
Recent Advances in Molecular Fluorescent Probes for CYP450 Sensing and Imaging




Abstract
:1. Introduction
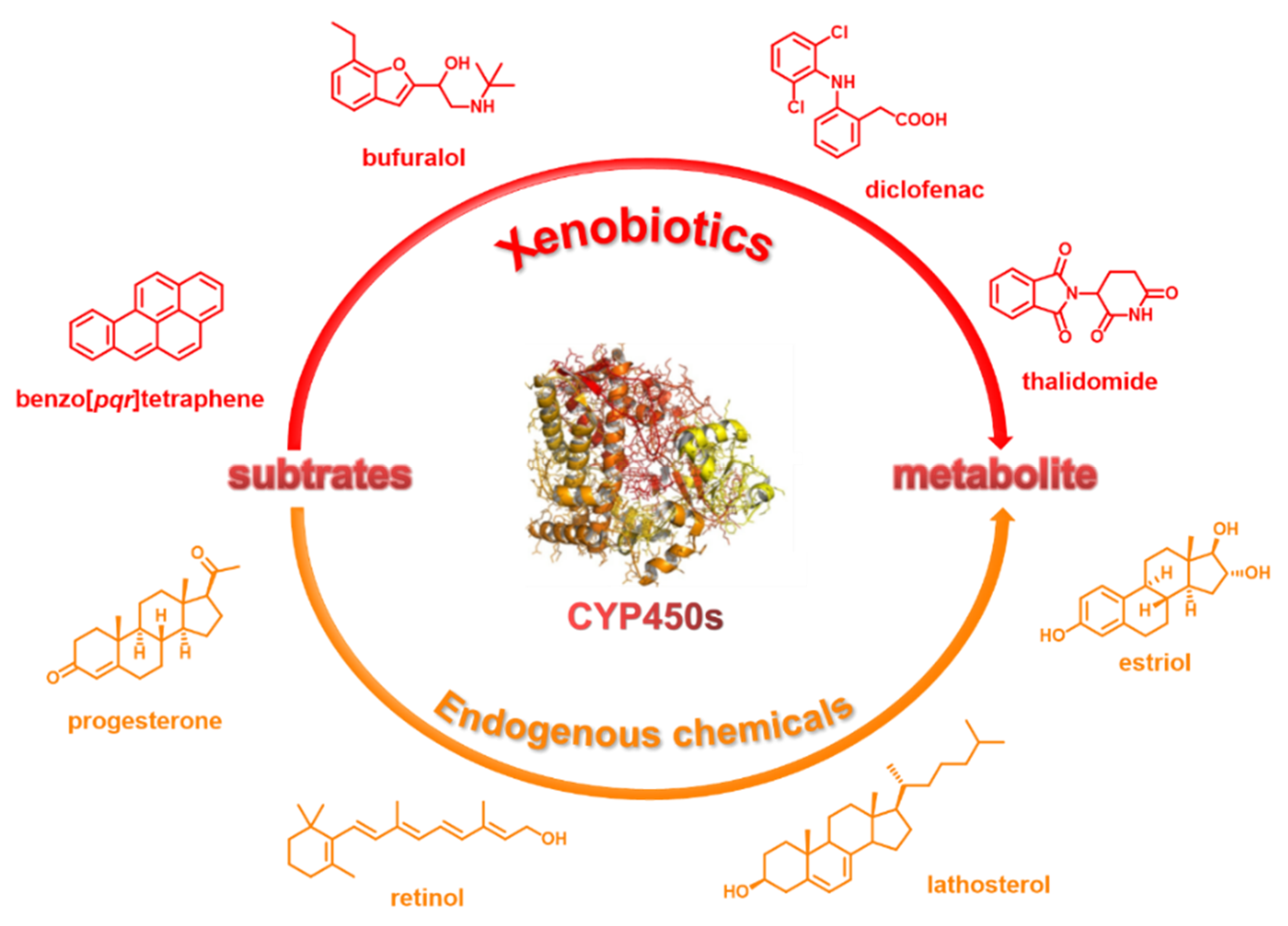
2. Brief Introduction of CYP450
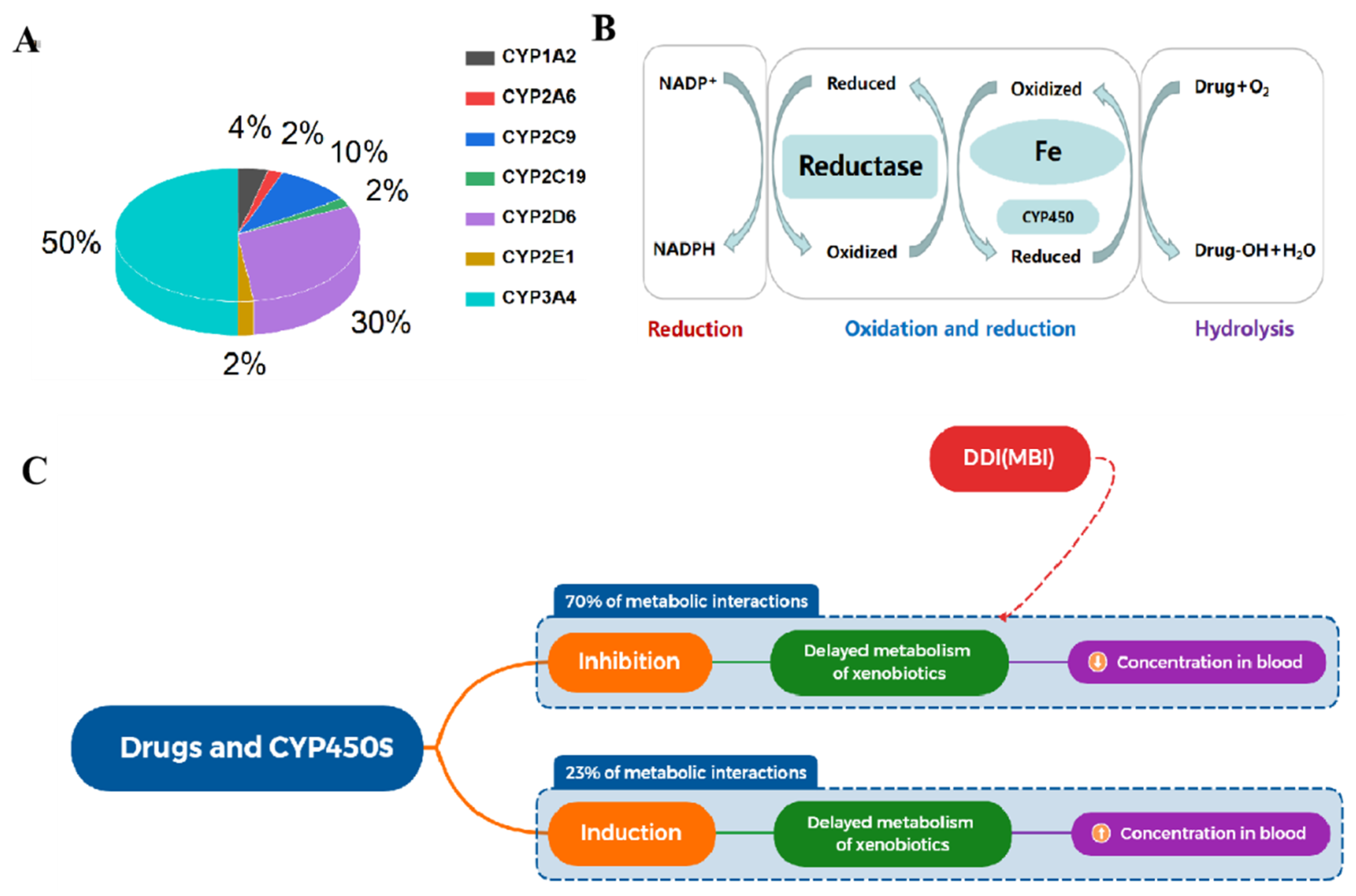
2.1. Relevance to Diseases and Drug Interactions
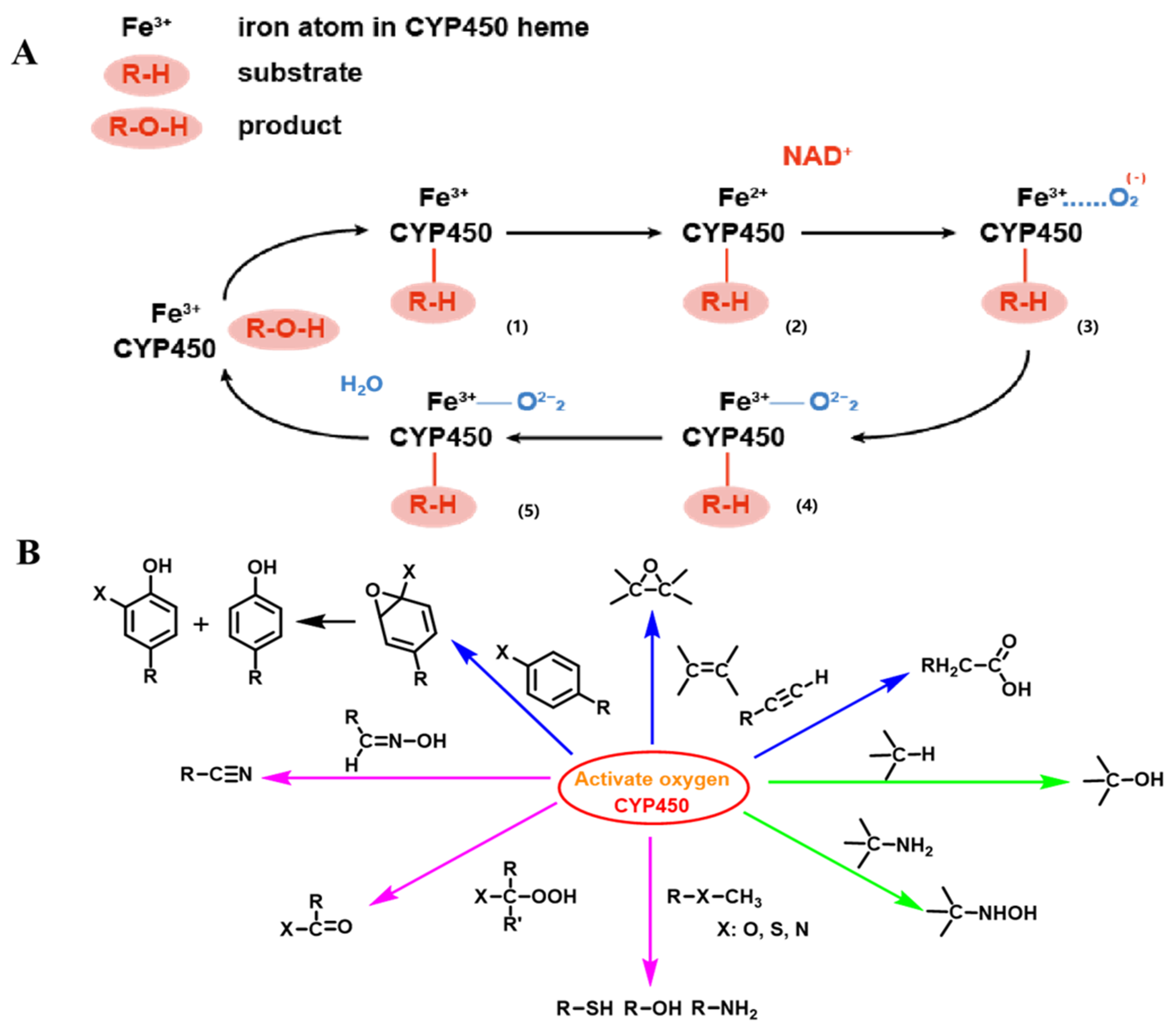
2.2. CYP450-Catalyzed Reactions
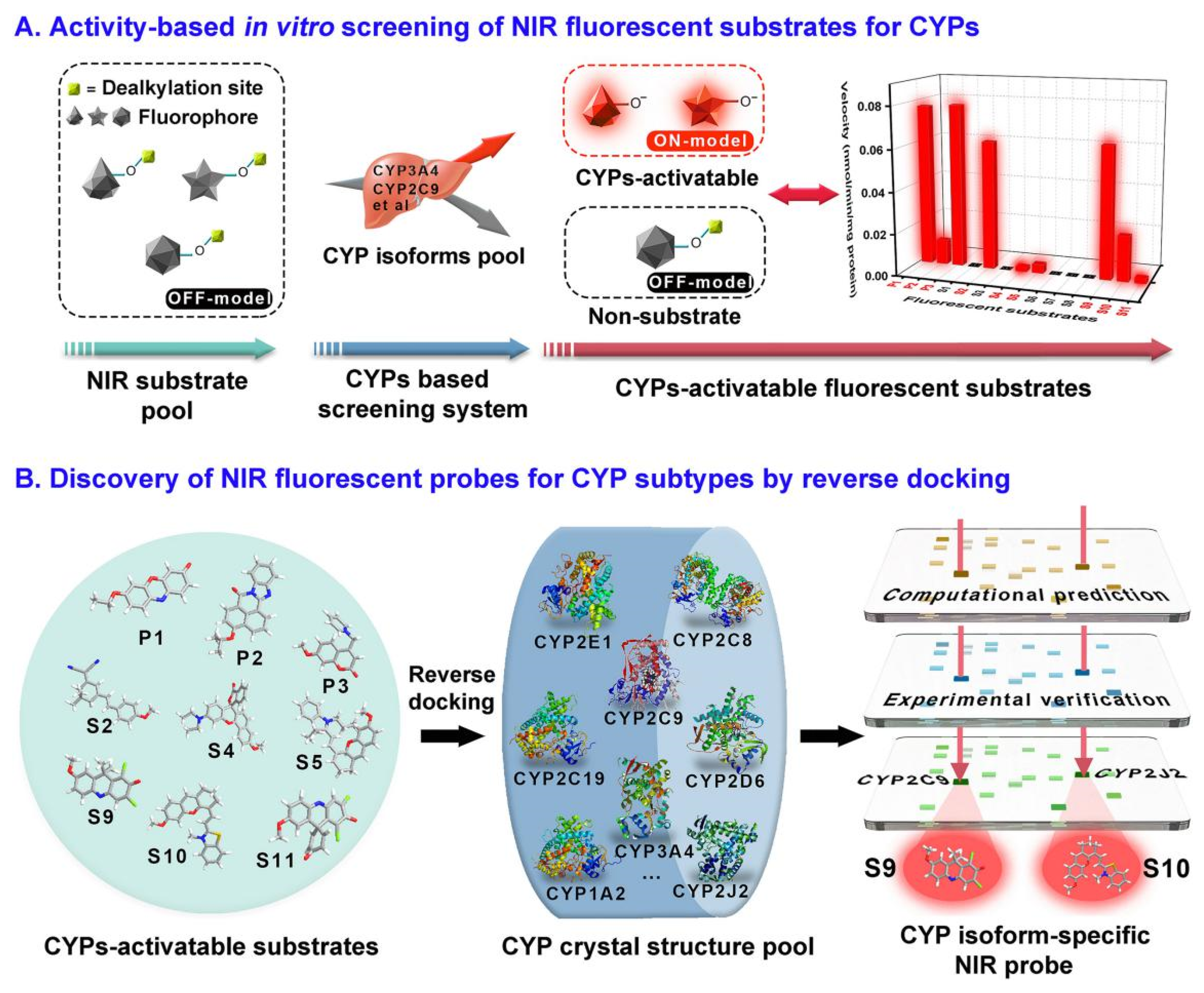
2.3. Assays for CYP450
3. Probes for CYP1 Family
