

Inhibitors of the RBD-ACE-2 Found among a Wide Range of Dyes by the Immunoassay Method

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemistry
2.2. Plasmid Construction, Recombinant Protein Expression, and Purification
2.3. Preparation ACE2-HRP Conjugate
2.4. ELISA-Based Competitive Inhibition of the RBD(S-trimer)/ACE2 Interaction
2.5. Statistical Analysis
3. Results and Discussion
3.1. The Testing System
3.2. Screening Assays
3.3. Binding Inhibition (Concentration–Response)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Novikov, F.N.; Stroylov, V.S.; Svitanko, I.V.; Nebolsin, V.E. Molecular basis of COVID-19 pathogenesis. Russ. Chem. Rev. 2020, 89, 858–878. [Google Scholar] [CrossRef]
- Yan, W.; Zheng, Y.; Zeng, X.; He, B.; Cheng, W. Structural biology of SARS-CoV-2: Open the door for novel therapies. Signal Transduct. Target. Ther. 2022, 7, 26. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Mishevich, J.L.; Mesecar, A.; Mitsuya, H. Recent Drug Development and Medicinal Chemistry Approaches for the Treatment of SARS-CoV-2 Infection and COVID-19. ChemMedChem 2022, 17, e202200440. [Google Scholar] [CrossRef]
- Tang, T.; Bidon, M.; Jaimes, J.A.; Whittaker, G.R.; Daniel, S. Coronavirus membrane fusion mechanism offers a potential target for antiviral development. Antivir. Res. 2020, 178, 104792. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Bojadzic, D.; Alcazar, O.; Chen, J.; Chuang, S.-T.; Condor Capcha, J.M.; Shehadeh, L.A.; Buchwald, P. Small-Molecule Inhibitors of the Coronavirus Spike: ACE2 Protein–Protein Interaction as Blockers of Viral Attachment and Entry for SARS-CoV-2. ACS Infect. Dis. 2021, 7, 1519–1534. [Google Scholar] [CrossRef]
- Limonta-Fernández, M.; Chinea-Santiago, G.; Martín-Dunn, A.M.; Gonzalez-Roche, D.; Bequet-Romero, M.; Marquez-Perera, G.; González-Moya, I.; Canaan-Haden-Ayala, C.; Cabrales-Rico, A.; Espinosa-Rodríguez, L.A.; et al. An engineered SARS-CoV-2 receptor-binding domain produced in Pichia pastoris as a candidate vaccine antigen. New Biotechnol. 2022, 72, 11–21. [Google Scholar] [CrossRef]
- Yarovaya, O.I.; Shcherbakov, D.N.; Borisevich, S.S.; Sokolova, A.S.; Gureev, M.A.; Khamitov, E.M.; Rudometova, N.B.; Zybkina, A.V.; Mordvinova, E.D.; Zaykovskaya, A.V.; et al. Borneol Ester Derivatives as Entry Inhibitors of a Wide Spectrum of SARS-CoV-2 Viruses. Viruses 2022, 14, 1295. [Google Scholar] [CrossRef]
- Filimonov, A.S.; Yarovaya, O.I.; Zaykovskaya, A.V.; Rudometova, N.B.; Shcherbakov, D.N.; Chirkova, V.Y.; Baev, D.S.; Borisevich, S.S.; Luzina, O.A.; Pyankov, O.V.; et al. (+)-Usnic Acid and Its Derivatives as Inhibitors of a Wide Spectrum of SARS-CoV-2 Viruses. Viruses 2022, 14, 2154. [Google Scholar] [CrossRef]
- Holme, I. Sir William Henry Perkin: A review of his life, work and legacy. Color. Technol. 2006, 122, 235–251. [Google Scholar] [CrossRef]
