B.; Lannoo, M.; Oyen, T.; Topal, B.; Fetter, G.; Nayler, S.; et al. Association of adipose tissue inflammation with histologic severity of nonalcoholic fatty liver disease. Gastroenterology 2015, 149, 635–648.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijnen, M.; Josefs, T.; Cuijpers, I.; Maalsen, C.J.; Van De Gaar, J.; Vroomen, M.; Wijnands, E.; Rensen, S.S.; Greve, J.W.M.; Hofker, M.H.; et al. Adipose tissue macrophages induce hepatic neutrophil recruitment and macrophage accumulation in mice. Gut 2017, 67, 1317–1327. [Google Scholar] [CrossRef]
- Divella, R.; Mazzocca, A.; Daniele, A.; Sabbà, C.; Paradiso, A. Obesity, nonalcoholic fatty liver disease and adipocytokines network in promotion of cancer. Int. J. Biol. Sci. 2019, 15, 610–616. [Google Scholar] [CrossRef] [Green Version]
- Polyzos, S.A.; Toulis, K.A.; Goulis, D.G.; Zavos, C.; Kountouras, J. Serum total adiponectin in nonalcoholic fatty liver disease: A systematic review and meta-analysis. Metabolism 2011, 60, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Aronis, K.; Kountouras, J.; Raptis, D.D.; Vasiloglou, M.; Mantzoros, C.S. Circulating leptin in non-alcoholic fatty liver disease: A systematic review and meta-analysis. Diabetologia 2016, 59, 30–43. [Google Scholar] [CrossRef]
- Salman, A.A.; Sultan, A.A.E.A.; Abdallah, A.; Abdelsalam, A.; Mikhail, H.; Tourky, M.; Omar, M.; Youssef, A.; Ahmed, R.A.; ElKassar, H.; et al. Effect of weight loss induced by laparoscopic sleeve gastrectomy on liver histology and serum adipokine levels. J. Gastroenterol. Hepatol. 2020, 35, 1769–1773. [Google Scholar] [CrossRef]
- Iruzubieta, P.; Medina, J.M.; Fernández-López, R.; Crespo, J.; De La Cruz, F. A role for gut microbiome fermentative pathways in fatty liver disease progression. J. Clin. Med. 2020, 9, 1369. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut–liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Martín-Mateos, R.; Albillos, A. The role of the gut-liver axis in metabolic dysfunction-associated fatty liver disease. Front. Immunol. 2021, 12, 660179. [Google Scholar] [CrossRef] [PubMed]
- Sharpton, S.R.; Yong, G.J.; Terrault, N.A.; Lynch, S.V. Gut microbial metabolism and nonalcoholic fatty liver disease. Hepatol. Commun. 2019, 3, 29–43. [Google Scholar] [CrossRef]
- Chen, P.; Stärkel, P.; Turner, J.R.; Ho, S.B.; Schnabl, B. Dysbiosis-induced intestinal inflammation activates tumor necrosis factor receptor I and mediates alcoholic liver disease in mice. Hepatology 2015, 61, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef]
- Sharpton, S.R.; Ajmera, V.; Loomba, R. Emerging role of the gut microbiome in nonalcoholic fatty liver disease: From composition to function. Clin. Gastroenterol. Hepatol. 2019, 17, 296–306. [Google Scholar] [CrossRef]
- Lee, G.; You, H.J.; Bajaj, J.S.; Joo, S.K.; Yu, J.; Park, S.; Kang, H.; Park, J.H.; Kim, J.H.; Lee, D.H.; et al. Distinct signatures of gut microbiome and metabolites associated with significant fibrosis in non-obese NAFLD. Nat. Commun. 2020, 11, 4982. [Google Scholar] [CrossRef] [PubMed]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.; Fizanne, L.; Araujo-Perez, F.; Guy, C.D.; Seed, P.C.; Rawls, J.F.; David, L.A.; et al. The severity of nonalcoholic fatty liver disease is associated with gut dysbiosis and shift in the metabolic function of the gut microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut microbiome-based metagenomic signature for non-invasive detection of advanced fibrosis in human nonalcoholic fatty liver disease. Cell Metab. 2017, 25, 1054–1062. [Google Scholar] [CrossRef]
- Caussy, C.; Hsu, C.; Min-Tzu, L.; Amy, L.; Bettencourt, R.; Ajmera, V.; Shirin, B.; Hooker, J.; Ethan, S.; Richards, L.; et al. Novel link between gut-microbiome derived metabolite and shared gene-effects with hepatic steatosis and fibrosis in NAFLD. J. Hepatol. 2018, 68, S29. [Google Scholar] [CrossRef]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clément, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xu, M.-J.; Gao, B. Hepatocytes: A key cell type for innate immunity. Cell. Mol. Immunol. 2016, 13, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Norris, C.A.; He, M.; Kang, L.-I.; Ding, M.; Radder, J.; Haynes, M.M.; Yang, Y.; Paranjpe, S.; Bowen, W.C.; Orr, A.; et al. Synthesis of IL-6 by hepatocytes is a normal response to common hepatic stimuli. PLoS ONE 2014, 9, e96053. [Google Scholar] [CrossRef] [Green Version]
- Jia, Q.; Li, C.; Xia, Y.; Zhang, Q.; Wu, H.; Du, H.; Liu, L.; Wang, C.; Shi, H.; Guo, X.; et al. Association between complement C3 and prevalence of fatty liver disease in an adult population: A cross-sectional study from the Tianjin chronic low-grade systemic inflammation and health (TCLSIHealth) cohort study. PLoS ONE 2015, 10, e0122026. [Google Scholar] [CrossRef]
- Rensen, S.S.; Slaats, Y.; Driessen, A.; Peutz-Kootstra, C.J.; Nijhuis, J.; Steffensen, R.; Greve, J.W.; Buurman, W.A. Activation of the complement system in human nonalcoholic fatty liver disease. Hepatology 2009, 50, 1809–1817. [Google Scholar] [CrossRef]
- Pandey, E.; Nour, A.S.; Harris, E.N. Prominent receptors of liver sinusoidal endothelial cells in liver homeostasis and disease. Front. Physiol. 2020, 11, 873. [Google Scholar] [CrossRef]
- Furuta, K.; Guo, Q.; Pavelko, K.D.; Lee, J.-H.; Robertson, K.D.; Nakao, Y.; Melek, J.; Shah, V.H.; Hirsova, P.; Ibrahim, S.H. Lipid-induced endothelial vascular cell adhesion molecule 1 promotes nonalcoholic steatohepatitis pathogenesis. J. Clin. Investig. 2021, 131, e143690. [Google Scholar] [CrossRef]
- Weston, C.J.; Shepherd, E.L.; Claridge, L.C.; Rantakari, P.; Curbishley, S.M.; Tomlinson, J.W.; Hübscher, S.G.; Reynolds, G.M.; Aalto, K.; Anstee, Q.M.; et al. Vascular adhesion protein-1 promotes liver inflammation and drives hepatic fibrosis. J. Clin. Investig. 2014, 125, 501–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, K.; Guo, Q.; Hirsova, P.; Ibrahim, S.H. Emerging roles of liver sinusoidal endothelial cells in nonalcoholic steatohepatitis. Biology 2020, 9, 395. [Google Scholar] [CrossRef] [PubMed]
- Kazankov, K.; Jørgensen, S.M.D.; Thomsen, K.L.; Møller, H.J.; Vilstrup, H.; George, J.; Schuppan, D.; Grønbæk, H. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 145–159. [Google Scholar] [CrossRef]
- Gadd, V.L.; Skoien, R.; Powell, E.; Fagan, K.J.; Winterford, C.; Horsfall, L.; Irvine, K.; Clouston, A.D. The portal inflammatory infiltrate and ductular reaction in human nonalcoholic fatty liver disease. Hepatology 2014, 59, 1393–1405. [Google Scholar] [CrossRef]
- Krenkel, O.; Tacke, F. Liver macrophages in tissue homeostasis and disease. Nat. Rev. Immunol. 2017, 17, 306–321. [Google Scholar] [CrossRef]
- Pan, J.; Ou, Z.; Cai, C.; Li, P.; Gong, J.; Ruan, X.Z.; He, K. Fatty acid activates NLRP3 inflammasomes in mouse Kupffer cells through mitochondrial DNA release. Cell. Immunol. 2018, 332, 111–120. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Y.; An, W.; Song, J.; Zhang, Y.; Zhao, X. STING-mediated inflammation in Kupffer cells contributes to progression of nonalcoholic steatohepatitis. J. Clin. Investig. 2019, 129, 546–555. [Google Scholar] [CrossRef]