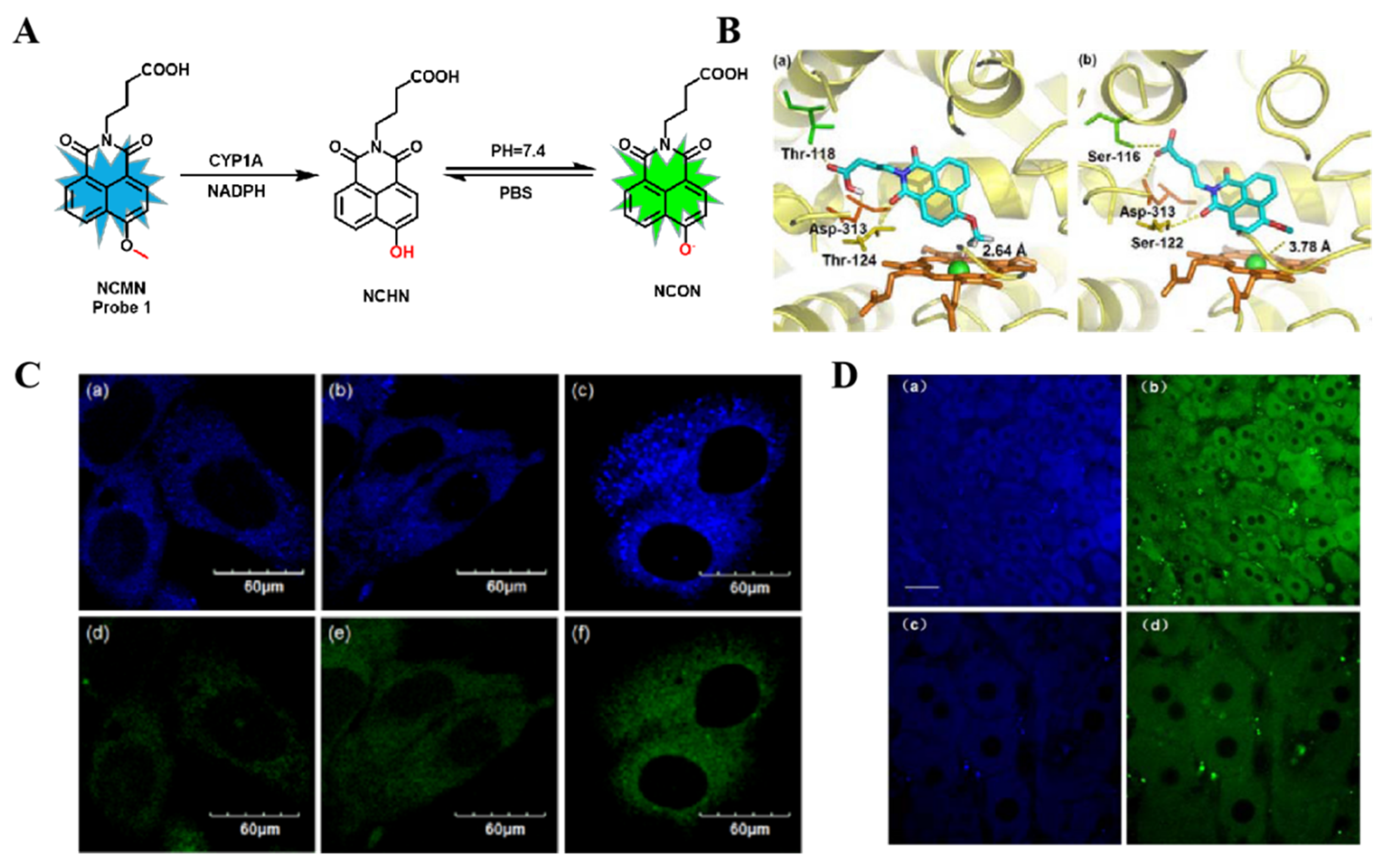
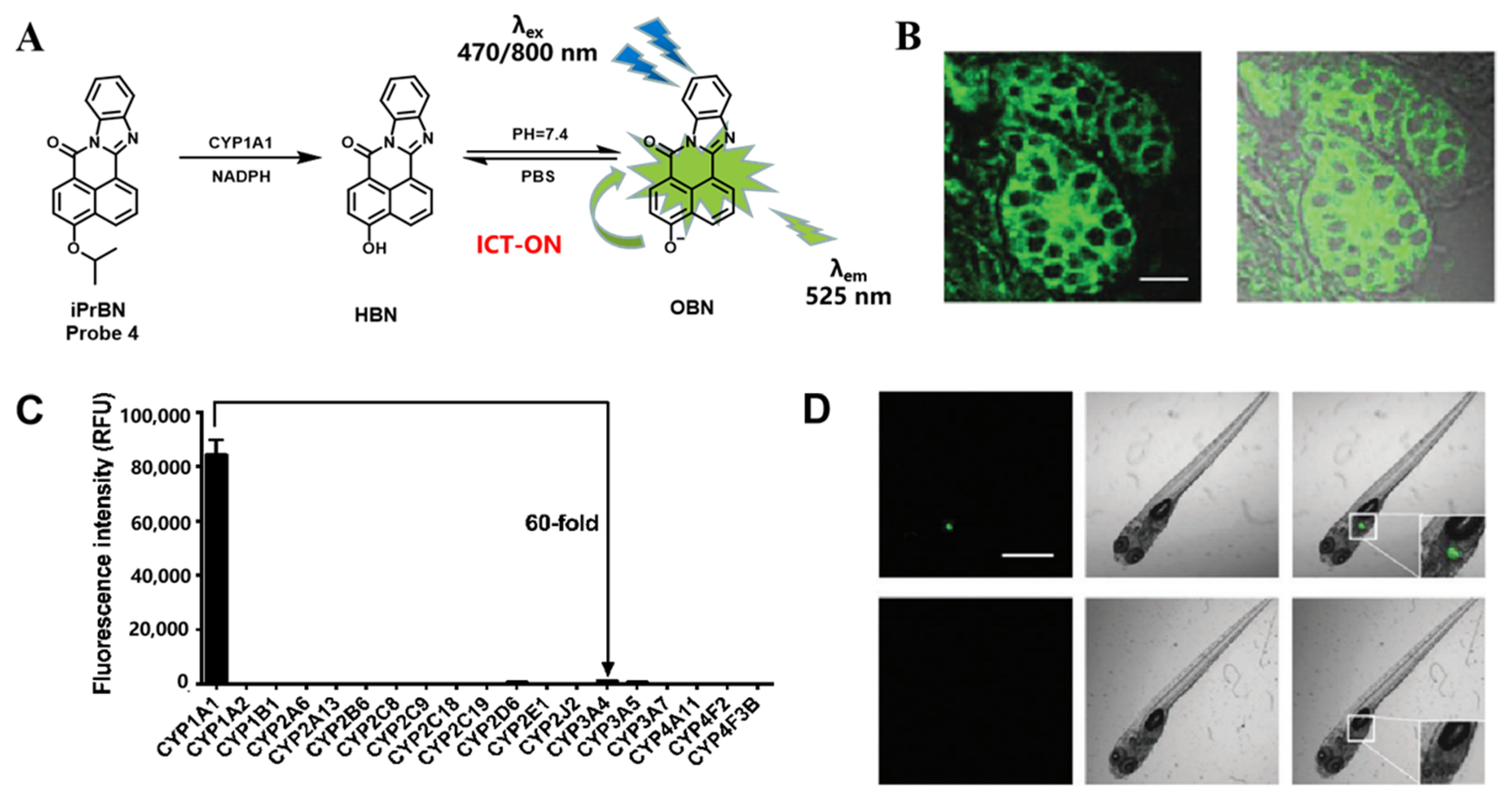
3.1. Probe(s) for CYP1A
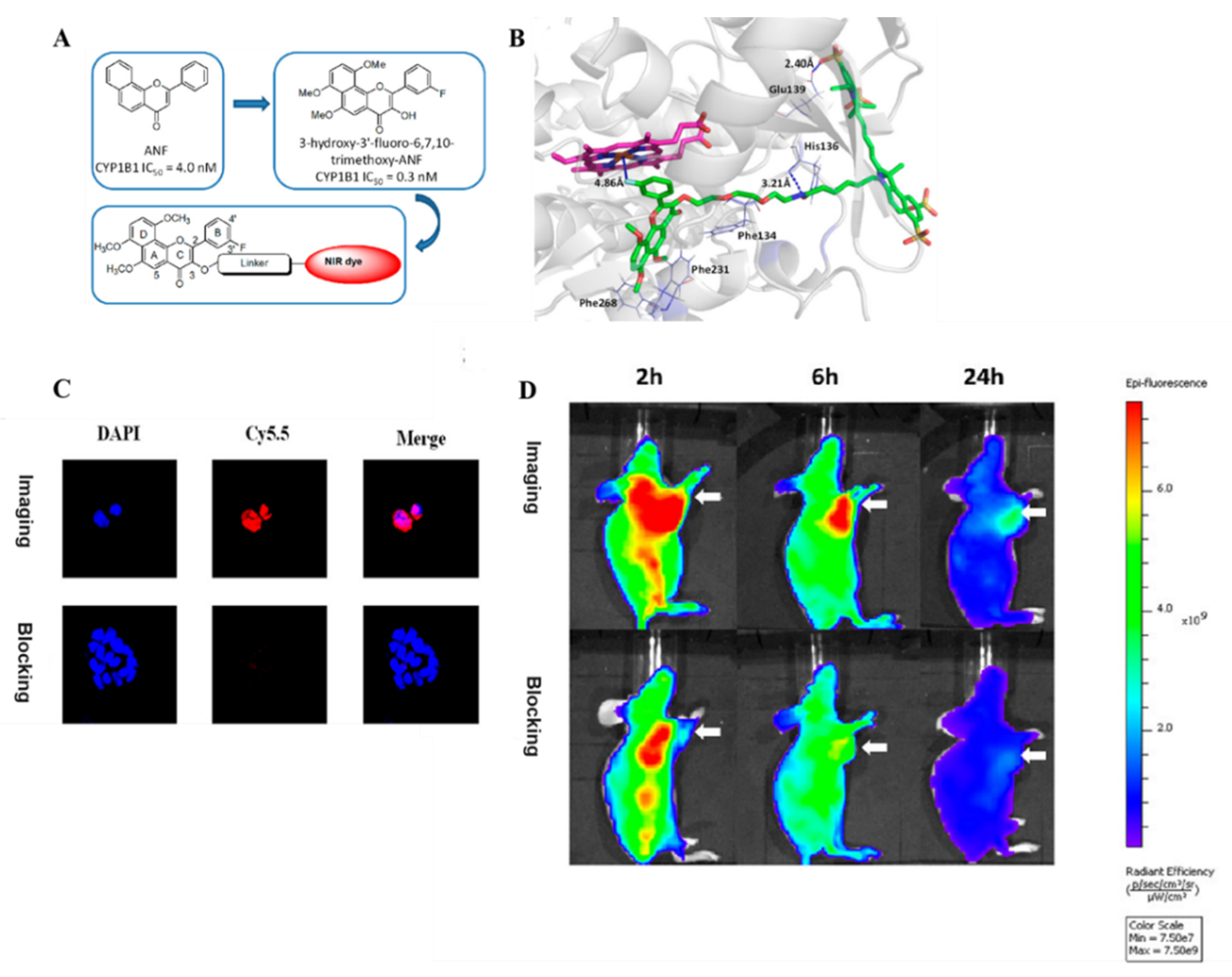
3.2. Probe(s) for CYP1B1
4. Probes for CYP2 Family
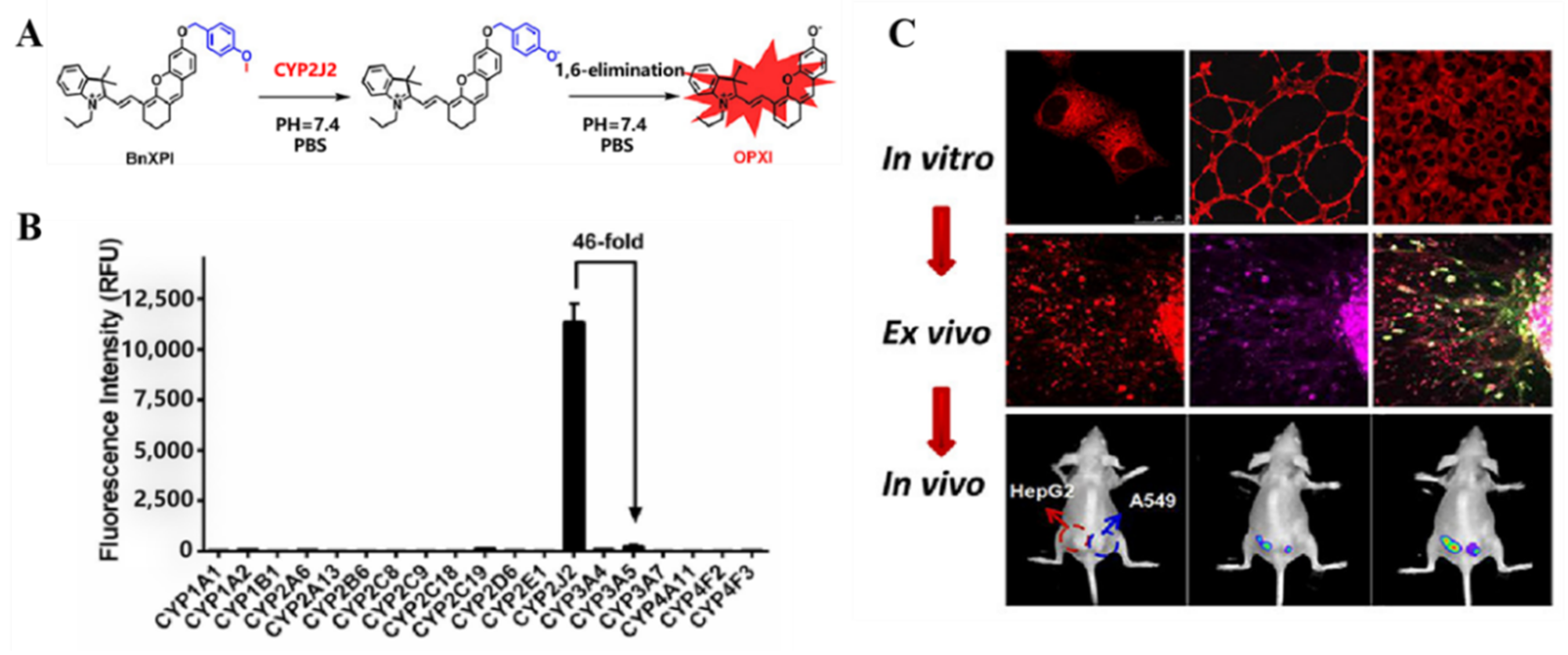
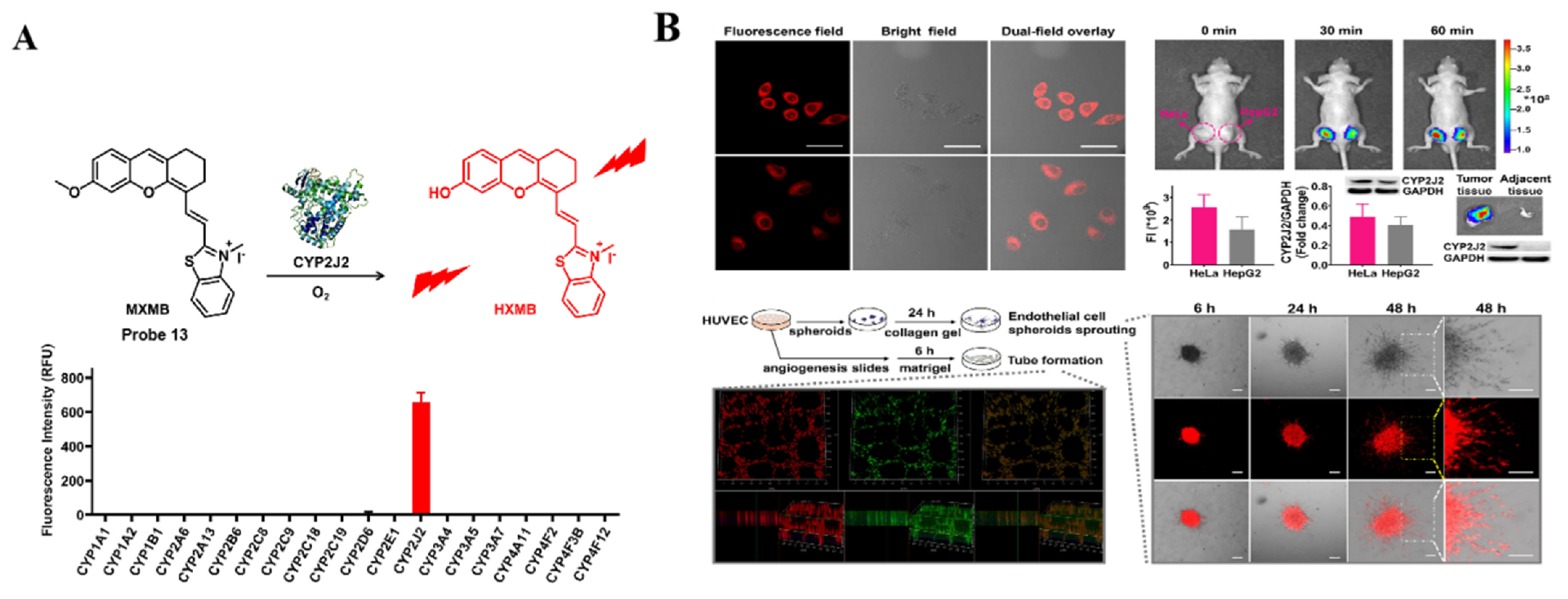
4.1. Probe(s) for CYP2J2
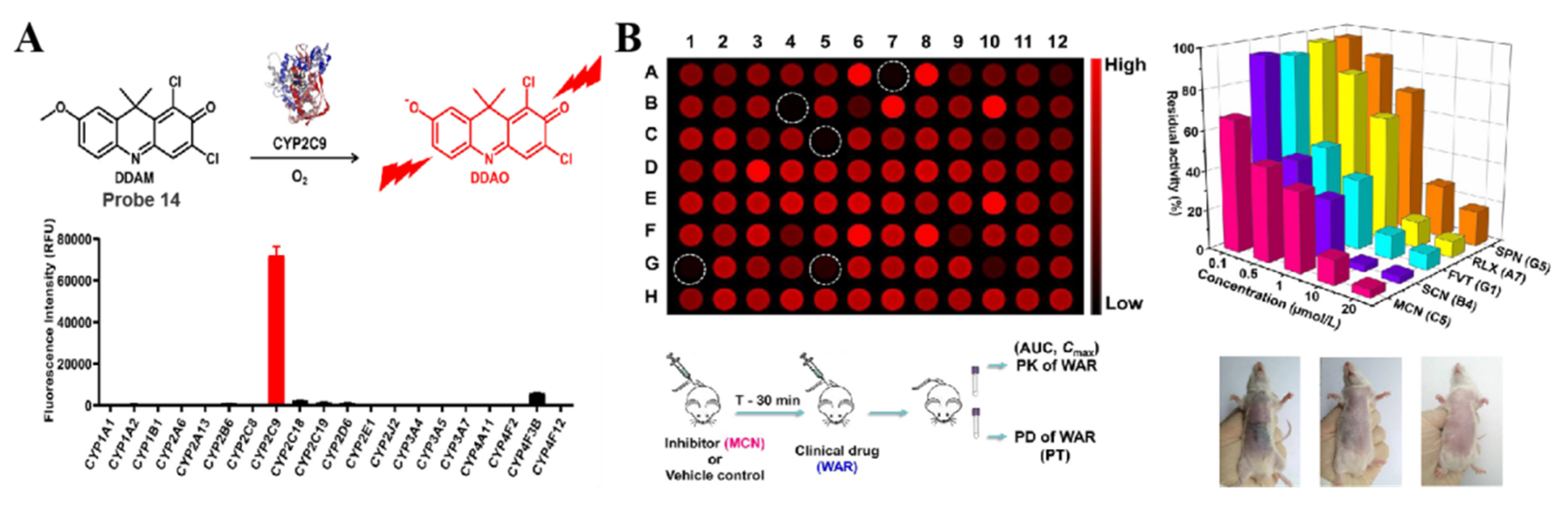
4.2. Probe(s) for CYP2C9
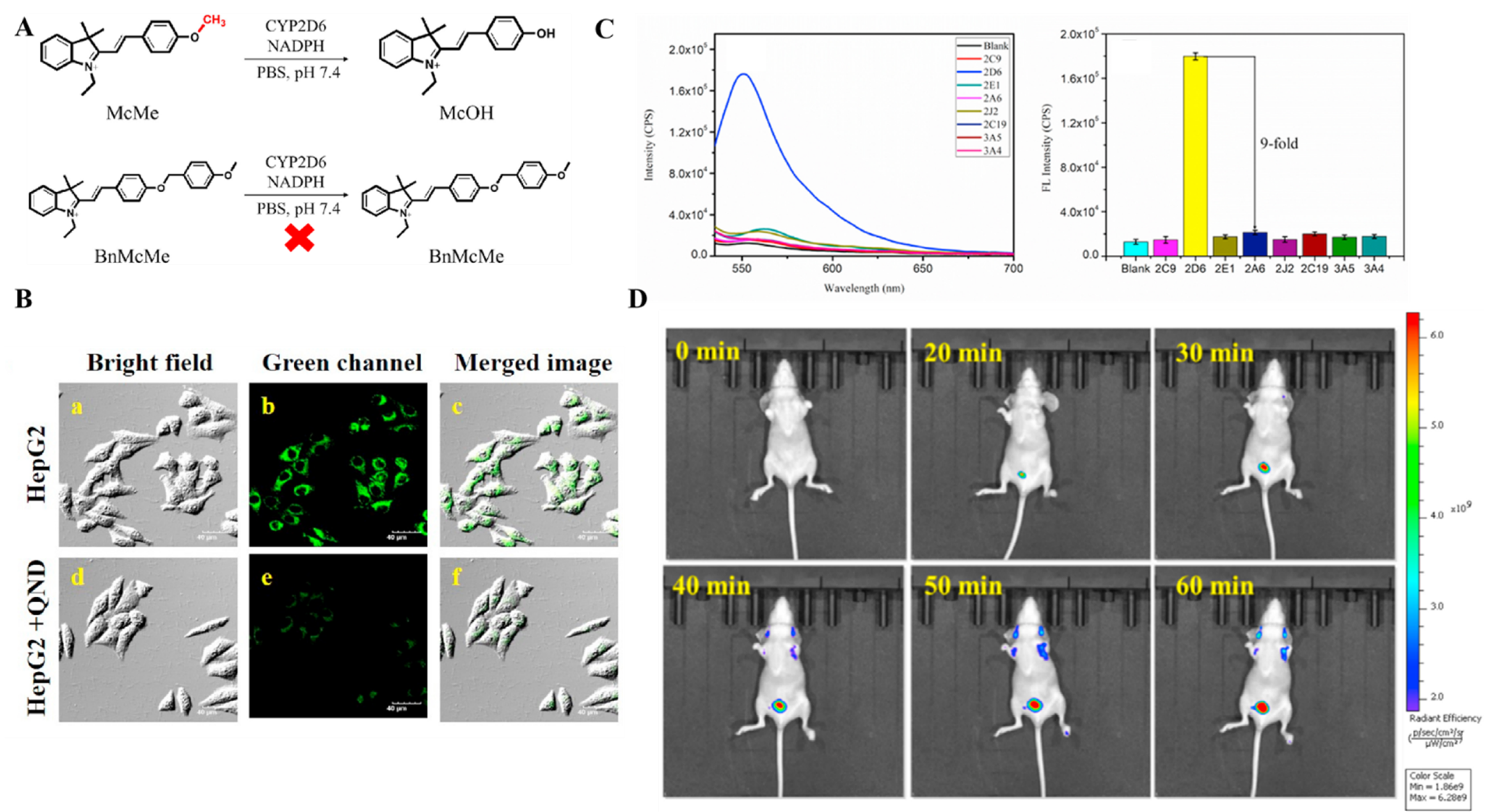
4.3. Probe(s) for CYP2D6