- Shi, G.; Shen, Q.; Zhang, C.; Ma, J.; Mohammed, A.; Zhao, H. Efficacy and Safety of Gabapentin in the Treatment of Chronic Cough: A Systematic Review. Tuberc. Respir. Dis. 2018, 81, 167. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.-T.; Papp, H.; Kuczmog, A.; Eells, R.; Condor Capcha, J.M.; Shehadeh, L.A.; Jakab, F.; Buchwald, P. Methylene Blue Is a Nonspecific Protein–Protein Interaction Inhibitor with Potential for Repurposing as an Antiviral for COVID-19. Pharmaceuticals 2022, 15, 621. [Google Scholar] [CrossRef] [PubMed]
- Fryk, J.J.; Marks, D.C.; Hobson-Peters, J.; Prow, N.A.; Watterson, D.; Hall, R.A.; Young, P.R.; Reichenberg, S.; Sumian, C.; Faddy, H.M. Dengue and chikungunya viruses in plasma are effectively inactivated after treatment with methylene blue and visible light. Transfusion 2016, 56, 2278–2285. [Google Scholar] [CrossRef] [PubMed]
- Eickmann, M.; Gravemann, U.; Handke, W.; Tolksdorf, F.; Reichenberg, S.; Müller, T.H.; Seltsam, A. Inactivation of Ebola virus and Middle East respiratory syndrome coronavirus in platelet concentrates and plasma by ultraviolet C light and methylene blue plus visible light, respectively. Transfusion 2018, 58, 2202–2207. [Google Scholar] [CrossRef]
- Faddy, H.M.; Fryk, J.J.; Hall, R.A.; Young, P.R.; Reichenberg, S.; Tolksdorf, F.; Sumian, C.; Gravemann, U.; Seltsam, A.; Marks, D.C. Inactivation of yellow fever virus in plasma after treatment with methylene blue and visible light and in platelet concentrates following treatment with ultraviolet C light. Transfusion 2019, 59, 2223–2227. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, K.; Liao, X.; Luo, G.; Kumthip, K.; Leetrakool, N.; Li, S.; Chen, L.; Yang, C.; Chen, Y. Inactivation of Zika virus in plasma and derivatives by four different methods. J. Med. Virol. 2019, 91, 2059–2065. [Google Scholar] [CrossRef]
- Ghahestani, S.M.; Shahab, E.; Karimi, S.; Madani, M.H. Methylene blue may have a role in the treatment of COVID-19. Med. Hypotheses 2020, 144, 110163. [Google Scholar] [CrossRef]
- Gendrot, M.; Andreani, J.; Duflot, I.; Boxberger, M.; Le Bideau, M.; Mosnier, J.; Jardot, P.; Fonta, I.; Rolland, C.; Bogreau, H.; et al. Methylene blue inhibits replication of SARS-CoV-2 in vitro. Int. J. Antimicrob. Agents 2020, 56, 106202. [Google Scholar] [CrossRef]
- Scigliano, G.; Scigliano, G.A. Methylene blue in covid-19. Med. Hypotheses 2021, 146, 110455. [Google Scholar] [CrossRef]
- Bojadzic, D.; Alcazar, O.; Buchwald, P. Methylene Blue Inhibits the SARS-CoV-2 Spike–ACE2 Protein-Protein Interaction–a Mechanism that can Contribute to its Antiviral Activity against COVID-19. Front. Pharmacol. 2021, 11, 2255. [Google Scholar] [CrossRef]