- Miura, K.; Yang, L.; van Rooijen, N.; Brenner, D.A.; Ohnishi, H.; Seki, E. Toll-like receptor 2 and palmitic acid cooperatively contribute to the development of nonalcoholic steatohepatitis through inflammasome activation in mice. Hepatology 2013, 57, 577–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, J.; Benkdane, M.; Teixeira-Clerc, F.; Bonnafous, S.; Louvet, A.; Lafdil, F.; Pecker, F.; Tran, A.; Gual, P.; Mallat, A.; et al. M2 Kupffer cells promote M1 Kupffer cell apoptosis: A protective mechanism against alcoholic and nonalcoholic fatty liver disease. Hepatology 2014, 59, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wang, Q.; Qi, M.; Zhang, C.; Li, Z.; Zhang, W. Ghrelin ameliorates nonalcoholic steatohepatitis induced by chronic low-grade inflammation via blockade of Kupffer cell M1 polarization. J. Cell. Physiol. 2020, 236, 5121–5133. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ni, X.; Xu, Q.; Wang, Q.; Li, X.; Hua, J. Regulation of lipid-induced macrophage polarization through modulating peroxisome proliferator-activated receptor-gamma activity affects hepatic lipid metabolism via a Toll-like receptor 4/NF-κB signaling pathway. J. Gastroenterol. Hepatol. 2020, 35, 1998–2008. [Google Scholar] [CrossRef]
- Li, H.; Zhou, Y.; Wang, H.; Zhang, M.; Qiu, P.; Zhang, R.; Zhao, Q.; Liu, J. Crosstalk between liver macrophages and surrounding cells in nonalcoholic steatohepatitis. Front. Immunol. 2020, 11, 1169. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.T.; Reyes, J.L.; McDonald, B.A.; Vo, T.; Reimer, R.A.; Eksteen, B. Kupffer cells undergo fundamental changes during the development of experimental NASH and are critical in initiating liver damage and inflammation. PLoS ONE 2016, 11, e0159524. [Google Scholar] [CrossRef] [Green Version]
- Remmerie, A.; Martens, L.; Thoné, T.; Castoldi, A.; Seurinck, R.; Pavie, B.; Roels, J.; Vanneste, B.; De Prijck, S.; Vanhockerhout, M.; et al. Osteopontin expression identifies a subset of recruited macrophages distinct from Kupffer cells in the fatty liver. Immunity 2020, 53, 641–657.e14. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jeong, J.-M.; Kim, S.J.; Seo, W.; Kim, M.-H.; Choi, W.-M.; Yoo, W.; Lee, J.-H.; Shim, Y.-R.; Yi, H.-S.; et al. Pro-inflammatory hepatic macrophages generate ROS through NADPH oxidase 2 via endocytosis of monomeric TLR4–MD2 complex. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Glass, O.; Henao, R.; Patel, K.; Guy, C.D.; Gruss, H.J.; Syn, W.-K.; Moylan, C.A.; Streilein, R.; Hall, R.; Diehl, A.M.; et al. Serum Interleukin-8, Osteopontin, and Monocyte Chemoattractant protein 1 are associated with hepatic fibrosis in patients with nonalcoholic fatty liver disease. Hepatol. Commun. 2018, 2, 1344–1355. [Google Scholar] [CrossRef]
- Antonucci, L.; Porcu, C.; Timperi, E.; Santini, S.J.; Iannucci, G.; Balsano, C. Circulating neutrophils of nonalcoholic steatohepatitis patients show an activated phenotype and suppress T lymphocytes activity. J. Immunol. Res. 2020, 2020, 1–15. [Google Scholar] [CrossRef]
- Khoury, T.; Mari, A.; Nseir, W.; Kadah, A.; Sbeit, W.; Mahamid, M. Neutrophil-to-lymphocyte ratio is independently associated with inflammatory activity and fibrosis grade in nonalcoholic fatty liver disease. Eur. J. Gastroenterol. Hepatol. 2019, 31, 1110–1115. [Google Scholar] [CrossRef]
- Hwang, S.; He, Y.; Xiang, X.; Seo, W.; Kim, S.; Ma, J.; Ren, T.; Park, S.H.; Zhou, Z.; Feng, D.; et al. Interleukin-22 Ameliorates neutrophil-driven nonalcoholic steatohepatitis through multiple targets. Hepatology 2019, 72, 412–429. [Google Scholar] [CrossRef] [PubMed]
- Ou, R.; Liu, J.; Lv, M.; Wang, J.; Wang, J.; Zhu, L.; Zhao, L.; Xu, Y. Neutrophil depletion improves diet-induced non-alcoholic fatty liver disease in mice. Endocrine 2017, 57, 72–82. [Google Scholar] [CrossRef]
- Rensen, S.S.; Slaats, Y.; Nijhuis, J.; Jans, A.; Bieghs, V.; Driessen, A.; Malle, E.; Greve, J.W.; Buurman, W.A. Increased hepatic myeloperoxidase activity in obese subjects with nonalcoholic steatohepatitis. Am. J. Pathol. 2009, 175, 1473–1482. [Google Scholar] [CrossRef] [Green Version]
- Rensen, S.S.; Bieghs, V.; Xanthoulea, S.; Arfianti, E.; Bakker, J.A.; Shiri-Sverdlov, R.; Hofker, M.H.; Greve, J.W.; Buurman, W.A. Neutrophil-derived myeloperoxidase aggravates non-alcoholic steatohepatitis in low-density lipoprotein receptor-deficient mice. PLoS ONE 2012, 7, e52411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulli, B.; Ali, M.; Iwamoto, Y.; Zeller, M.W.; Schob, S.; Linnoila, J.J.; Chen, J.W. Myeloperoxidase–hepatocyte–stellate cell cross talk promotes hepatocyte injury and fibrosis in experimental nonalcoholic steatohepatitis. Antioxid. Redox Signal. 2015, 23, 1255–1269. [Google Scholar] [CrossRef] [Green Version]
- Mirea, A.-M.; Toonen, E.J.M.; Munckhof, I.V.D.; Munsterman, I.D.; Tjwa, E.T.T.L.; Jaeger, M.; Oosting, M.; Schraa, K.; Rutten, J.H.W.; Van Der Graaf, M.; et al. Increased proteinase 3 and neutrophil elastase plasma concentrations are associated with non-alcoholic fatty liver disease (NAFLD) and type 2 diabetes. Mol. Med. 2019, 25, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Liang, B.; Bian, D.; Luo, Y.; Yang, J.; Li, Z.; Zhuang, Z.; Zang, S.; Shi, J. Knockout of neutrophil elastase protects against western diet induced nonalcoholic steatohepatitis in mice by regulating hepatic ceramides metabolism. Biochem. Biophys. Res. Commun. 2019, 518, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Yang, K.; Zang, S.; Lin, Z.; Chau, H.T.; Wang, Y.; Wang, Y. Lipocalin-2 mediates non-alcoholic steatohepatitis by promoting neutrophil-macrophage crosstalk via the induction of CXCR2. J. Hepatol. 2016, 65, 988–997. [Google Scholar] [CrossRef]
- van der Windt, D.J.; Sud, V.; Zhang, H.; Varley, P.R.; Goswami, J.; Yazdani, H.O.; Tohme, S.; Loughran, P.; O’Doherty, R.M.; Minervini, M.I.; et al. Neutrophil extracellular traps promote inflammation and development of hepatocellular carcinoma in nonalcoholic steatohepatitis. Hepatology 2018, 68, 1347–1360. [Google Scholar] [CrossRef] [Green Version]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate lymphoid cells: 10 years on. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shen, J.; Man, K.; Chu, E.S.; Yau, T.O.; Sung, J.C.; Go, M.Y.; Deng, J.; Lu, L.; Wong, V.W.; et al. CXCL10 plays a key role as an inflammatory mediator and a non-invasive biomarker of non-alcoholic steatohepatitis. J. Hepatol. 2014, 61, 1365–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiglund, N.; Strand, K.; Cornillet, M.; Stål, P.; Thorell, A.; Zimmer, C.L.; Näslund, E.; Karlgren, S.; Nilsson, H.; Mellgren, G.; et al. Retained NK cell phenotype and functionality in non-alcoholic fatty liver disease. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Kahraman, A.; Schlattjan, M.; Kocabayoglu, P.; Yildiz-Meziletoglu, S.; Schlensak, M.; Fingas, C.D.; Wedemeyer, I.; Marquitan, G.; Gieseler, R.K.; Baba, H.A.; et al. Major histocompatibility complex class I-related chains A and B (MIC A/B): A novel role in nonalcoholic steatohepatitis. Hepatology 2010, 51, 92–102. [Google Scholar] [CrossRef]
- Cuff, A.O.; Sillito, F.; Dertschnig, S.; Hall, A.; Luong, T.V.; Chakraverty, R.; Male, V. The obese liver environment mediates conversion of NK cells to a less cytotoxic ILC1-Like phenotype. Front. Immunol. 2019, 10, 2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Santos, L.; Luka, Z.; Wagner, C.; Fernandez-Alvarez, S.; Lu, S.C.; Mato, J.M.; Martinez-Chantar, M.L.; Beraza, N. Inhibition of natural killer cells protects the liver against acute injury in the absence of glycine N-methyltransferase. Hepatology 2012, 56, 747–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosello-Trampont, A.-C.; Krueger, P.; Narayanan, S.; Landes, S.G.; Leitinger, N.; Hahn, Y.S. NKp46+ natural killer cells attenuate metabolism-induced hepatic fibrosis by regulating macrophage activation in mice. Hepatology 2016, 63, 799–812. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Zhang, W.; Wei, H.; Sun, R.; Tian, Z.; Chen, Y. Hepatic NK cells attenuate fibrosis progression of non-alcoholic steatohepatitis in dependent of CXCL10-mediated recruitment. Liver Int. 2019, 40, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.I.; Di Santo, J.P. ILC-poiesis: Ensuring tissue ILC differentiation at the right place and time. Eur. J. Immunol. 2019, 49, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Hendrikx, T.; Binder, C.J. Oxidation-specific epitopes in non-alcoholic fatty liver disease. Front. Endocrinol. 2020, 11, 607011. [Google Scholar] [CrossRef]