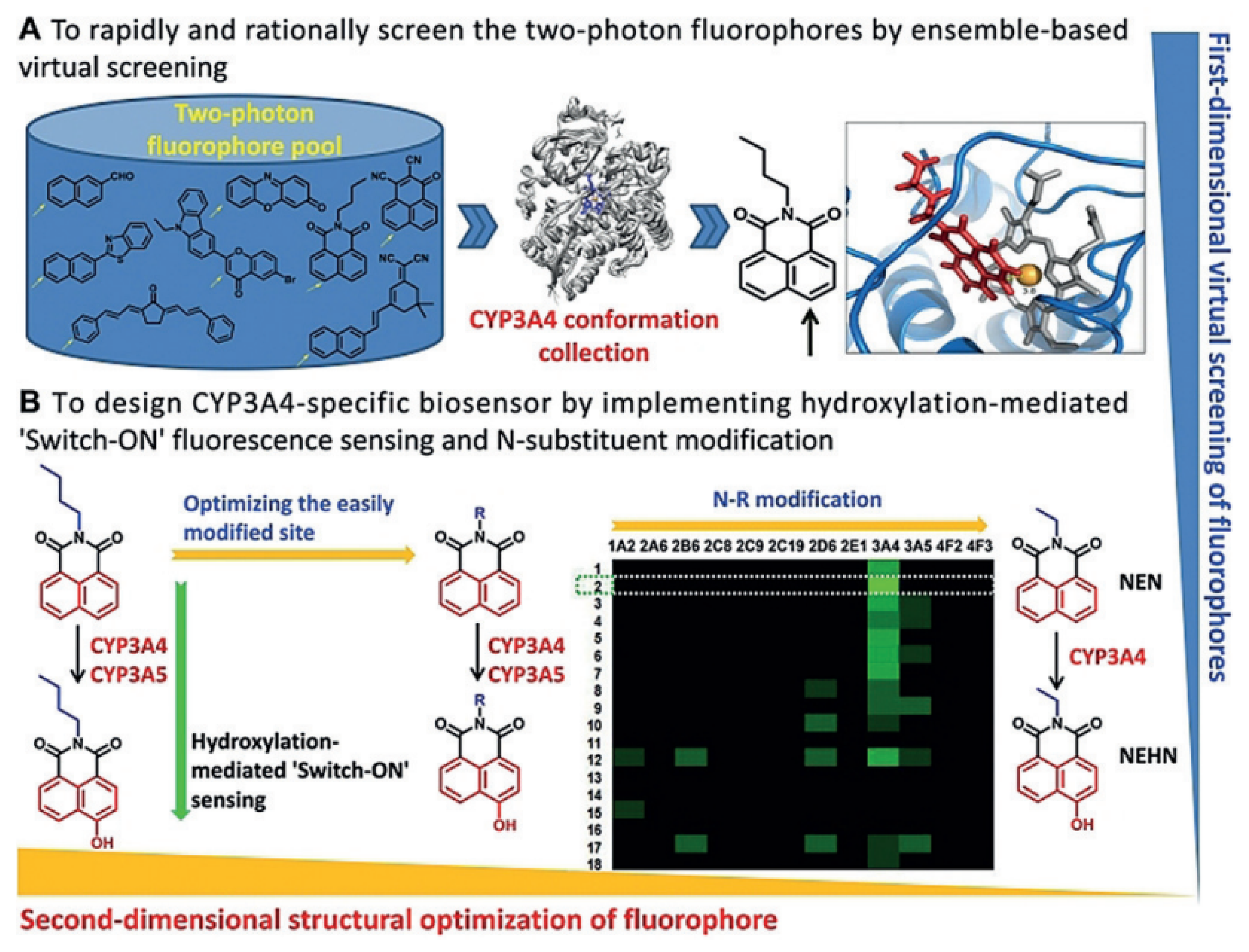
5. Probes for CYP3 Family
6. Summary and Prospects
- Development of highly selective probes for imaging of other important CYP450 isoforms. The paucity of fluorescent probes for other important CYP450 isoforms (such as CYP2A6, CYP2E1, and CYP1A2) is primarily due to the similar catalytic activities and structures among the isoforms. It is challenging to design isoform-specific probe substrates because many isoforms of the CYP450 family often have similar structures and largely overlapping substrate profiles. Other selective probe design strategies may be helpful to improve the specificity of discrimination CYP450 isomerase isoforms. For example, Ma’s team designed a highly selective monoamine oxidase A (MAO-A) probe based on a targeting moiety of utilizing MAO-A specific inhibitor to realize differentiate detection of MAO-A from MAO-B in living cells [97].