- Li, Z.; Xu, J.; Lang, Y.; Wu, X.; Hu, S.; Samrat, S.K.; Tharappel, A.M.; Kuo, L.; Butler, D.; Song, Y.; et al. In vitro and in vivo characterization of erythrosin B and derivatives against Zika virus. Acta Pharm. Sin. B 2022, 12, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Merkuleva, I.A.; Shcherbakov, D.N.; Borgoyakova, M.B.; Shanshin, D.V.; Rudometov, A.P.; Karpenko, L.I.; Belenkaya, S.V.; Isaeva, A.A.; Nesmeyanova, V.S.; Kazachinskaia, E.I.; et al. Comparative Immunogenicity of the Recombinant Receptor-Binding Domain of Protein S SARS-CoV-2 Obtained in Prokaryotic and Mammalian Expression Systems. Vaccines 2022, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Merkuleva, I.A.; Shcherbakov, D.N.; Borgoyakova, M.B.; Isaeva, A.A.; Nesmeyanova, V.S.; Volkova, N.V.; Aripov, V.S.; Shanshin, D.V.; Karpenko, L.I.; Belenkaya, S.V.; et al. Are Hamsters a Suitable Model for Evaluating the Immunogenicity of RBD-Based Anti-COVID-19 Subunit Vaccines? Viruses 2022, 14, 1060. [Google Scholar] [CrossRef] [PubMed]
- Yaneva, Z.; Ivanova, D.; Nikolova, N.; Toneva, M. Organic dyes in contemporary medicinal chemistry and biomedicine. I. From the chromophore to the bioimaging/bioassay agent. Biotechnol. Biotechnol. Equip. 2022, 36, 1–14. [Google Scholar] [CrossRef]
- Saeidnia, S.; Manayi, A. Phenolphthalein. In Encyclopedia of Toxicology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 3, pp. 877–880. ISBN 9780123864543. [Google Scholar]
- Schoof, M.; Faust, B.; Saunders, R.A.; Sangwan, S.; Rezelj, V.; Hoppe, N.; Boone, M.; Billesbølle, C.B.; Puchades, C.; Azumaya, C.M.; et al. An ultrapotent synthetic nanobody neutralizes SARS-CoV-2 by stabilizing inactive Spike. Science 2020, 370, 1473–1479. [Google Scholar] [CrossRef]
- Svyatchenko, V.A.; Nikonov, S.D.; Mayorov, A.P.; Gelfond, M.L.; Loktev, V.B. Antiviral photodynamic therapy: Inactivation and inhibition of SARS-CoV-2 in vitro using methylene blue and Radachlorin. Photodiagnosis Photodyn. Ther. 2021, 33, 102112. [Google Scholar] [CrossRef]
- Lobo, C.S.; Rodrigues-Santos, P.; Pereira, D.; Núñez, J.; Trêpa, J.C.D.; Sousa, D.L.; Lourenço, J.V.; Coelho, M.F.; de Almeida, L.P.; da Cunha, J.S.; et al. Photodynamic disinfection of SARS-CoV-2 clinical samples using a methylene blue formulation. Photochem. Photobiol. Sci. 2022, 21, 1101–1109. [Google Scholar] [CrossRef]
- Chen, P.; Liang, Y.; Xu, Y.; Zhao, Y.; Song, S. Synchronous photosensitized degradation of methyl orange and methylene blue in water by visible-light irradiation. J. Mol. Liq. 2021, 334, 116159. [Google Scholar] [CrossRef]
- Chiou, W.-C.; Chen, J.-C.; Chen, Y.-T.; Yang, J.-M.; Hwang, L.-H.; Lyu, Y.-S.; Yang, H.-Y.; Huang, C. The inhibitory effects of PGG and EGCG against the SARS-CoV-2 3C-like protease. Biochem. Biophys. Res. Commun. 2022, 591, 130–136. [Google Scholar] [CrossRef]
- Tan, H.; Ma, C.; Wang, J. Invalidation of dieckol and 1,2,3,4,6-pentagalloylglucose (PGG) as SARS-CoV-2 main protease inhibitors and the discovery of PGG as a papain-like protease inhibitor. Med. Chem. Res. 2022, 31, 1147–1153. [Google Scholar] [CrossRef]