- Bruzzi’, S.; Sutti, S.; Giudici, G.; Burlone, M.E.; Ramavath, N.N.; Toscani, A.; Bozzola, C.; Schneider, P.; Morello, E.; Parola, M.; et al. B2-Lymphocyte responses to oxidative stress-derived antigens contribute to the evolution of nonalcoholic fatty liver disease (NAFLD). Free. Radic. Biol. Med. 2018, 124, 249–259. [Google Scholar] [CrossRef]
- Zhang, F.; Jiang, W.W.; Li, X.; Qiu, X.Y.; Wu, Z.; Chi, Y.J.; Cong, X.; Liu, Y.L. Role of intrahepatic B cells in non-alcoholic fatty liver disease by secreting pro-inflammatory cytokines and regulating intrahepatic T cells. J. Dig. Dis. 2016, 17, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Barrow, F.; Khan, S.; Fredrickson, G.; Wang, H.; Dietsche, K.; Parthiban, P.; Robert, S.; Kaiser, T.; Winer, S.; Herman, A.; et al. Microbiota-driven activation of intrahepatic B cells aggravates NASH through innate and adaptive signaling. Hepatology 2021, 74, 704–722. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.P.; Liu, Y.; Iyer, S.S.; Liu, S.; Gupta, B.; Desai, C.; Kumar, P.; Smith, T.; Singhi, A.D.; Nusrat, A.; et al. Blocking integrin α4β7-mediated CD4 T cell recruitment to the intestine and liver protects mice from western diet-induced non-alcoholic steatohepatitis. J. Hepatol. 2020, 73, 1013–1022. [Google Scholar] [CrossRef]
- Ma, C.; Kesarwala, A.; Eggert, T.; Medina-Echeverz, J.; Kleiner, D.E.; Jin, P.; Stroncek, P.J.D.F.; Terabe, M.; Kapoor, V.; Elgindi, M.; et al. NAFLD causes selective CD4+ T lymphocyte loss and promotes hepatocarcinogenesis. Nat. Cell Biol. 2016, 531, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Haas, J.T.; Vonghia, L.; Mogilenko, D.A.; Verrijken, A.; Molendi-Coste, O.; Fleury, S.; DePrince, A.; Nikitin, A.; Woitrain, E.; Ducrocq-Geoffroy, L.; et al. Transcriptional network analysis implicates altered hepatic immune function in NASH development and resolution. Nat. Metab. 2019, 1, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Solari, N.E.F.; Inzaugarat, M.E.; Baz, P.; De Matteo, E.; Lezama, C.; Galoppo, M.; Galoppo, C.; Cherñavsky, A.C. The role of innate cells is coupled to a Th1-polarized immune response in pediatric nonalcoholic steatohepatitis. J. Clin. Immunol. 2012, 32, 611–621. [Google Scholar] [CrossRef]
- Luo, X.-Y.; Takahara, T.; Kawai, K.; Fujino, M.; Sugiyama, T.; Tsuneyama, K.; Tsukada, K.; Nakae, S.; Zhong, L.; Li, X.-K. IFN-γ deficiency attenuates hepatic inflammation and fibrosis in a steatohepatitis model induced by a methionine- and choline-deficient high-fat diet. Am. J. Physiol. Liver Physiol. 2013, 305, G891–G899. [Google Scholar] [CrossRef] [Green Version]
- Diedrich, T.; Kummer, S.; Galante, A.; Drolz, A.; Schlicker, V.; Lohse, A.W.; Kluwe, J.; Eberhard, J.M.; Wiesch, J.S.Z. Characterization of the immune cell landscape of patients with NAFLD. PLoS ONE 2020, 15, e0230307. [Google Scholar] [CrossRef] [Green Version]
- Rau, M.; Schilling, A.-K.; Meertens, J.; Hering, I.; Weiss, J.; Jurowich, C.; Kudlich, T.; Hermanns, H.M.; Bantel, H.; Beyersdorf, N.; et al. Progression from nonalcoholic fatty liver to nonalcoholic steatohepatitis is marked by a higher frequency of Th17 cells in the liver and an increased Th17/resting regulatory t cell ratio in peripheral blood and in the liver. J. Immunol. 2015, 196, 97–105. [Google Scholar] [CrossRef]
- Tang, Y.; Bian, Z.; Zhao, L.; Liu, Y.; Liang, S.; Wang, Q.; Han, X.; Peng, Y.; Chen, X.; Shen, L.; et al. Interleukin-17 exacerbates hepatic steatosis and inflammation in non-alcoholic fatty liver disease. Clin. Exp. Immunol. 2011, 166, 281–290. [Google Scholar] [CrossRef]
- Harley, I.T.; Stankiewicz, T.E.; Giles, D.A.; Softic, S.; Flick, L.M.; Cappelletti, M.; Sheridan, R.; Xanthakos, S.A.; Steinbrecher, K.A.; Sartor, R.B.; et al. IL-17 signaling accelerates the progression of nonalcoholic fatty liver disease in mice. Hepatology 2014, 59, 1830–1839. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; She, W.; Wang, F.; Li, J.; Wang, J.; Jiang, W. 3, 3′-diindolylmethane alleviates steatosis and the progression of NASH partly through shifting the imbalance of Treg/Th17 cells to Treg dominance. Int. Immunopharmacol. 2014, 23, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Wang, K.; Aoyama, T.; Grivennikov, S.I.; Paik, Y.; Scholten, D.; Cong, M.; Iwaisako, K.; Liu, X.; Zhang, M.; et al. Interleukin-17 signaling in inflammatory, kupffer cells, and hepatic stellate cells exacerbates liver fibrosis in mice. Gastroenterology 2012, 143, 765–776.e3. [Google Scholar] [CrossRef] [Green Version]
- Rolla, S.; Alchera, E.; Imarisio, C.; Bardina, V.; Valente, G.; Cappello, P.; Mombello, C.; Follenzi, A.; Novelli, F.; Carini, R. The balance between IL-17 and IL-22 produced by liver-infiltrating T-helper cells critically controls NASH development in mice. Clin. Sci. 2015, 130, 193–203. [Google Scholar] [CrossRef]
- Söderberg, C.; Marmur, J.; Eckes, K.; Glaumann, H.; Sällberg, M.; Frelin, L.; Rosenberg, P.; Stål, P.; Hultcrantz, R. Microvesicular fat, inter cellular adhesion molecule-1 and regulatory T-lymphocytes are of importance for the inflammatory process in livers with non-alcoholic steatohepatitis. APMIS 2011, 119, 412–420. [Google Scholar] [CrossRef] [Green Version]
- He, B.; Wu, L.; Xie, W.; Shao, Y.; Jiang, J.; Zhao, Z.; Yan, M.; Chen, Z.; Cui, D. The imbalance of Th17/Treg cells is involved in the progression of nonalcoholic fatty liver disease in mice. BMC Immunol. 2017, 18, 33. [Google Scholar] [CrossRef] [Green Version]
- Ghazarian, M.; Revelo, X.S.; Nøhr, M.K.; Luck, H.; Zeng, K.; Lei, H.; Tsai, S.; Schroer, S.A.; Park, Y.J.; Chng, M.H.Y.; et al. Type I interferon responses drive intrahepatic T cells to promote metabolic syndrome. Sci. Immunol. 2017, 2, 7616. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef]
- Wang, T.; Sun, G.; Wang, Y.; Li, S.; Zhao, X.; Zhang, C.; Jin, H.; Tian, D.; Liu, K.; Shi, W.; et al. The immunoregulatory effects of CD8 T-cell–derived perforin on diet-induced nonalcoholic steatohepatitis. FASEB J. 2019, 33, 8490–8503. [Google Scholar] [CrossRef]
- Inzaugarat, M.E.; Solari, N.E.F.; Billordo, L.A.; Abecasis, R.; Gadano, A.C.; Cherñavsky, A.C. Altered phenotype and functionality of circulating immune cells characterize adult patients with nonalcoholic steatohepatitis. J. Clin. Immunol. 2011, 31, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Adler, M. Intrahepatic natural killer T cell populations are increased in human hepatic steatosis. World J. Gastroenterol. 2011, 17, 1725–1731. [Google Scholar] [CrossRef]
- Maricic, I.; Marrero, I.; Eguchi, A.; Nakamura, R.; Johnson, C.D.; Dasgupta, S.; Hernandez, C.D.; Nguyen, P.S.; Swafford, A.D.; Knight, R.; et al. Differential activation of hepatic invariant NKT cell subsets plays a key role in progression of nonalcoholic steatohepatitis. J. Immunol. 2018, 201, 3017–3035. [Google Scholar] [CrossRef] [Green Version]