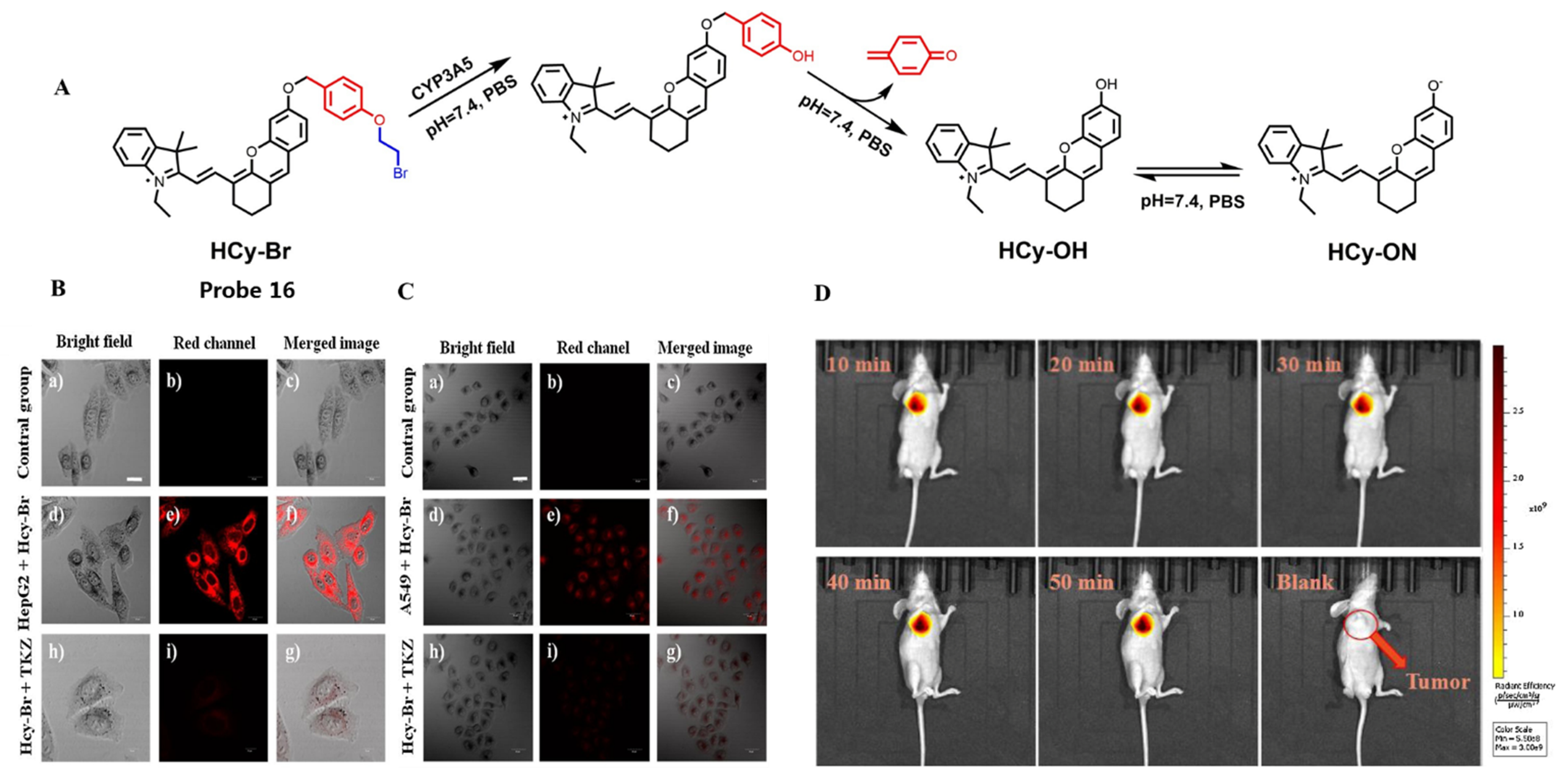
- Investigate and identify more adjustable NIR fluorophores. 1,8-Naphthylamide has evolved into the “star” dye for CYP450-specific probe design because of its exceptional photophysical characteristics. However, many probes based on 1,8-naphthylamide derivatives were insufficient for practical applications intended for in vivo environments. NIR fluorophore-based probes may well address this problem because of their capability to improve deep tissue penetration and reduce photon scattering, such as probe 17, which is able to achieve high-fidelity and high-resolution detection of CYP3A5 activity in tumor-bearing nude mice.
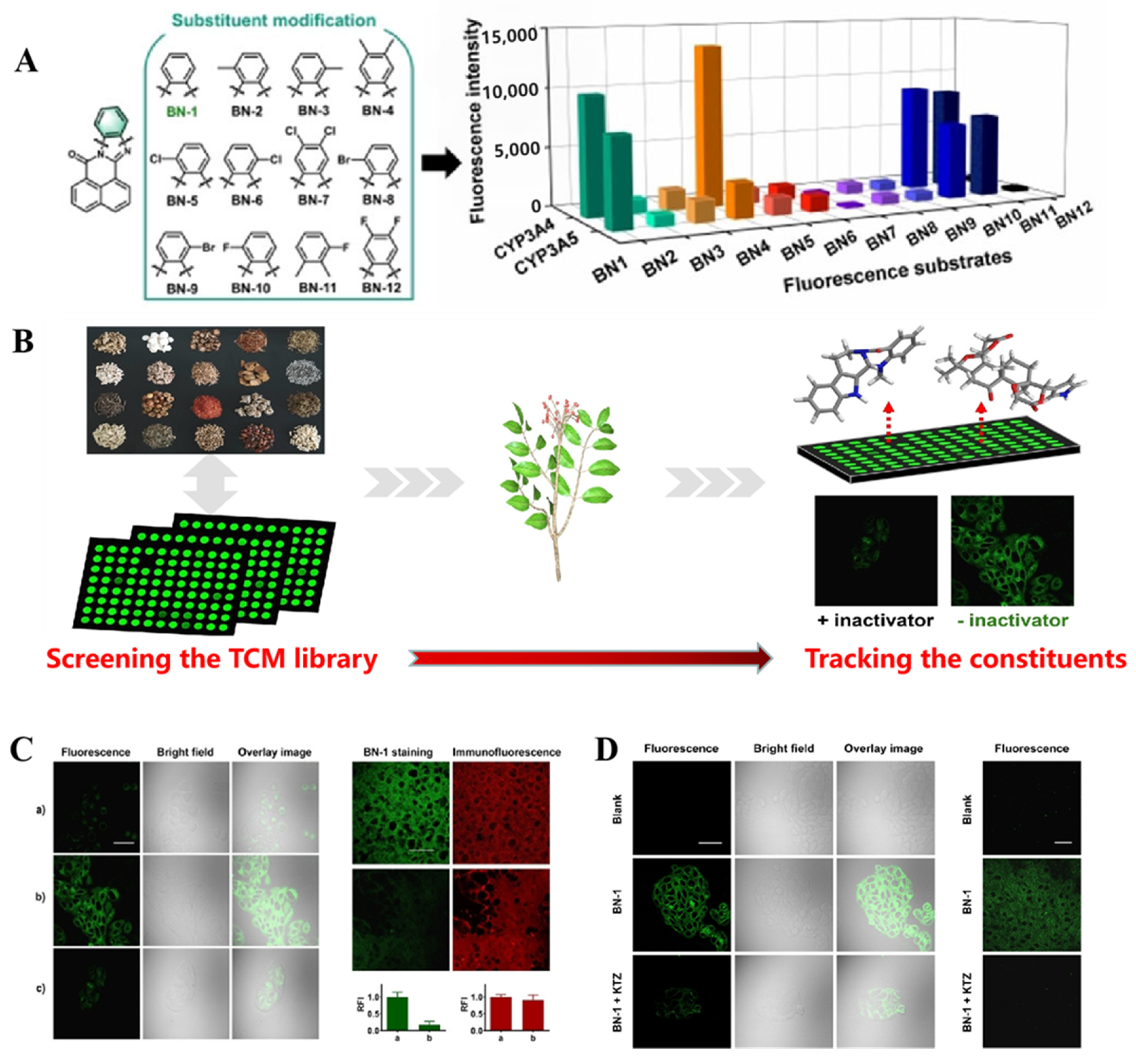
- Development of fluorescent probes with diverse recognition reactions. CYP450 is a multifunctional catalytic metabolic enzyme that can catalyze a variety of reactions in Figure 4. Nevertheless, the fluorescent probes developed now mainly rely on O-dealkylation and hydroxylation, so we may be able to develop more efficient fluorescent probes based on other CYP450-catalyzed reactions (Figure 4B), such as dealkylation of N- and S- atoms. Furthermore, numerous probe design strategies suggest that molecular docking and inhibitor biomimicry is beneficial to the development of a novel recognition site for highly selective CYP450 isoform probes.