- Ganesan, L.; Margolles-Clark, E.; Song, Y.; Buchwald, P. The food colorant erythrosine is a promiscuous protein-protein interaction inhibitor. Biochem. Pharmacol. 2011, 81, 810–818. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Compound | Structure of the Compound | Name | Color Index Name (Number) | ||||||
|---|---|---|---|---|---|---|---|---|---|
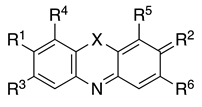 | |||||||||
| X | R1 | R2 | R3 | R4 | R5 | R6 | |||
| 1a | S | N(CH3)2 | N+(CH3)2Cl− | H | H | H | H | Methylene blue (chloride) | Basic Blue 9 (52015) |
| 1b | S | N(CH3)2 | N+(CH3)2Cl− | H | H | H | H | Methylene blue (double zinc) | Basic Blue 9 (52015) |
| 2 | S | NH2 | N+(CH3)2Cl− | CH3 | H | H | H | Toluidine blue | Basic Blue 17 (52040) |
| 3 | S | N(CH3)2 | N+(CH3)2Cl− | H | H | NO2 | H | Methylene green | Basic Green 5 (52020) |
| 4 | O | N(CH3)2 | O | H | H | OH | COOH | Gallocyanine | Mordant Blue 10 (51030) |
| 5 | O | 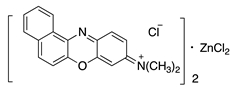 | Meldola’s Blue | Basic Blue 6 (51175) | |||||
| 6 | N | 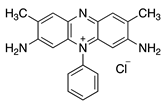 | Safranine | Basic Red 2 (50240) | |||||
| Compound | Structure | Name | Color Index Name (Number) | |||||
|---|---|---|---|---|---|---|---|---|
 | ||||||||
| X | R1 | R2 | R3 | R4 | R5 | |||
| 7 | N | H | NH2 | NH2+Cl− | CH3 | H | Acridine Yellow | Basic Yellow K (46025) |
| 8 | O |  | OH | O | H | H | Fluorescein | Acid Yellow 73 (45350) |
| 9 | O | 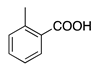 | OH | O | H | Br | 4′,5′-dibromofluorescein | Acid Orange 11 (45370) |
| 10a | O | 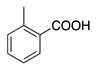 | OH | O | Br | Br | Eosin (free acid) | Acid Red 87 (45380) |
| 10b | O |  | ONa | O | Br | Br | Eosin (Na salt) | Acid Red 87 (45380) |
| 11 | O | 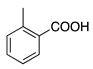 | OH | O | I | I | Erythrosine B | Solvent Red 140 (45430) |
| 12 | O | 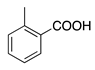 | N(C2H5)2 | N(C2H5)2+Cl− | H | H | Rhodamine B | Basic Violet 10 (45170) |
| 13 | O | 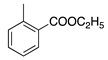 | N(C2H5)2 | N(C2H5)2+Cl− | CH3 | H | Rhodamine 6G | Basic Red 1 (45160) |
| Compound | Structure | Name | Color Index Name (Number) | ||
|---|---|---|---|---|---|
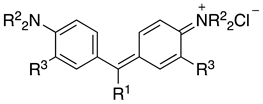 | |||||
| R1 | R2 | R3 | |||
| 14 | NH2 | CH3 | H | Auramine O | Basic Yellow 2 (41000) |
| 15 |  | H | H | Fuchsin | Basic Red 9 (42500) |
| 16 |  | H | CH3 | New Fuchsin | Basic Violet 2 (42520) |
| 17 | 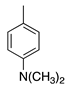 | CH3 | H | Crystal Violet | Basic Violet 3 (42555) |
| 18 |  | CH3 | H | Methyl Green | Basic Blue 20 (42585) |
| 19 | C6H5 | CH3 | H | Malachite Green | Basic Green 4 (42000) |
| 20 | 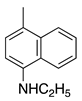 | C2H5 | H | Victoria Pure Blue BO | Basic Blue 7 (42595) |
| 21 | 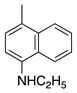 | CH3 | H | Basic Blue 3 | Basic Blue 11 (44040) |
| 22 | 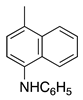 | CH3 | H | Victoria Blue B | Basic Blue 26 (44045) |