- Tajiri, K.; Shimizu, Y.; Tsuneyama, K.; Sugiyama, T. Role of liver-infiltrating CD3+CD56+ natural killer T cells in the pathogenesis of nonalcoholic fatty liver disease. Eur. J. Gastroenterol. Hepatol. 2009, 21, 673–680. [Google Scholar] [CrossRef]
- Syn, W.-K.; Oo, Y.H.; Pereira, T.A.; Karaca, G.F.; Jung, Y.; Omenetti, A.; Witek, R.P.; Choi, S.S.; Guy, C.D.; Fearing, C.M.; et al. Accumulation of natural killer T cells in progressive nonalcoholic fatty liver disease. Hepatology 2010, 51, 1998–2007. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, J.; Kirby, M.; Softic, S.; Miles, L.; Salazar-Gonzalez, R.-M.; Shivakumar, P.; Kohli, R. Hepatic natural killer T-cell and CD8+ T-cell signatures in mice with nonalcoholic steatohepatitis. Hepatol. Commun. 2017, 1, 299–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyagi, T.; Takehara, T.; Uemura, A.; Nishio, K.; Shimizu, S.; Kodama, T.; Hikita, H.; Li, W.; Sasakawa, A.; Tatsumi, T.; et al. Absence of invariant natural killer T cells deteriorates liver inflammation and fibrosis in mice fed high-fat diet. J. Gastroenterol. 2010, 45, 1247–1254. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Soloski, M.J.; Diehl, A.M. Dietary factors alter hepatic innate immune system in mice with nonalcoholic fatty liver disease. Hepatology 2005, 42, 880–885. [Google Scholar] [CrossRef]
- Kremer, M.; Thomas, E.; Milton, R.J.; Perry, A.W.; van Rooijen, N.; Wheeler, M.D.; Zacks, S.; Fried, M.; Rippe, R.A.; Hines, I.N. Kupffer cell and interleukin-12-dependent loss of natural killer T cells in hepatosteatosis. Hepatology 2010, 51, 130–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolte, F.J.; Rehermann, B. Mucosal-associated invariant T cells in chronic inflammatory liver disease. Semin. Liver Dis. 2018, 38, 060–065. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, L.; Lehuen, A. MAIT cells in metabolic diseases. Mol. Metab. 2019, 27, S114–S121. [Google Scholar] [CrossRef]
- Hegde, P.; Weiss, E.; Paradis, V.; Wan, J.; Mabire, M.; Sukriti, S.; Rautou, P.-E.; Albuquerque, M.; Picq, O.; Gupta, A.C.; et al. Mucosal-associated invariant T cells are a profibrogenic immune cell population in the liver. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Huang, B.; Jiang, X.; Chen, W.; Zhang, J.; Wei, Y.; Chen, Y.; Lian, M.; Bian, Z.; Miao, Q.; et al. Mucosal-associated invariant T cells improve nonalcoholic fatty liver disease through regulating macrophage polarization. Front. Immunol. 2018, 9, 1994. [Google Scholar] [CrossRef]
- Li, F.; Hao, X.; Chen, Y.; Bai, L.; Gao, X.; Lian, Z.; Wei, H.; Sun, R.; Tian, Z. The microbiota maintain homeostasis of liver-resident γδT-17 cells in a lipid antigen/CD1d-dependent manner. Nat. Commun. 2017, 8, 13839. [Google Scholar] [CrossRef] [PubMed]
- Torres-Hernandez, A.; Wang, W.; Nikiforov, Y.; Tejada, K.; Torres, L.; Kalabin, A.; Adam, S.; Wu, J.; Lu, L.; Chen, R.; et al. γδ T cells promote steatohepatitis by orchestrating innate and adaptive immune programming. Hepatology 2019, 71, 477–494. [Google Scholar] [CrossRef]
- Vilar-Gomez, E.; Martinez-Perez, Y.; Calzadilla-Bertot, L.; Torres-Gonzalez, A.; Gra-Oramas, B.; Gonzalez-Fabian, L.; Friedman, S.L.; Diago, M.; Romero-Gomez, M. Weight loss through lifestyle modification significantly reduces features of nonalcoholic steatohepatitis. Gastroenterology 2015, 149, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Francque, S.M.; Marchesini, G.; Kautz, A.; Walmsley, M.; Dorner, R.; Lazarus, J.V.; Zelber-Sagi, S.; Hallsworth, K.; Busetto, L.; Frühbeck, G.; et al. Non-alcoholic fatty liver disease: A patient guideline. JHEP Rep. 2021, 3, 100322. [Google Scholar] [CrossRef]
- Lassailly, G.; Caiazzo, R.; Ntandja-Wandji, L.-C.; Gnemmi, V.; Baud, G.; Verkindt, H.; Ningarhari, M.; Louvet, A.; Leteurtre, E.; Raverdy, V.; et al. Bariatric surgery provides long-term resolution of nonalcoholic steatohepatitis and regression of fibrosis. Gastroenterology 2020, 159, 1290–1301.e5. [Google Scholar] [CrossRef]
- Lavín-Alconero, L.; Fernández-Lanas, T.; Iruzubieta-Coz, P.; Arias-Loste, M.T.; Rodriguez-Duque, J.C.; Rivas, C.; Cagigal, M.L.; Montalbán, C.; Useros, A.L.; Álvarez-Cancelo, A.; et al. Efficacy and safety of endoscopic sleeve gastroplasty versus laparoscopic sleeve gastrectomy in obese subjects with Non-Alcoholic SteatoHepatitis (NASH): Study protocol for a randomized controlled trial (TESLA-NASH study). Trials 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Sharaiha, R.Z.; Kumta, N.A.; Saumoy, M.; Desai, A.P.; Sarkisian, A.M.; Benevenuto, A.; Tyberg, A.; Kumar, R.; Igel, L.; Verna, E.C.; et al. Endoscopic sleeve gastroplasty significantly reduces body mass index and metabolic complications in obese patients. Clin. Gastroenterol. Hepatol. 2017, 15, 504–510. [Google Scholar] [CrossRef]
- Shu, Y.; Lu, Y.; Pang, X.; Zheng, W.; Huang, Y.; Li, J.; Ji, J.; Zhang, C.; Shen, P. Phosphorylation of PPARγ at Ser84 promotes glycolysis and cell proliferation in hepatocellular carcinoma by targeting PFKFB4. Oncotarget 2016, 7, 76984–76994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Colca, J.R.; McDonald, W.G.; McCommis, K.; Finck, B.N. Treating fatty liver disease by modulating mitochondrial pyruvate metabolism. Hepatol. Commun. 2017, 1, 193–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gastaldelli, A.; Cusi, K. From NASH to diabetes and from diabetes to NASH: Mechanisms and treatment options. JHEP Rep. 2019, 1, 312–328. [Google Scholar] [CrossRef] [Green Version]
- Musso, G.; Cassader, M.; Paschetta, E.; Gambino, R. Thiazolidinediones and advanced liver fibrosis in nonalcoholic steatohepatitis. JAMA Intern. Med. 2017, 177, 633–640. [Google Scholar] [CrossRef]
- Ratziu, V.; Charlotte, F.; Bernhardt, C.; Giral, P.; Halbron, M.; Lenaour, G.; Hartmann-Heurtier, A.; Bruckert, E.; Poynard, T. LIDO study group long-term efficacy of rosiglitazone in nonalcoholic steatohepatitis: Results of the fatty liver improvement by rosiglitazone therapy (FLIRT 2) extension trial. Hepatology 2009, 51, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Chalasani, N.; Kowdley, K.V.; McCullough, A.; Diehl, A.M.; Bass, N.M.; Neuschwander-Tetri, B.A.; Lavine, J.E.; Tonascia, J.; Unalp, A.; et al. Pioglitazone, vitamin E, or placebo for nonalcoholic steatohepatitis. N. Engl. J. Med. 2010, 362, 1675–1685. [Google Scholar] [CrossRef] [Green Version]
- Cusi, K.; Orsak, B.; Bril, F.; Lomonaco, R.; Hecht, J.; Ortiz-Lopez, C.; Tio, F.; Hardies, J.; Darland, C.; Musi, N.; et al. Long-term Pioglitazone treatment for patients with nonalcoholic steatohepatitis and prediabetes or type 2 diabetes mellitus. Ann. Intern. Med. 2016, 165, 305–315. [Google Scholar] [CrossRef]
- Chalasani, N.; Younossi, Z.; LaVine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the study of liver diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef]
- Mehtälä, J.; Khanfir, H.; Bennett, D.; Ye, Y.; Korhonen, P.; Hoti, F. Pioglitazone use and risk of bladder cancer: A systematic literature review and meta-analysis of observational studies. Diabetol. Int. 2018, 10, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Viscoli, C.M.; Inzucchi, S.E.; Young, L.H.; Insogna, K.L.; Conwit, R.; Furie, K.L.; Gorman, M.; Kelly, M.A.; Lovejoy, A.M.; Kernan, W.N. Pioglitazone and risk for bone fracture: Safety data from a randomized clinical trial. J. Clin. Endocrinol. Metab. 2016, 102, 914–922. [Google Scholar] [CrossRef] [Green Version]