- Discovery of probes that explore the relationship between CYP450 isoforms and their relevant carcinogens. CYP450s are involved in the metabolism of carcinogens in different types of cancer (including lung cancer, oral cancer, breast cancer, prostate cancer, hepatocellular and colorectal cancer, etc.), and the application of the above probes in this area is currently limited by the detection of expression levels in hepatocellular and breast cancers. However, other CYP450s-related diseases are seriously neglected, such as drug-induced liver injury, which is the common and the main reason for the current drug withdrawal from the market [98]. In addition, the current biological applications of CYP450 fluorescent probes are relatively simple and are limited to endogenous detection of cell tissues in mice. However, its related disease pathogenesis and its pathological role are still unclear, needing further exploration and discovery. Despite the detection and the diagnosis of many tumor mice models, there is still a long way to go before the real clinical application of CYP450 fluorescent probes.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korobkova, E.A. Effect of Natural Polyphenols on CYP Metabolism: Implications for Diseases. Chem. Res. Toxicol. 2015, 28, 1359–1390. [Google Scholar] [CrossRef] [PubMed]
- Munro, A.W.; Girvan, H.M.; Mason, A.E.; Dunford, A.J.; McLean, K.J. What makes a P450 tick? Trends Biochem. Sci. 2013, 38, 140–150. [Google Scholar] [CrossRef]
- Sangar, M.C.; Bansal, S.; Avadhani, N.C. Bimodal targeting of microsomal cytochrome P450s to mitochondria: Implications in drug metabolism and toxicity. Expert Opin. Drug Metab. Toxicol. 2010, 6, 1231–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mast, N.; Norcross, R.; Andersson, U.; Shou, M.G.; Nakayama, K.; Bjorkhem, I.; Pikuleva, I.A. Broad substrate specificity of human cytochrome P450 46A1 which initiates cholesterol degradation in the brain. Biochemistry 2003, 42, 14284–14292. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Intersection of the Roles of Cytochrome P450 Enzymes with Xenobiotic and Endogenous Substrates: Relevance to Toxicity and Drug Interactions. Chem. Res. Toxicol. 2017, 30, 2–12. [Google Scholar] [CrossRef]
- Stipp, M.C.; Acco, A. Involvement of cytochrome P450 enzymes in inflammation and cancer: A review. Cancer Chemother. Pharmacol. 2021, 87, 295–309. [Google Scholar] [CrossRef]
- Nebert, D.W.; Russell, D.W. Clinical importance of the cytochromes P450. Lancet 2002, 360, 1155–1162. [Google Scholar] [CrossRef]
- Court, M.H. Interindividual variability in hepatic drug glucuronidation: Studies into the role of age, sex, enzyme inducers, and genetic polymorphism using the human liver bank as a model system. Drug Metab. Rev. 2010, 42, 209–224. [Google Scholar] [CrossRef]
- Gong, L.; Zhang, C.-M.; Lv, J.-F.; Zhou, H.-H.; Fan, L. Polymorphisms in cytochrome P450 oxidoreductase and its effect on drug metabolism and efficacy. Pharmacogenet. Genom. 2017, 27, 337–346. [Google Scholar] [CrossRef]
- Ragia, G.; Giannakopoulou, E.; Karaglani, M.; Karantza, I.-M.; Tavridou, A.; Manolopoulos, V.G. Frequency of CYP450 enzyme gene polymorphisms in the Greek population: Review of the literature, original findings and clinical significance. Drug Metab. Drug Interact. 2014, 29, 235–248. [Google Scholar] [CrossRef]
- Gharavi, N.; El-Kadi, A.O.S. Expression of cytochrome P450 in lung tumor. Curr. Drug Metab. 2004, 5, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Sun, E.; Song, J.; Yang, L.; Zhang, Z.-H.; Ning, Q.; Jia, X.-B. [Relationship between hepatic drug-metabolizing enzymes CYP450 and traditional Chinese medicine-induced hepatotoxicity]. China J. Chin. Mater. Med. 2016, 41, 2774–2780. [Google Scholar]
- Lu, T.-l.; Su, L.-l.; Ji, D.; Gu, W.; Mao, C.-q. Interaction between CYP450 enzymes and metabolism of traditional Chinese medicine as well as enzyme activity assay. China J. Chin. Mater. Med. 2015, 40, 3524–3529. [Google Scholar]
- Wu, J.; Guan, X.; Dai, Z.; He, R.; Ding, X.; Yang, L.; Ge, G. Molecular probes for human cytochrome P450 enzymes: Recent progress and future perspectives. Coord. Chem. Rev. 2021, 427, 213600. [Google Scholar] [CrossRef]
- Feng, L.; Ning, J.; Tian, X.; Wang, C.; Yu, Z.; Huo, X.; Xie, T.; Zhang, B.; James, T.D.; Ma, X. Fluorescent probes for the detection and imaging of Cytochrome P450. Coord. Chem. Rev. 2021, 437, 213740. [Google Scholar] [CrossRef]
- Chen, J.; Fang, Y.; Sun, L.; Zeng, F.; Wu, S. An activatable probe for detecting alcoholic liver injury via multispectral optoacoustic tomography and fluorescence imaging. Chem. Comm. 2020, 56, 11102–11105. [Google Scholar] [CrossRef]
- Gao, Y.; Lin, Y.; Liu, T.; Chen, H.; Yang, X.; Tian, C.; Du, L.; Li, M. Bioluminescent Probe for Tumor Hypoxia Detection via CYP450 Reductase in Living Animals. Anal. Chem. 2017, 89, 12488–12493. [Google Scholar] [CrossRef]
- O’Connor, L.J.; Mistry, I.N.; Collins, S.L.; Folkes, L.K.; Brown, G.; Conway, S.J.; Hammond, E.M. CYP450 Enzymes Effect Oxygen-Dependent Reduction of Azide-Based Fluorogenic Dyes. ACS Cent. Sci. 2017, 3, 20–30. [Google Scholar] [CrossRef]
- Yin, F.C.; Zhang, X.; Wang, L.; Wang, Y.Q.; Zhu, Y.J.; Li, Z.Y.; Tao, T.T.; Chen, W.W.; Yu, H.; Qin, J.H. HiPSC-derived multi-organoids-on-chip system for safety assessment of antidepressant drugs. Lab Chip 2021, 21, 571–581. [Google Scholar] [CrossRef]
- Nelson, D.R. A world of cytochrome P450s. Philos. Trans. R. Soc. B 2013, 368, 20120430. [Google Scholar] [CrossRef]
- Wei, P.; Zhang, J.; Egan-Hafley, M.; Liang, S.; Moore, D.D. The nuclear receptor CAR mediates specific xenobiotic induction of drug metabolism. Nature 2000, 407, 920–923. [Google Scholar] [CrossRef] [PubMed]
- Mittal, B.; Tulsyan, S.; Kumar, S.; Mittal, R.D.; Agarwal, G. Cytochrome P450 in Cancer Susceptibility and Treatment. Adv. Clin. Chem. 2015, 71, 77–139. [Google Scholar]
- Lynch, T.; Price, A. The effect of cytochrome P450 metabolism on drug response, interactions, and adverse effects. Am. Fam. Physician 2007, 76, 391–396. [Google Scholar] [PubMed]
- Deodhar, M.; Al Rihani, S.B.; Arwood, M.J.; Darakjian, L.; Dow, P.; Turgeon, J.; Michaud, V. Mechanisms of CYP450 Inhibition: Understanding Drug-Drug Interactions Due to Mechanism-Based Inhibition in Clinical Practice. Pharmaceutics 2020, 12, 846. [Google Scholar] [CrossRef] [PubMed]
- Fontana, E.; Dansette, P.M.; Poli, S.M. Cytochrome p450 enzymes mechanism based inhibitors: Common sub-structures and reactivity. Curr. Drug Metab. 2005, 6, 413–454. [Google Scholar] [CrossRef] [PubMed]
- Silverman, R.B. Design and Mechanism of GABA Aminotransferase Inactivators. Treatments for Epilepsies and Addictions. Chem. Rev. 2018, 118, 4037–4070. [Google Scholar] [CrossRef] [PubMed]
- Munro, A.W.; Girvan, H.M.; McLean, K.J. Variations on a (t)heme--novel mechanisms, redox partners and catalytic functions in the cytochrome P450 superfamily. Nat. Prod. Rep. 2007, 24, 585–609. [Google Scholar] [CrossRef]
- Schlichting, I.; Berendzen, J.; Chu, K.; Stock, A.M.; Maves, S.A.; Benson, D.E.; Sweet, B.M.; Ringe, D.; Petsko, G.A.; Sligar, S.G. The catalytic pathway of cytochrome P450cam at atomic resolution. Science 2000, 287, 1615–1622. [Google Scholar] [CrossRef]
- Guengerich, F.P. Common and Uncommon Cytochrome P450 Reactions Related to Metabolism and Chemical Toxicity. Chem. Res. Toxicol. 2001, 14, 611–650. [Google Scholar] [CrossRef] [PubMed]
- Isin, E.M.; Guengerich, F.P. Complex reactions catalyzed by cytochrome P450 enzymes. Biochim. Biophys. Acta 2007, 1770, 314–329. [Google Scholar] [CrossRef]
- Bernhardt, R. Cytochromes P450 as versatile biocatalysts. J. Biotechnol. 2006, 124, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Shaik, S.; Kumar, D.; de Visser, S.P.; Altun, A.; Thiel, W. Theoretical Perspective on the Structure and Mechanism of Cytochrome P450 Enzymes. Chem. Rev. 2005, 105, 2279–2328. [Google Scholar] [CrossRef] [PubMed]
- Ung, Y.T.; Ong, C.E.; Pan, Y. Current High-Throughput Approaches of Screening Modulatory Effects of Xenobiotics on Cytochrome P450 (CYP) Enzymes. High-Throughput 2018, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlokarnik, G.; Grootenhuis, P.D.; Watson, J.B. High throughput P450 inhibition screens in early drug discovery. Drug Discov. Today 2005, 10, 1443–1450. [Google Scholar] [CrossRef]
- Zhang, H.; Fan, J.; Wang, J.; Dou, B.; Zhou, F.; Cao, J.; Qu, J.; Cao, Z.; Zhao, W.; Peng, X. Fluorescence discrimination of cancer from inflammation by molecular response to COX-2 enzymes. J. Am. Chem. Soc. 2013, 135, 17469–17475. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tian, X.; Shin, I.; Yoon, J. Fluorescent and luminescent probes for detection of reactive oxygen and nitrogen species. Chem. Soc. Rev. 2011, 40, 4783–4804. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Peng, J.; Lv, Y.; Su, D.; Liu, D.; Chen, M.; Yuan, L.; Zhang, X. De Novo Design of Chemical Stability Near-Infrared Molecular Probes for High-Fidelity Hepatotoxicity Evaluation In Vivo. J. Am. Chem. Soc. 2019, 141, 6352–6361. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zeng, Z.; Jiang, J.H.; Chang, Y.T.; Yuan, L. Discerning the Chemistry in Individual Organelles with Small-Molecule Fluorescent Probes. Angew. Chem. Int. Ed. 2016, 55, 13658–13699. [Google Scholar] [CrossRef]