| 23 |  | CH3 | H | Permanent Purple | Basic Violet 1 (42535:2) |
| 24 | 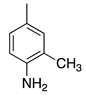 | H | H | Basic Fuchsin | Basic Violet 14 (42510) |
| 25 | 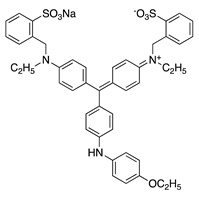 | Brilliant Blue R | Acid Blue 83 (42660) | ||
| 26 | 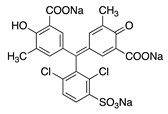 | Chrome Azurol S | Mordant Blue 29 (43825) | ||
| 27 | 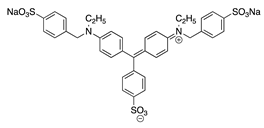 | Light Green SF | Acid Green 5 (42095) | ||
| 28 | 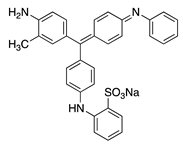 | Alkali Blue 4B | Acid Blue 110 (42750) | ||
| Compound | Structure | Name | |||
|---|---|---|---|---|---|
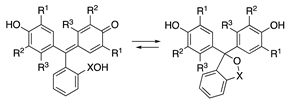 | |||||
| X | R1 | R2 | R3 | ||
| 29 | CO | H | CH3 | H | o-Cresolphthalein |
| 30 | CO | CH(CH3)2 | H | CH3 | Thymolphthalein |
| 31 | CO | CH(CH3)2 | CHN(CH2COOH)2 | CH3 | Thymolphthalexone |
| 32 | SO2 | Br | Br | H | Bromophenol Blue |
| 33 | SO2 | CH(CH3)2 | H | CH3 | Thymol Blue |
| 34 | SO2 | CH3 | CHN(CH2COONa)2 | H | Xylenol Orange |
| Compound | IC50, μM RBD-Wuhan | IC50, μM S Trimer-Wuhan | IC50, μM RBD-Delta | IC50, μM S Trimer-Delta |
|---|---|---|---|---|
| 8 | 791 ± 9 | 328 ± 15 | 484 ± 21 | 168 ± 21 |
| 10a | 91 ± 12 | 34 ± 3 | 66 ± 5 | 12 ± 5 |
| 10b | 67 ± 5 | 24 ± 2 | 47 ± 6 | 13 ± 3 |
| 11 | 42 ± 1 | 8.5 ± 0.5 | 43 ± 3 | 3.4 ± 0.8 |
| 27 | 156 ± 13 | 58 ± 9 | 245 ± 8 | 43 ± 7 |
| 29 | 239 ± 14 | 138 ± 20 | NT * | 23 ± 7 |
| 30 | >350 | >350 | NT | 212 ± 24 |
| 31 | >350 | >350 | NT | 113 ± 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mordvinova, E.D.; Nikitina, P.A.; Yarovaya, O.I.; Volosnikova, E.A.; Murashkin, D.E.; Isaeva, A.A.; Koldaeva, T.Y.; Perevalov, V.P.; Salakhutdinov, N.F.; Shcherbakov, D.N. Inhibitors of the RBD-ACE-2 Found among a Wide Range of Dyes by the Immunoassay Method. Chemosensors 2023, 11, 135. https://doi.org/10.3390/chemosensors11020135
Mordvinova ED, Nikitina PA, Yarovaya OI, Volosnikova EA, Murashkin DE, Isaeva AA, Koldaeva TY, Perevalov VP, Salakhutdinov NF, Shcherbakov DN. Inhibitors of the RBD-ACE-2 Found among a Wide Range of Dyes by the Immunoassay Method. Chemosensors. 2023; 11(2):135. https://doi.org/10.3390/chemosensors11020135
Chicago/Turabian StyleMordvinova, Ekaterina D., Polina A. Nikitina, Olga I. Yarovaya, Ekaterina A. Volosnikova, Denis E. Murashkin, Anastasiya A. Isaeva, Tatiana Y. Koldaeva, Valery P. Perevalov, Nariman F. Salakhutdinov, and Dmitriy N. Shcherbakov. 2023. "Inhibitors of the RBD-ACE-2 Found among a Wide Range of Dyes by the Immunoassay Method" Chemosensors 11, no. 2: 135. https://doi.org/10.3390/chemosensors11020135
APA StyleMordvinova, E. D., Nikitina, P. A., Yarovaya, O. I., Volosnikova, E. A., Murashkin, D. E., Isaeva, A. A., Koldaeva, T. Y., Perevalov, V. P., Salakhutdinov, N. F., & Shcherbakov, D. N. (2023). Inhibitors of the RBD-ACE-2 Found among a Wide Range of Dyes by the Immunoassay Method. Chemosensors, 11(2), 135. https://doi.org/10.3390/chemosensors11020135