- Parlati, L.; Régnier, M.; Guillou, H.; Postic, C. New targets for NAFLD. JHEP Rep. 2021, 3, 100346. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Hong, S.H.; Park, Y.-J.; Sung, J.-H.; Suh, W.; Lee, K.W.; Jung, K.; Lim, C.; Kim, J.-H.; Kim, H.; et al. MD001, a novel Peroxisome proliferator-activated receptor α/γ agonist, improves glucose and lipid metabolism. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Francque, S.M.; Bedossa, P.; Ratziu, V.; Anstee, Q.M.; Bugianesi, E.; Sanyal, A.J.; Loomba, R.; Harrison, S.A.; Balabanska, R.; Mateva, L.; et al. A randomized, controlled trial of the pan-PPAR agonist lanifibranor in NASH. N. Engl. J. Med. 2021, 385, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Struik, D.; Dommerholt, M.B.; Jonker, J.W. Fibroblast growth factors in control of lipid metabolism. Curr. Opin. Lipidol. 2019, 30, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Esler, W.P.; Bence, K.K. Metabolic targets in nonalcoholic fatty liver disease. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 247–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanyal, A.; Charles, E.D.; Neuschwander-Tetri, A.B.; Loomba, R.; Harrison, A.S.; Abdelmalek, M.F.; Lawitz, E.J.; Halegoua-DeMarzio, D.; Kundu, S.; Noviello, S.; et al. Pegbelfermin (BMS-986036), a PEGylated fibroblast growth factor 21 analogue, in patients with non-alcoholic steatohepatitis: A randomised, double-blind, placebo-controlled, phase 2a trial. Lancet 2018, 392, 2705–2717. [Google Scholar] [CrossRef]
- Loomba, R.; Sanyal, A.J.; Nakajima, A.; Neuschwander-Tetri, B.A.; Goodman, Z.; Harrison, S.A.; Lawitz, E.J.; Gunn, N.; Imajo, K.; Ravendhran, N.; et al. Efficacy and safety of pegbelfermin in patients with nonalcoholic steatohepatitis and stage 3 fibrosis: Results from the phase 2b, randomized, double-blind, placebo-controlled FALCON 1 study. Hepatology 2021, 74, 9–10. [Google Scholar]
- Shevell, D.E.; Brown, E.; Du, S. Comparison of manual vs machine learning approaches to liver biopsy scoring for NASH and fibrosis: A post hoc analysis of the FALCON 1 study. Hepatology 2021, 74, 65–66. [Google Scholar]
- Sanyal, A.J.; Shevell, D.E.; Brown, E. Effect of pegbelfermin on noninvasive biomarkers of NASH and fibrosis: A post hoc analysis of the FALCON 1 trial. Hepatology 2021, 74, 67–68. [Google Scholar]
- Abdelmalek, M.F.; Sanyal, A.J.; Nakajima, A.; Neuschwander-Tetri, A.B.; Goodman, Z.; Lawitz, E.J.; Harrison, S.A.; Jacobson, I.M.; Imajo, K.; Gunn, N.; et al. Efficacy and safety of pegbelfermin in patients with nonalcoholic steatohepatitis and compensated cirrhosis: Results from the phase 2b, randomized, double-blind, placebo-controlled FALCON 2 study. Hepatology 2021, 74, 27–28. [Google Scholar]
- Harrison, S.A.; Ruane, P.J.; Freilich, B.L.; Neff, G.; Patil, R.; Behling, C.A.; Hu, C.; Fong, E.; de Temple, B.; Tillman, E.J.; et al. Efruxifermin in non-alcoholic steatohepatitis: A randomized, double-blind, placebo-controlled, phase 2a trial. Nat. Med. 2021, 27, 1262–1271. [Google Scholar] [CrossRef]
- Depaoli, A.; Phung, V.; Bashir, M.R.; Morrow, L.; Beysen, C.; Yan, A.; Ling, L.; Baxter, B.; Luskey, K.L.; Olefsky, J.M. 140-LB: NGM313, a novel activator of b-Klotho/FGFR1c, improves insulin resistance and reduces hepatic fat in obese, nondiabetic subjects. Diabetes 2019, 68, 140. [Google Scholar] [CrossRef]
- Lawitz, E.J.; Coste, A.; Poordad, F.; Alkhouri, N.; Loo, N.; McColgan, B.J.; Tarrant, J.M.; Nguyen, T.; Han, L.; Chung, C.; et al. Acetyl-CoA Carboxylase inhibitor GS-0976 for 12 weeks reduces hepatic de novo lipogenesis and steatosis in patients with nonalcoholic steatohepatitis. Clin. Gastroenterol. Hepatol. 2018, 16, 1983–1991.e3. [Google Scholar] [CrossRef]
- Calle, R.A.; Amin, N.B.; Carvajal-Gonzalez, S.; Ross, T.T.; Bergman, A.; Aggarwal, S.; Crowley, C.; Rinaldi, A.; Mancuso, J.; Aggarwal, N.; et al. ACC inhibitor alone or co-administered with a DGAT2 inhibitor in patients with non-alcoholic fatty liver disease: Two parallel, placebo-controlled, randomized phase 2a trials. Nat. Med. 2021, 27, 1836–1848. [Google Scholar] [CrossRef]
- Loomba, R.; Mohseni, R.; Lucas, K.J.; Gutierrez, J.A.; Perry, R.G.; Trotter, J.F.; Rahimi, R.S.; Harrison, S.A.; Ajmera, V.; Wayne, J.D.; et al. TVB-2640 (FASN Inhibitor) for the treatment of nonalcoholic steatohepatitis: FASCINATE-1, a randomized, placebo-controlled phase 2a trial. Gastroenterology 2021, 161, 1475–1486. [Google Scholar] [CrossRef]
- Ratziu, V.; de Guevara, L.; Safadi, R.; Poordad, F.; Fuster, F.; Flores-Figueroa, J.; Arrese, M.; Fracanzani, A.L.; Ben Bashat, D.; Lackner, K.; et al. Aramchol in patients with nonalcoholic steatohepatitis: A randomized, double-blind, placebo-controlled phase 2b trial. Nat. Med. 2021, 27, 1825–1835. [Google Scholar] [CrossRef]
- Ajmera, V.H.; Cachay, E.; Ramers, C.; Vodkin, I.; Bassirian, S.; Singh, S.; Mangla, N.; Bettencourt, R.; Aldous, J.L.; Park, D.; et al. MRI assessment of treatment response in HIV-associated NAFLD: A randomized trial of a stearoyl-coenzyme-a-desaturase-1 inhibitor (ARRIVE Trial). Hepatology 2019, 70, 1531–1545. [Google Scholar] [CrossRef] [PubMed]
- Kucsera, D.; Tóth, V.E.; Gergő, D.; Vörös, I.; Onódi, Z.; Görbe, A.; Ferdinandy, P.; Varga, Z.V. Characterization of the CDAA diet-induced non-alcoholic steatohepatitis model: Sex-specific differences in inflammation, fibrosis, and cholesterol metabolism in middle-aged mice. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Vilar-Gomez, E.; Gawrieh, S.; Liang, T.; McIntyre, A.D.; Hegele, R.A.; Chalasani, N. Interrogation of selected genes influencing serum LDL-Cholesterol levels in patients with well characterized NAFLD. J. Clin. Lipidol. 2020, 15, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Dongiovanni, P.; Paolini, E.; Corsini, A.; Sirtori, C.R.; Ruscica, M. Nonalcoholic fatty liver disease or metabolic dysfunction-associated fatty liver disease diagnoses and cardiovascular diseases: From epidemiology to drug approaches. Eur. J. Clin. Investig. 2021, 51, e13519. [Google Scholar] [CrossRef]
- Scicali, R.; Di Pino, A.; Urbano, F.; Ferrara, V.; Marchisello, S.; Di Mauro, S.; Scamporrino, A.; Filippello, A.; Rabuazzo, A.M.; Purrello, F.; et al. Analysis of steatosis biomarkers and inflammatory profile after adding on PCSK9 inhibitor treatment in familial hypercholesterolemia subjects with nonalcoholic fatty liver disease: A single lipid center real-world experience. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 869–879. [Google Scholar] [CrossRef]
- Marra, F.; Svegliati-Baroni, G. Lipotoxicity and the gut-liver axis in NASH pathogenesis. J. Hepatol. 2018, 68, 280–295. [Google Scholar] [CrossRef]
- Neuschwander-Tetri, B.A.; Loomba, R.; Sanyal, A.J.; Lavine, J.E.; Van Natta, M.L.; Abdelmalek, M.F.; Chalasani, N.; Dasarathy, S.; Diehl, A.M.; Hameed, B.; et al. Farnesoid X nuclear receptor ligand obeticholic acid for non-cirrhotic, non-alcoholic steatohepatitis (FLINT): A multicentre, randomised, placebo-controlled trial. Lancet 2015, 385, 956–965. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.; Harrison, S.A.; Elkhashab, M.; Trotter, J.F.; Herring, R.; Rojter, S.E.; Kayali, Z.; Wong, V.W.-S.; Greenbloom, S.; Jayakumar, S.; et al. Cilofexor, a nonsteroidal FXR agonist, in patients with noncirrhotic NASH: A phase 2 randomized controlled trial. Hepatology 2020, 72, 58–71. [Google Scholar] [CrossRef]
- Loomba, R.; Noureddin, M.; Kowdley, K.V.; Kohli, A.; Sheikh, A.; Neff, G.; Bhandari, B.R.; Gunn, N.; Caldwell, S.H.; Goodman, Z.; et al. Combination therapies including cilofexor and firsocostat for bridging fibrosis and cirrhosis attributable to NASH. Hepatology 2021, 73, 625–643. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.S.; Bril, F.; Cusi, K.; Newsome, P.N. Modulation of insulin resistance in nonalcoholic fatty liver disease. Hepatology 2019, 70, 711–724. [Google Scholar] [CrossRef]