- Chauret, N.; Tremblay, N.; Lackman, R.L.; Gauthier, J.-Y.; Silva, J.M.; Marois, J.; Yergey, J.A.; Nicoll-Griffith, D.A. Description of a 96-Well Plate Assay to Measure Cytochrome P4503A Inhibition in Human Liver Microsomes Using a Selective Fluorescent Probe. Anal. Biochem. 1999, 276, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Li, M.; Lv, S.; Zhang, K.; Lu, M.; Tang, D. In situ synthesis of fluorescent polydopamine nanoparticles coupled with enzyme-controlled dissolution of MnO2 nanoflakes for a sensitive immunoassay of cancer biomarkers. J. Mater. Chem. 2017, 5, 8506–8513. [Google Scholar] [CrossRef]
- Lv, S.; Tang, Y.; Zhang, K.; Tang, D. Wet NH3-Triggered NH2-MIL-125(Ti) Structural Switch for Visible Fluorescence Immunoassay Impregnated on Paper. Anal. Chem. 2018, 90, 14121–14125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Zhu, L.; Lv, Z.; Li, M.; Tang, D. Persistent luminescence nanorods-based autofluorescence-free biosensor for prostate-specific antigen detection. Talanta 2021, 233, 122563. [Google Scholar] [CrossRef] [PubMed]
- Hasler, J.A.; Estabrook, R.; Murray, M.; Pikuleva, I.; Waterman, M.; Capdevila, J.; Holla, V.; Helvig, C.; Falck, J.R.; Farrell, G.; et al. Human cytochromes P450. Mol. Aspects Med. 1999, 20, 1–137. [Google Scholar] [CrossRef]
- Bulus, H.; Oguztuzun, S.; Simsek, G.G.; Kilic, M.; Ada, A.O.; Gol, S.; Kocdogan, A.K.; Kaygin, P.; Bozer, B.; Iscan, M. Expression of CYP and GST in human normal and colon tumor tissues. Biotech. Histochem. 2019, 94, 1–9. [Google Scholar] [CrossRef]
- Lang, D.; Radtke, M.; Bairlein, M. Highly Variable Expression of CYP1A1 in Human Liver and Impact on Pharmacokinetics of Riociguat and Granisetron in Humans. Chem. Res. Toxicol. 2019, 32, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Fujii-Kuriyama, Y. Metabolic activation of polycyclic aromatic hydrocarbons to carcinogens by cytochromes P450 1A1 and1B1. Cancer Sci. 2004, 95, 1–6. [Google Scholar] [CrossRef]
- Roberts, E.A.; Xie, Z.W.; Yang, S.; Lipa, J. Inducibility of enzyme activities associated with the cytochrome P-450 1A family, ethoxyresorufin O-deethylase, and methoxyresorufin O-demethylase in human hepatocyte lines derived from normal liver tissue. Drug Metab. Dispos.. 1993, 21, 56–61. [Google Scholar]
- Zhang, C.; Ren, A.M.; Guo, J.F.; Wang, D.; Yu, L.Y. Theoretical design and investigation of 1,8-naphthalimide-based two-photon fluorescent probes for detecting cytochrome P450 1A with separated fluorescence signal. Phys. Chem. Chem. Phys. 2018, 20, 13290–13305. [Google Scholar] [CrossRef]
- Dai, Z.R.; Ge, G.B.; Feng, L.; Ning, J.; Hu, L.H.; Jin, Q.; Wang, D.D.; Lv, X.; Dou, T.Y.; Cui, J.N.; et al. A Highly Selective Ratiometric Two-Photon Fluorescent Probe for Human Cytochrome P450 1A. J. Am. Chem. Soc. 2015, 137, 14488–14495. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, Y.; Gu, X.; Cheng, Y.; Hong, M.; Yan, L.; Ma, F.; Qi, Z. Synthesis of a selective ratiometric fluorescent probe based on Naphthalimide and its application in human cytochrome P450 1A. Talanta 2018, 186, 413–420. [Google Scholar] [CrossRef]
- Hammons, G.J.; Milton, D.; Stepps, K.; Guengerich, F.P.; Tukey, R.H.; Kadlubar, F.F. Metabolism of carcinogenic heterocyclic and aromatic amines by recombinant human cytochrome P450 enzymes. Carcinogenesis 1997, 18, 851–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, D.; Wu, B.; Chow, D.; Hu, M. Substrate selectivity of drug-metabolizing cytochrome P450s predicted from crystal structures and in silico modeling. Drug Metab. Rev. 2012, 44, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.R.; Feng, L.; Jin, Q.; Cheng, H.L.; Li, Y.; Ning, J.; Yu, Y.; Ge, G.B.; Cui, J.N.; Yang, L. A practical strategy to design and develop an isoform-specific fluorescent probe for a target enzyme: CYP1A1 as a case study. Chem. Sci. 2017, 8, 2795–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q. Induction of CYP1A1. The AhR/DRE paradigm: Transcription, receptor regulation, and expanding biological roles. Curr. Drug Metab. 2001, 2, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Wohak, L.E.; Krais, A.M.; Kucab, J.E.; Stertmann, J.; Ovrebo, S.; Seidel, A.; Phillips, D.H.; Arlt, V.M. Carcinogenic polycyclic aromatic hydrocarbons induce CYP1A1 in human cells via a p53-dependent mechanism. Arch. Toxicol. 2016, 90, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Ning, J.; Tian, Z.H.; Wang, B.; Ge, G.B.; An, Y.; Hou, J.; Wang, C.; Zhao, X.Y.; Li, Y.N.; Tian, X.G.; et al. A highly sensitive and selective two-photon fluorescent probe for real-time sensing of cytochrome P450 1A1 in living systems. Mater. Chem. Front. 2018, 2, 2013–2020. [Google Scholar] [CrossRef]
- Xue, T.Z.; Dai, Y.P.; Zhang, X.X.; Cheng, Y.; Gu, X.F.; Ji, H.F.; Misal, S.; Qi, Z.J. Ultrasensitive near-infrared fluorescent probe with large stokes shift for real-time tracing of CYP1A1 in living cells and zebrafish model. Sens. Actuators B 2019, 293, 265–272. [Google Scholar] [CrossRef]
- Ji, H.F.; Zhang, X.X.; Dai, Y.P.; Xue, T.Z.; Misal, S.; Qi, Z.J. A highly selective ratiometric fluorescent probe based on naphthalimide for detection and imaging of CYP1A1 in living cells and zebrafish. Analyst 2019, 144, 7390–7397. [Google Scholar] [CrossRef]
- Jin, Q.; Ma, H.Y.; Feng, L.; Wang, P.; He, R.J.; Ning, J.; Yang, L.; Ge, G.B. Sensing cytochrome P450 1A1 activity by a resorufin-based isoform-specific fluorescent probe. Chin. Chem. Lett. 2020, 31, 2945–2949. [Google Scholar] [CrossRef]
- Huang, Y.; Yu, L.; Lu, P.P.; Wei, Y.H.; Wang, X.Y.; Chen, L.X. Evaluate the inhibition of cytochrome P450 1A1 for enhancing breast cancer chemotherapy with a turn-on fluorescent probe. Sens. Actuators B 2021, 344, 130233. [Google Scholar] [CrossRef]
- Ozeki, J.; Uno, S.; Ogura, M.; Choi, M.; Maeda, T.; Sakurai, K.; Matsuo, S.; Amano, S.; Nebert, D.W.; Makishima, M. Aryl hydrocarbon receptor ligand 2,3,7,8-tetrachlorodibenzo-p-dioxin enhances liver damage in bile duct-ligated mice. Toxicology 2011, 280, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Berg, M.; Birnbaum, L.S.; Denison, M.; De Vito, M.; Farland, W.; Feeley, M.; Fiedler, H.; Hakansson, H.; Hanberg, A.; Haws, L.; et al. The 2005 World Health Organization Reevaluation of Human and Mammalian Toxic Equivalency Factors for Dioxins and Dioxin-Like Compounds. Toxicol. Sci. 2006, 93, 223–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, B.C.; Mackenzie, P.I.; Miners, J.O. Comparative homology modeling of human cytochrome P4501A1 (CYP1A1) and confirmation of residues involved in 7-ethoxyresorufin O-deethylation by site-directed mutagenesis and enzyme kinetic analysis. Arch. Biochem. Biophys. 2007, 468, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, A.P.; Kaminski, D.L.; Ruh, M.F. 2,3,7,8 Tetrachlorodibenzo-p-dioxin induction of cytochrome P4501A in cultured rat and human hepatocytes. Chem.-Biol. Interact. 2000, 124, 173–189. [Google Scholar] [CrossRef]
- Shangguan, L.; Wei, Y.; Wang, K.; Zhang, Y.; Liu, S. Highly sensitive fluorescent bioassay of 2,3,7,8-tetrachloro-dibenzo-p-dioxin based on abnormal expression of cytochrome P450 1A2 in human cells. Anal. Chim. Acta 2019, 1046, 179–184. [Google Scholar] [CrossRef]
- Li, F.; Zhu, W.F.; Gonzalez, F.J. Potential role of CYP1B1 in the development and treatment of metabolic diseases. Pharmacol. Ther. 2017, 178, 18–30. [Google Scholar] [CrossRef]
- Meng, Q.Q.; Wang, Z.T.; Cui, J.H.; Cui, Q.; Dong, J.Y.; Zhang, Q.J.; Li, S.S. Design, Synthesis, and Biological Evaluation of Cytochrome P450 1B1 Targeted Molecular Imaging Probes for Colorectal Tumor Detection. J. Med. Chem. 2018, 61, 10901–10909. [Google Scholar] [CrossRef]
- Pozzi, A.; Macias-Perez, I.; Abair, T.; Wei, S.Z.; Su, Y.; Zent, R.; Falck, J.R.; Capdevila, J.H. Characterization of 5,6-and 8,9-epoxyeicosatrienoic acids (5,6-and 8,9-EET) as potent in vivo angiogenic lipids. J. Biol. Chem. 2005, 280, 27138–27146. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, S.; Yamazoe, Y. Involvement of multiple human cytochromes P450 in the liver microsomal metabolism of astemizole and a comparison with terfenadine. Br. J. Clin. Pharmacol. 2001, 51, 133–142. [Google Scholar]
- Ning, J.; Liu, T.; Dong, P.P.; Wang, W.; Ge, G.B.; Wang, B.; Yu, Z.L.; Shi, L.; Tian, X.G.; Huo, X.K.; et al. Molecular Design Strategy to Construct the Near-Infrared Fluorescent Probe for Selectively Sensing Human Cytochrome P450 2J2. J. Am. Chem. Soc. 2019, 141, 1126–1134. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Xue, T.; Ji, H.; Zhang, P.; Zhang, D.; Qi, Z. In situ target enzyme-activated near-infrared fluorescent probe: A case study of CYP2J2 using three-fragmentary molecular assembly engineering. Sens. Actuators B 2021, 328, 129034. [Google Scholar] [CrossRef]
- Feng, L.; Tian, X.G.; Yao, D.H.; Yu, Z.L.; Huo, X.K.; Tian, Z.H.; Ning, J.; Cui, J.N.; James, T.D.; Ma, X.C. A practical strategy to develop isoform-selective near-infrared fluorescent probes for human cytochrome P450 enzymes. Acta Pharm. Sin. B 2022, 12, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, S.; Holm, L.; Myrberg, O.; Sundstrom, A.; Yue, Q.Y. Noscapine may increase the effect of warfarin. Br. J. Clin. Pharmacol. 2008, 65, 277–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenborg, S.; Stenberg, M.; Otto, S.; Ostervall, J.; Masquelier, M.; Yue, Q.Y.; Bertilsson, L.; Eliasson, E. Clinically significant CYP2C inhibition by noscapine but not by glucosamine. Clin. Pharmacol. Ther. 2010, 88, 343–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Yang, L.P.; Zhang, X.Z.; Huang, S.Q.; Bartlam, M.; Zhou, S.F. New insights into the structural characteristics and functional relevance of the human cytochrome P450 2D6 enzyme. Drug Metab. Rev. 2009, 41, 573–643. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Li, Q.; Wang, R.; Guo, Y.; Wen, S.; Xu, L.; Wang, S. Relationship of CYP2D6 genetic polymorphisms and the pharmacokinetics of tramadol in Chinese volunteers. J. Clin. Pharm. Ther. 2010, 35, 239–247. [Google Scholar] [CrossRef]