- Macauley, M.; Hollingsworth, K.G.; Smith, F.E.; Thelwall, P.E.; Al-Mrabeh, A.; Schweizer, A.; Foley, J.E.; Taylor, R. Effect of vildagliptin on hepatic steatosis. J. Clin. Endocrinol. Metab. 2015, 100, 1578–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.A.; Mells, J.; Dunham, R.M.; Grakoui, A.; Handy, J.; Saxena, N.K.; Anania, F.A. Glucagon-like peptide-1 receptor is present on human hepatocytes and has a direct role in decreasing hepatic steatosis in vitro by modulating elements of the insulin signaling pathway. Hepatology 2010, 51, 1584–1592. [Google Scholar] [CrossRef] [Green Version]
- Htike, Z.Z.; Zaccardi, F.; Papamargaritis, D.; Webb, D.R.; Khunti, K.; Davies, M. Efficacy and safety of glucagon-like peptide-1 receptor agonists in type 2 diabetes: A systematic review and mixed-treatment comparison analysis. Diabetes Obes. Metab. 2017, 19, 524–536. [Google Scholar] [CrossRef] [Green Version]
- Rowlands, J.; Heng, J.; Newsholme, P.; Carlessi, R. Pleiotropic effects of GLP-1 and analogs on cell signaling, metabolism, and function. Front. Endocrinol. 2018, 9, 672. [Google Scholar] [CrossRef] [Green Version]
- Bifari, F.; Manfrini, R.; Cas, M.D.; Berra, C.; Siano, M.; Zuin, M.; Paroni, R.; Folli, F. Multiple target tissue effects of GLP-1 analogues on non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH). Pharmacol. Res. 2018, 137, 219–229. [Google Scholar] [CrossRef]
- Armstrong, M.; Gaunt, P.; Aithal, G.; Barton, D.; Hull, D.; Parker, R.; Hazlehurst, J.M.; Guo, K.; Abouda, G.; Aldersley, M.A.; et al. Liraglutide safety and efficacy in patients with non-alcoholic steatohepatitis (LEAN): A multicentre, double-blind, randomised, placebo-controlled phase 2 study. Lancet 2016, 387, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Newsome, P.N.; Buchholtz, K.; Cusi, K.; Linder, M.; Okanoue, T.; Ratziu, V.; Sanyal, A.J.; Sejling, A.-S.; Harrison, S.A. A placebo-controlled trial of subcutaneous semaglutide in nonalcoholic steatohepatitis. N. Engl. J. Med. 2021, 384, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Sorli, C.; Harashima, S.-I.; Tsoukas, G.M.; Unger, J.; Karsbøl, J.D.; Hansen, T.; Bain, S.C. Efficacy and safety of once-weekly semaglutide monotherapy versus placebo in patients with type 2 diabetes (SUSTAIN 1): A double-blind, randomised, placebo-controlled, parallel-group, multinational, multicentre phase 3a trial. Lancet Diabetes Endocrinol. 2017, 5, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Ahrén, B.; Masmiquel, L.; Kumar, H.; Sargin, M.; Karsbøl, J.D.; Jacobsen, S.H.; Chow, F. Efficacy and safety of once-weekly semaglutide versus once-daily sitagliptin as an add-on to metformin, thiazolidinediones, or both, in patients with type 2 diabetes (SUSTAIN 2): A 56-week, double-blind, phase 3a, randomised trial. Lancet Diabetes Endocrinol. 2017, 5, 341–354. [Google Scholar] [CrossRef]
- Ahmann, A.J.; Capehorn, M.; Charpentier, G.; Dotta, F.; Henkel, E.; Lingvay, I.; Holst, A.G.; Annett, M.P.; Aroda, V.R. Efficacy and safety of once-weekly semaglutide versus exenatide ER in subjects with type 2 diabetes (SUSTAIN 3): A 56-week, open-label, randomized clinical trial. Diabetes Care 2017, 41, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Aroda, V.R.; Bain, S.C.; Cariou, B.; Piletič, M.; Rose, L.; Axelsen, M.; Rowe, E.; DeVries, J.H. Efficacy and safety of once-weekly semaglutide versus once-daily insulin glargine as add-on to metformin (with or without sulfonylureas) in insulin-naive patients with type 2 diabetes (SUSTAIN 4): A randomised, open-label, parallel-group, multicentre, multinational, phase 3a trial. Lancet Diabetes Endocrinol. 2017, 5, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Rodbard, H.W.; Lingvay, I.; Reed, J.; De La Rosa, R.; Rose, L.; Sugimoto, D.; Araki, E.; Chu, P.-L.; Wijayasinghe, N.; Norwood, P. Semaglutide added to basal insulin in type 2 diabetes (SUSTAIN 5): A randomized, controlled trial. J. Clin. Endocrinol. Metab. 2018, 103, 2291–2301. [Google Scholar] [CrossRef] [Green Version]
- Marso, S.P.; Bain, S.C.; Consoli, A.; Eliaschewitz, F.G.; Jódar, E.; Leiter, L.A.; Lingvay, I.; Rosenstock, J.; Seufert, J.; Warren, M.L.; et al. Semaglutide and cardiovascular outcomes in patients with type 2 diabetes. N. Engl. J. Med. 2016, 375, 1834–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratley, R.E.; Aroda, V.R.; Lingvay, I.; Lüdemann, J.; Andreassen, C.; Navarria, A.; Viljoen, A. Semaglutide versus dulaglutide once weekly in patients with type 2 diabetes (SUSTAIN 7): A randomised, open-label, phase 3b trial. Lancet Diabetes Endocrinol. 2018, 6, 275–286. [Google Scholar] [CrossRef]
- Lingvay, I.; Catarig, A.-M.; Frias, J.P.; Kumar, H.; Lausvig, N.L.; le Roux, C.W.; Thielke, D.; Viljoen, A.; McCrimmon, R. Efficacy and safety of once-weekly semaglutide versus daily canagliflozin as add-on to metformin in patients with type 2 diabetes (SUSTAIN 8): A double-blind, phase 3b, randomised controlled trial. Lancet Diabetes Endocrinol. 2019, 7, 834–844. [Google Scholar] [CrossRef] [Green Version]
- Capehorn, M.; Catarig, A.-M.; Furberg, J.; Janez, A.; Price, H.; Tadayon, S.; Vergès, B.; Marre, M. Efficacy and safety of once-weekly semaglutide 1.0 mg vs once-daily liraglutide 1.2 mg as add-on to 1–3 oral antidiabetic drugs in subjects with type 2 diabetes (SUSTAIN 10). Diabetes Metab. 2020, 46, 100–109. [Google Scholar] [CrossRef]
- Flint, A.; Andersen, G.; Hockings, P.; Johansson, L.; Morsing, A.; Palle, M.S.; Vogl, T.J.; Plum-Moerschel, L. Semaglutide treatment in subjects with NAFLD: Effects assessed by magnetic resonance elastography and magnetic resonance imaging proton density fat fraction. Hepatology 2020, 72, 131–1159. [Google Scholar] [CrossRef]
- Hartman, M.L.; Sanyal, A.J.; Loomba, R.; Wilson, J.M.; Nikooienejad, A.; Bray, R.; Karanikas, C.A.; Duffin, K.L.; Robins, D.A.; Haupt, A. Effects of novel dual GIP and GLP-1 receptor agonist tirzepatide on biomarkers of nonalcoholic steatohepatitis in patients with type 2 diabetes. Diabetes Care 2020, 43, 1352–1355. [Google Scholar] [CrossRef]
- Nahra, R.; Gadde, K.; Hirshberg, B.; Jermutus, L.; Maaske, J.; Stumvoll, M.; Wang, T.; Ambery, P. Effects of cotadutide on biomarkers of non-alcoholic steatohepatitis in overweight or obese subjects with type 2 diabetes mellitus: A 54-week analysis of a randomized phase 2b study. J. Hepatol. 2020, 73, S54–S55. [Google Scholar] [CrossRef]
- Vivian, E.M. Sodium-glucose co-transporter 2 (SGLT2) inhibitors: A growing class of antidiabetic agents. Drugs Context 2014, 3, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Perakakis, N.; Chrysafi, P.; Feigh, M.; Veidal, S.; Mantzoros, C. Empagliflozin improves metabolic and hepatic outcomes in a non-diabetic obese biopsy-proven mouse model of advanced NASH. Int. J. Mol. Sci. 2021, 22, 6332. [Google Scholar] [CrossRef] [PubMed]
- Shiba, K.; Tsuchiya, K.; Komiya, C.; Miyachi, Y.; Mori, K.; Shimazu, N.; Yamaguchi, S.; Ogasawara, N.; Katoh, M.; Itoh, M.; et al. Canagliflozin, an SGLT2 inhibitor, attenuates the development of hepatocellular carcinoma in a mouse model of human NASH. Sci. Rep. 2018, 8, 2362. [Google Scholar] [CrossRef] [Green Version]
- Goto, R.; Kamimura, K.; Shinagawa-Kobayashi, Y.; Sakai, N.; Nagoya, T.; Niwa, Y.; Ko, M.; Ogawa, K.; Inoue, R.; Yokoo, T.; et al. Inhibition of sodium glucose cotransporter 2 (SGLT 2) delays liver fibrosis in a medaka model of nonalcoholic steatohepatitis (NASH). FEBS Open Bio 2019, 9, 643–652. [Google Scholar] [CrossRef]
- Honda, Y.; Imajo, K.; Kato, T.; Kessoku, T.; Ogawa, Y.; Tomeno, W.; Kato, S.; Mawatari, H.; Fujita, K.; Yoneda, M.; et al. The selective SGLT2 inhibitor ipragliflozin has a therapeutic effect on nonalcoholic steatohepatitis in mice. PLoS ONE 2016, 11, e0146337. [Google Scholar] [CrossRef] [Green Version]
- Jojima, T.; Tomotsune, T.; Iijima, T.; Akimoto, K.; Suzuki, K.; Aso, Y. Empagliflozin (an SGLT2 inhibitor), alone or in combination with linagliptin (a DPP-4 inhibitor), prevents steatohepatitis in a novel mouse model of non-alcoholic steatohepatitis and diabetes. Diabetol. Metab. Syndr. 2016, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Kuchay, M.S.; Krishan, S.; Mishra, S.K.; Farooqui, K.J.; Singh, M.K.; Wasir, J.S.; Bansal, B.; Kaur, P.; Jevalikar, G.; Gill, H.K.; et al. Effect of empagliflozin on liver fat in patients with type 2 diabetes and nonalcoholic fatty liver disease: A randomized controlled trial (E-LIFT Trial). Diabetes Care 2018, 41, 1801–1808. [Google Scholar] [CrossRef] [Green Version]
- Sattar, N.; Fitchett, D.; Hantel, S.; George, J.T.; Zinman, B. Empagliflozin is associated with improvements in liver enzymes potentially consistent with reductions in liver fat: Results from randomised trials including the EMPA-REG OUTCOME® trial. Diabetologia 2018, 61, 2155–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, H.; Brown, R.; Bhullar, L.; Sohi, N.; Kalra, S.; Aronson, R. SGLT2 inhibitors and incretin agents: Associations with alanine aminotransferase activity in type 2 diabetes. Diabetes Metab. 2018, 44, 493–499. [Google Scholar] [CrossRef]
- Tobita, H.; Sato, S.; Miyake, T.; Ishihara, S.; Kinoshita, Y. Effects of dapagliflozin on body composition and liver tests in patients with nonalcoholic steatohepatitis associated with type 2 diabetes mellitus: A prospective, open-label, uncontrolled study. Curr. Ther. Res. 2017, 87, 13–19. [Google Scholar] [CrossRef]
- Seko, Y.; Sumida, Y.; Sasaki, K.; Itoh, Y.; Iijima, H.; Hashimoto, T.; Ishii, S.; Inagaki, N. Effects of canagliflozin, an SGLT2 inhibitor, on hepatic function in Japanese patients with type 2 diabetes mellitus: Pooled and subgroup analyses of clinical trials. J. Gastroenterol. 2018, 53, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Ohki, T.; Isogawa, A.; Toda, N.; Tagawa, K. Effectiveness of Ipragliflozin, a sodium-glucose co-transporter 2 inhibitor, as a second-line treatment for non-alcoholic fatty liver disease patients with type 2 diabetes mellitus who do not respond to incretin-based therapies including glucagon-like peptide-1 analogs and dipeptidyl peptidase-4 inhibitors. Clin. Drug Investig. 2016, 36, 313–319. [Google Scholar] [CrossRef]
- Ito, D.; Shimizu, S.; Inoue, K.; Saito, D.; Yanagisawa, M.; Inukai, K.; Akiyama, Y.; Morimoto, Y.; Noda, M.; Shimada, A. Comparison of Ipragliflozin and Pioglitazone effects on nonalcoholic fatty liver disease in patients with type 2 diabetes: A randomized, 24-week, open-label, active-controlled trial. Diabetes Care 2017, 40, 1364–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bando, Y.; Ogawa, A.; Ishikura, K.; Kanehara, H.; Hisada, A.; Notumata, K.; Okafuji, K.; Toya, D. The effects of ipragliflozin on the liver-to-spleen attenuation ratio as assessed by computed tomography and on alanine transaminase levels in Japanese patients with type 2 diabetes mellitus. Diabetol. Int. 2017, 8, 218–227. [Google Scholar] [CrossRef]
- Ohta, A.; Kato, H.; Ishii, S.; Sasaki, Y.; Nakamura, Y.; Nakagawa, T.; Nagai, Y.; Tanaka, Y. Ipragliflozin, a sodium glucose co-transporter 2 inhibitor, reduces intrahepatic lipid content and abdominal visceral fat volume in patients with type 2 diabetes. Expert Opin. Pharmacother. 2017, 18, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.-L.; Vethakkan, S.R.; Mustapha, N.R.N.; Mahadeva, S.; Chan, W.-K. Empagliflozin for the treatment of nonalcoholic Steatohepatitis in patients with type 2 diabetes mellitus. Dig. Dis. Sci. 2020, 65, 623–631. [Google Scholar] [CrossRef]
- De Boer, R.A.; Núñez, J.; Kozlovski, P.; Wang, Y.; Proot, P.; Keefe, D. Effects of the dual sodium–glucose linked transporter inhibitor, licogliflozin vs placebo or empagliflozin in patients with type 2 diabetes and heart failure. Br. J. Clin. Pharmacol. 2020, 86, 1346–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diehl, A.; Harrison, S.; Caldwell, S.; Rinella, M.; Paredes, A.; Moylan, C.; Guy, C.; Bashir, M.; Wang, Y.; Miller, L.; et al. JKB-121 in patients with nonalcoholic steatohepatitis: A phase 2 double blind randomized placebo control study. J. Hepatol. 2018, 68, S103. [Google Scholar] [CrossRef]
- Krenkel, O.; Puengel, T.; Govaere, O.; Abdallah, A.T.; Mossanen, J.C.; Kohlhepp, M.; Liepelt, A.; Lefebvre, E.; Luedde, T.; Hellerbrand, C.; et al. Therapeutic inhibition of inflammatory monocyte recruitment reduces steatohepatitis and liver fibrosis. Hepatology 2018, 67, 1270–1283. [Google Scholar] [CrossRef] [Green Version]
- Friedman, S.L.; Ratziu, V.; Harrison, S.A.; Abdelmalek, M.; Aithal, G.; Caballeria, J.; Francque, S.; Farrell, G.; Kowdley, K.V.; Craxi, A.; et al. A randomized, placebo-controlled trial of cenicriviroc for treatment of nonalcoholic steatohepatitis with fibrosis. Hepatology 2018, 67, 1754–1767. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Abenavoli, L.; Mykhalchyshyn, G.; Kononenko, L.; Boccuto, L.; Kyriienko, D.; Dynnyk, O. A multi-strain probiotic reduces the fatty liver index, cytokines and aminotransferase levels in NAFLD patients: Evidence from a randomized clinical trial. J. Gastrointest. Liver Dis. 2018, 27, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Behrouz, V.; Jazayeri, S.; Aryaeian, N.; Zahedi, M.J.; Hosseini, F. Effects of probiotic and prebiotic supplementation on leptin, adiponectin, and glycemic parameters in non-alcoholic fatty liver disease: A randomized clinical trial. Middle East J. Dig. Dis. 2017, 9, 150–157. [Google Scholar] [CrossRef] [Green Version]
- Eslamparast, T.; Poustchi, H.; Zamani, F.; Sharafkhah, M.; Malekzadeh, R.; Hekmatdoost, A. Synbiotic supplementation in nonalcoholic fatty liver disease: A randomized, double-blind, placebo-controlled pilot study. Am. J. Clin. Nutr. 2014, 99, 535–542. [Google Scholar] [CrossRef]
- Shidfar, F.; Ekhlasi, G.; Mohammadi, R.K.; Agah, S.; Zarrati, M.; Hosseini, A.F.; Arabshahi, S.S.S. Do symbiotic and vitamin E supplementation have favorite effects in nonalcoholic fatty liver disease? A randomized, double-blind, placebo-controlled trial. J. Res. Med. Sci. 2016, 21, 106. [Google Scholar] [CrossRef] [PubMed]
- Bakhshimoghaddam, F.; Shateri, K.; Sina, M.; Hashemian, M.; Alizadeh, M. Daily consumption of synbiotic yogurt decreases liver steatosis in patients with nonalcoholic fatty liver disease: A randomized controlled clinical trial. J. Nutr. 2018, 148, 1276–1284. [Google Scholar] [CrossRef] [Green Version]
- Scorletti, E.; Afolabi, P.R.; Miles, E.A.; Smith, D.E.; Almehmadi, A.; AlShathry, A.; Childs, C.E.; Del Fabbro, S.; Bilson, J.; Moyses, H.E.; et al. Synbiotics alter fecal microbiomes, but not liver fat or fibrosis, in a randomized trial of patients with nonalcoholic fatty liver disease. Gastroenterology 2020, 158, 1597–1610.e7. [Google Scholar] [CrossRef]
- Sayari, S.; Neishaboori, H.; Jameshorani, M. Combined effects of synbiotic and sitagliptin versus sitagliptin alone in patients with nonalcoholic fatty liver disease. Clin. Mol. Hepatol. 2018, 24, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askari, G.; Asgharian, A.; Esmailzade, A.; Feizi, A.; Mohammadi, V. The effect of symbiotic supplementation on liver enzymes, c-reactive protein and ultrasound findings in patients with non-alcoholic fatty liver disease: A clinical trial. Int. J. Prev. Med. 2016, 7, 59. [Google Scholar] [CrossRef]
- Javadi, L.; Ghavami, M.; Khoshbaten, M.; Safaiyan, A.; Barzegari, A.; Gargari, B.P. The effect of probiotic and/or prebiotic on liver function tests in patients with nonalcoholic fatty liver disease: A double blind randomized clinical trial. Iran. Red Crescent Med. J. 2017, 19, e46017. [Google Scholar] [CrossRef]
- Kristensen, N.B.; Bryrup, T.; Allin, K.; Nielsen, T.; Hansen, T.H.; Pedersen, O. Alterations in fecal microbiota composition by probiotic supplementation in healthy adults: A systematic review of randomized controlled trials. Genome Med. 2016, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Craven, L.; Rahman, A.; Nair Parvathy, S.; Beaton, M.; Silverman, J.; Qumosani, K.; Hramiak, I.; Hegele, R.; Joy, T.; Meddings, J.; et al. Allogenic fecal microbiota transplantation in patients with nonalcoholic fatty liver disease improves abnormal small intestinal permeability: A randomized control trial. Am. J. Gastroenterol. 2020, 115, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Kootte, R.S.; Levin, E.; Salojärvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of insulin sensitivity after lean donor feces in metabolic syndrome is driven by baseline intestinal microbiota composition. Cell Metab. 2017, 26, 611–619.e6. [Google Scholar] [CrossRef] [Green Version]
- Witjes, J.J.; Smits, L.P.; Pekmez, C.T.; Prodan, A.; Meijnikman, A.S.; Troelstra, M.A.; Bouter, K.E.; Herrema, H.; Levin, E.; Holleboom, A.G.; et al. Donor fecal microbiota transplantation alters gut microbiota and metabolites in obese individuals with steatohepatitis. Hepatol. Commun. 2020, 4, 1578–1590. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef]
- Li, S.S.; Zhu, A.; Benes, V.; Costea, P.I.; Hercog, R.; Hildebrand, F.; Huerta-Cepas, J.; Nieuwdorp, M.; Salojärvi, J.; Voigt, A.Y.; et al. Durable coexistence of donor and recipient strains after fecal microbiota transplantation. Science 2016, 352, 586–589. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.; Scheithauer, T.; Bakker, G.J.; Prodan, A.; Levin, E.; Khan, M.T.; Herrema, H.; Ackermans, M.; Serlie, M.J.M.; De Brauw, M.; et al. Donor metabolic characteristics drive effects of faecal microbiota transplantation on recipient insulin sensitivity, energy expenditure and intestinal transit time. Gut 2020, 69, 502–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allegretti, J.R.; Kassam, Z.; Mullish, B.H.; Chiang, A.; Carrellas, M.; Hurtado, J.; Marchesi, J.R.; McDonald, J.A.K.; Pechlivanis, A.; Barker, G.F.; et al. Effects of fecal microbiota transplantation with oral capsules in obese patients. Clin. Gastroenterol. Hepatol. 2020, 18, 855–863. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Y.; Yang, Y.; Zhang, N.; Niu, C.; Shen, X.; Wu, J. Novel approaches to intervene gut microbiota in the treatment of chronic liver diseases. FASEB J. 2021, 35, e21871. [Google Scholar] [CrossRef]
- Suh, G.A. Phage therapy for alcohol-associated hepatitis. Hepatology 2020, 73, 1609–1610. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Llorente, C.; Lang, S.; Brandl, K.; Chu, H.; Jiang, L.; White, R.C.; Clarke, T.H.; Nguyen, K.; Torralba, M.; et al. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature 2019, 575, 505–511. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, C.; Cui, J.; Lu, J.; Yan, C.; Wei, X.; Zhao, X.; Li, N.; Li, S.; Xue, G.; et al. Fatty liver disease caused by high-alcohol-producing klebsiella pneumoniae. Cell Metab. 2019, 30, 1172. [Google Scholar] [CrossRef]
- Mikolasevic, I.; Delija, B.; Mijic, A.; Stevanovic, T.; Skenderevic, N.; Sosa, I.; Krznaric-Zrnic, I.; Abram, M.; Krznaric, Z.; Domislovic, V.; et al. Small intestinal bacterial overgrowth and non-alcoholic fatty liver disease diagnosed by transient elastography and liver biopsy. Int. J. Clin. Pract. 2021, 75, e13947. [Google Scholar] [CrossRef]
- Haas, J.T.; Francque, S.M.; Staels, B. Pathophysiology and mechanisms of nonalcoholic fatty liver disease. Annu. Rev. Physiol. 2016, 78, 18.1–18.25. [Google Scholar] [CrossRef]
- Hallsworth, K.; Adams, L.A. Lifestyle modification in NAFLD/NASH: Facts and figures. JHEP Rep. 2019, 1, 468–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younossi, Z.M. Nonalcoholic fatty liver disease and nonalcoholic steatohepatitis: Implications for liver transplantation. Liver Transplant. 2017, 24, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Caussy, C.; Reeder, S.B.; Sirlin, C.B.; Loomba, R. Noninvasive, quantitative assessment of liver fat by MRI-PDFF as an endpoint in NASH Trials. Hepatology 2018, 68, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Young, S.; Tariq, R.; Provenza, J.; Satapathy, S.K.; Faisal, K.; Choudhry, A.; Friedman, S.L.; Singal, A.K. Prevalence and profile of nonalcoholic fatty liver disease in lean adults: Systematic review and meta-analysis. Hepatol. Commun. 2020, 4, 953–972. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, J.; Frings, V.G.; Geier, A.; Goebeler, M.; Kerstan, A. Non-alcoholic fatty liver disease and psoriasis—Is there a shared proinflammatory network? J. Dtsch. Dermatol. Ges. 2021, 19, 517–528. [Google Scholar] [CrossRef]
- Karaivazoglou, K.; Konstantakis, C.; Tourkochristou, E.; Assimakopoulos, S.F.; Triantos, C. Non-alcoholic fatty liver disease in inflammatory bowel disease patients. Eur. J. Gastroenterol. Hepatol. 2020, 32, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Majzoub, A.M.; Nayfeh, T.; Barnard, A.; Munaganuru, N.; Dave, S.; Singh, S.; Murad, M.H.; Loomba, R. Systematic review with network meta-analysis: Comparative efficacy of pharmacologic therapies for fibrosis improvement and resolution of NASH. Aliment. Pharmacol. Ther. 2021, 54, 880–889. [Google Scholar] [CrossRef]
- Ampuero, J.; Romero-Gomez, M. Stratification of patients in NASH clinical trials: A pitfall for trial success. JHEP Rep. 2020, 2, 100148. [Google Scholar] [CrossRef]
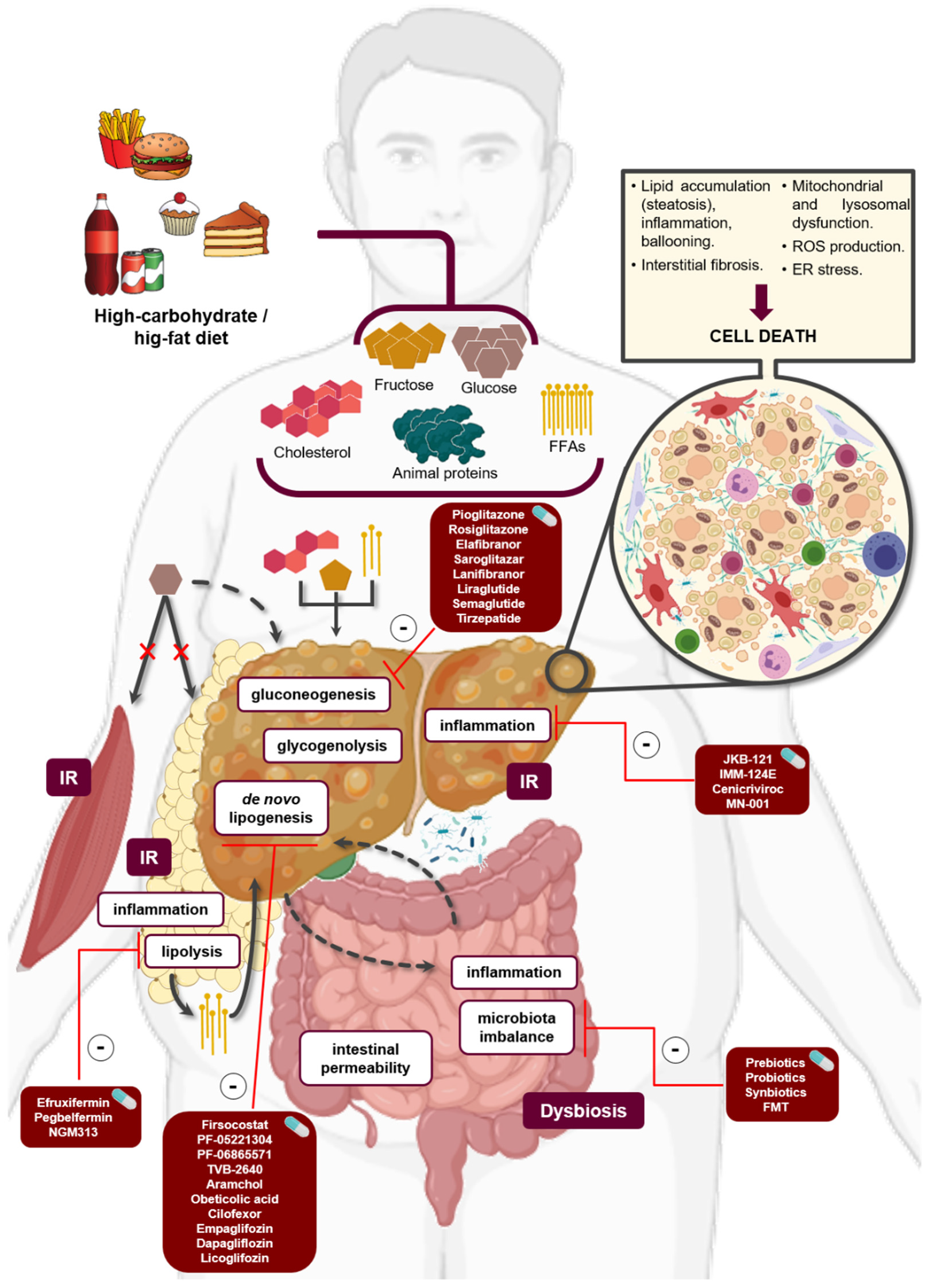
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Laso, A.; Gutiérrez-Larrañaga, M.; Alonso-Peña, M.; Medina, J.M.; Iruzubieta, P.; Arias-Loste, M.T.; López-Hoyos, M.; Crespo, J. Pathophysiological Mechanisms in Non-Alcoholic Fatty Liver Disease: From Drivers to Targets. Biomedicines 2022, 10, 46. https://doi.org/10.3390/biomedicines10010046
Santos-Laso A, Gutiérrez-Larrañaga M, Alonso-Peña M, Medina JM, Iruzubieta P, Arias-Loste MT, López-Hoyos M, Crespo J. Pathophysiological Mechanisms in Non-Alcoholic Fatty Liver Disease: From Drivers to Targets. Biomedicines. 2022; 10(1):46. https://doi.org/10.3390/biomedicines10010046
Chicago/Turabian StyleSantos-Laso, Alvaro, María Gutiérrez-Larrañaga, Marta Alonso-Peña, Juan M. Medina, Paula Iruzubieta, María Teresa Arias-Loste, Marcos López-Hoyos, and Javier Crespo. 2022. "Pathophysiological Mechanisms in Non-Alcoholic Fatty Liver Disease: From Drivers to Targets" Biomedicines 10, no. 1: 46. https://doi.org/10.3390/biomedicines10010046
APA StyleSantos-Laso, A., Gutiérrez-Larrañaga, M., Alonso-Peña, M., Medina, J. M., Iruzubieta, P., Arias-Loste, M. T., López-Hoyos, M., & Crespo, J. (2022). Pathophysiological Mechanisms in Non-Alcoholic Fatty Liver Disease: From Drivers to Targets. Biomedicines, 10(1), 46. https://doi.org/10.3390/biomedicines10010046