- Xu, S.Q.; Xing, X.Y.; Liu, Y.Y.; Gao, H. A hemicyanine-based fluorescence probe for selective detection of CYP2D6 in living cells and tumor-bearing mice. Dyes Pigm. 2022, 198, 109959. [Google Scholar] [CrossRef]
- Shimada, T. Special issue on human cytochromes P450 (human CYPs): Human cytochrome P450 enzymes, a status report summarizing their reactions, substrates, inducers, and inhibitors - 1st update - Concluding remarks. Drug Metab. Rev. 2002, 34, 449–450. [Google Scholar] [CrossRef]
- Mao, J.L.; Mohutsky, M.A.; Harrelson, J.P.; Wrighton, S.A.; Hall, S.D. Prediction of CYP3A-Mediated Drug-Drug Interactions Using Human Hepatocytes Suspended in Human Plasma. Drug Metab. Dispos.. 2011, 39, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Guengerich, F.P.; Kaminsky, L.S. 16Alpha-hydroxylation of estrone by human cytochrome P4503A4/5. Carcinogenesis 1998, 19, 867–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, J.N.; Lajoie, J.; Zijenah, L.S.; Barama, A.; Poirier, C.; Ward, B.J.; Roger, M. Cyp3A5 genetic polymorphisms in different ethnic populations. Drug Metab. Dispos.. 2005, 33, 884–887. [Google Scholar] [CrossRef] [PubMed]
- Kuehl, P.; Zhang, J.; Lin, Y.; Lamba, J.; Assem, M.; Schuetz, J.; Watkins, P.B.; Daly, A.; Wrighton, S.A.; Hall, S.D.; et al. Sequence diversity in CYP3A promoters and characterization of the genetic basis of polymorphic CYP3A5 expression. Nat. Genet. 2001, 27, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P. Cytochrome P-450 3A4: Regulation and role in drug metabolism. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 1–17. [Google Scholar] [CrossRef]
- Chougnet, A.; Stoessel, C.; Woggon, W.D. Design and synthesis of a new fluorescent probe for cytochrome P450 3A4 (CYP 3A4). Bioorg. Med. Chem. Lett. 2003, 13, 3643–3645. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.B.; Ning, J.; Hu, L.H.; Dai, Z.R.; Hou, J.; Cao, Y.F.; Yu, Z.W.; Ai, C.Z.; Gu, J.K.; Ma, X.C.; et al. A highly selective probe for human cytochrome P450 3A4: Isoform selectivity, kinetic characterization and its applications. Chem. Commun. 2013, 49, 9779–9781. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Wang, W.; Ge, G.B.; Chu, P.; Long, F.D.; Yang, Y.L.; Peng, Y.L.; Feng, L.; Ma, X.C.; James, T.D. Target Enzyme-Activated Two-Photon Fluorescent Probes: A Case Study of CYP3A4 Using a Two-Dimensional Design Strategy. Angew. Chem. Int. Ed. 2019, 58, 9959–9963. [Google Scholar] [CrossRef] [PubMed]
- Downie, D.; McFadyen, M.C.; Rooney, P.H.; Cruickshank, M.E.; Parkin, D.E.; Miller, I.D.; Telfer, C.; Melvin, W.T.; Murray, G.I. Profiling cytochrome P450 expression in ovarian cancer: Identification of prognostic markers. Clin. Cancer Res. 2005, 11, 7369–7375. [Google Scholar] [CrossRef] [Green Version]
- Kumarakulasingham, M.; Rooney, P.H.; Dundas, S.R.; Telfer, C.; Melvin, W.T.; Curran, S.; Murray, G.I. Cytochrome p450 profile of colorectal cancer: Identification of markers of prognosis. Clin. Cancer Res. 2005, 11, 3758–3765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.J.; Cao, Y.F.; Feng, L.; He, Y.Q.; Hong, J.Y.; Dou, T.Y.; Wang, P.; Hao, D.C.; Ge, G.B.; Yang, L. A Naturally Occurring Isoform-Specific Probe for Highly Selective and Sensitive Detection of Human Cytochrome P450 3A5. J. Med. Chem. 2017, 60, 3804–3813. [Google Scholar] [CrossRef]
- Ji, H.F.; Ma, F.L.; Dai, Y.P.; Zhao, X.X.; Xue, K.; Misal, S.; Zhang, P.; Qi, Z.J.; Zhu, H.Y. A near-infrared ratiometric fluorescent probe for selectively tracking human cytochrome P450 3A5 in living cells and tumor-bearing mice. Sens. Actuators B 2021, 331, 129372. [Google Scholar] [CrossRef]
- Dohnal, V.; Wu, Q.H.; Kuca, K. Metabolism of aflatoxins: Key enzymes and interindividual as well as interspecies differences. Arch. Toxicol. 2014, 88, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Bissada, J.E.; Truong, V.; Abouda, A.A.; Wines, K.J.; Crouch, R.D.; Jackson, K.D. Interindividual Variation in CYP3A Activity Influences Lapatinib Bioactivation. Drug Metab. Dispos. 2019, 47, 1257–1269. [Google Scholar] [CrossRef]
- Shirasaka, Y.; Chang, S.Y.; Grubb, M.F.; Peng, C.C.; Thummel, K.E.; Isoherranen, N.; Rodrigues, A.D. Effect of CYP3A5 Expression on the Inhibition of CYP3A-Catalyzed Drug Metabolism: Impact on Modeling CYP3A-Mediated Drug-Drug Interactions. Drug Metab. Dispos. 2013, 41, 1566–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, J.; Tian, Z.H.; Wang, J.Y.; Wang, B.; Tian, X.G.; Yu, Z.L.; Huo, X.K.; Feng, L.; Cui, J.N.; James, T.D.; et al. Rational Design of a Two-Photon Fluorescent Probe for Human Cytochrome P450 3A and the Visualization of Mechanism-Based Inactivation. Angew. Chem. Int. Ed. 2022, 61, e202113191. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Viloca, M.; Gao, J.; Karplus, M.; Truhlar, D.G. How enzymes work: Analysis by modern rate theory and computer simulations. Science 2004, 303, 186–195. [Google Scholar] [CrossRef]
- Wu, X.; Shi, W.; Li, X.; Ma, H. A Strategy for Specific Fluorescence Imaging of Monoamine Oxidase A in Living Cells. Angew. Chem. Int. Ed. 2017, 56, 15319–15323. [Google Scholar] [CrossRef]
- Lee, W.M.; Larrey, D.; Olsson, R.; Lewis, J.H.; Keisu, M.; Auclert, L.; Sheth, S. Hepatic Findings in Long-Term Clinical Trials of Ximelagatran. Drug Saf. 2005, 28, 351–370. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Subfamily | Gene Identifier |
|---|---|---|
| CYP1 | CYP1A | CYP1A1 |
| CYP1A2 | ||
| CYP2 | CYP2A | CYP2A1 |
| CYP2B | CYP2B1, CYP2B6 | |
| CYP2C | CYP2C8, CYP2C9, CYP2C18, CYP2C19 | |
| CYP2D | CYP2D6 | |
| CYP2E | CYP2E1 | |
| CYP2J | CYP2J2 | |
| CYP3 | CYP3A | CYP3A4, CYP3A5, CYP3A7 |
| Bio-Marker | Name | Structure a | LOD/nM | Km/μM | Ex/Em b nm/nm | Biological Application |
|---|---|---|---|---|---|---|
| CYP1A | NCMN Probe 1 [49] |  | 50 | 11.9 | 450/564 | TPM c imaging identifies CYP1A induced by different concentrations of inducer and 3D Depth Imaging of CYP1A in rat liver tissues |
| NEMN Probe 2 [50] | 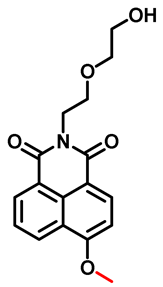 | 8.10/ 15.75 | / | 446/552 | Fluorescence imaging of endogenous CYP1A in living cells | |
| CYP1A1 | NBCeN Probe 3 [53] | 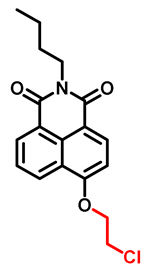 | 2.5 | 0.84 | 450/562 | Fluorescence imaging of endogenous CYP1A1 in living cells, TPM c images of a fresh rat liver slice |
| iPrBN Probe 4 [56] | 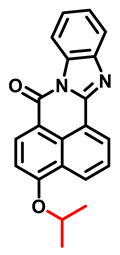 | 0.036 | 0.29 | 470/525 | TPM c imaging of CYP1A1 activities in living human cancer cells, tumor tissue slices, zebrafish | |
| DPCl Probe 5 [57] | 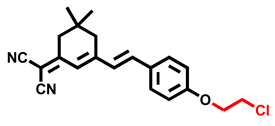 | 0.026 | / | 555/673 | Fluorescence imaging of endogenous CYP1A1 in living cells, zebrafish | |
| NEiPN Probe 6 [58] |  | 0.04874 | / | 374/446 | Fluorescence imaging of endogenous CYP1A1 in living cells, zebrafish | |
| CHPO Probe 7 [59] | 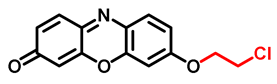 | 0.073 | 1.2 | 540/590 | Fluorescence imaging of endogenous CYP1A1 in living cells | |
| BCy-CYP Probe 8 [60] | 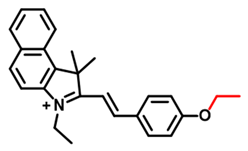 | 2.7 | / | 500/625 | Fluorescence imaging of endogenous CYP1A1 in living cells, effects of synergistic therapy of carnosol and cisplatin in the MCF-7 tumor-bearing mice | |
| CYP1A2 | HBMN Probe 9 [65] | 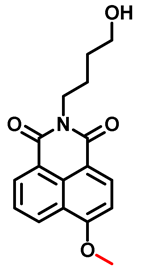 | 0.05 pg/mL−1 | 6.01 | 370/460 | Fluorescence imaging identifies CYP1A1 induced by different concentrations of TCDD d in living cells |
| CYP1B1 | 6b e Probe 10 [67] | 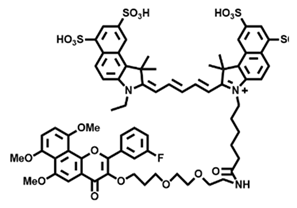 | / | / | 675/720 | Fluorescence imaging CYP1A1 in HCT-15 colon cancer cells, in vivo and in vitro NIR imaging for HCT-15 tumor-bearing mice |
| Bio-Marker | Name | Structure a | LOD/nM | Km/μM | Ex/Em b nm/nm | Biological Application |
|---|---|---|---|---|---|---|
| CYP2J2 | BnXPI Probe 11 [70] | 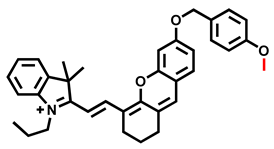 | 0.024 mg/mL | 4.2 | 656/718 | Bioimaging and graphical quantification of average fluorescence intensity of CYP2J2 activities in living human cancer cells, bioimaging of angiogenesis in vivo and ex vivo, in vivo NIR fluorescence imaging of CYP2J2 in tumor-bearing nude mice |
| DPBM Probe 12 [71] | 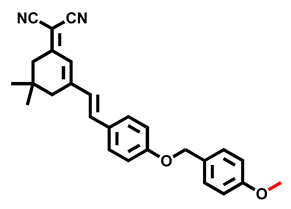 | 0.09 nM | / | 555/673 | Fluorescence imaging of endogenous CYP2J2 in living cells, BALB/c nude mice | |
| MXMB Probe 13 [72] | 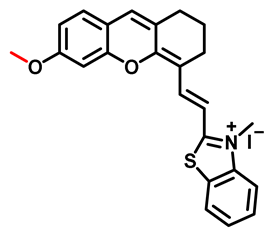 | / | 0.51 | 670/724 | Fluorescence bioimaging of CYP2J2 in vitro and in vivo, 2D and 3D confocal fluorescence imaging of tube formation | |
| CYP2C9 | DDAM Probe 14 [72] | 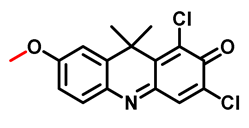 | / | 3.8 | 600/658 | Fluorescence imaging of endogenous CYP2C9 in living cells, inhibitory effects of various clinical drugs toward human CYP2C9 |
| CYP2D6 | McMe Probe 15 [78] |  | 0.052 nM | / | 400/555 | Fluorescence imaging of endogenous CYP2D6 in living cells, tumor-bearing mice |
| Bio-Marker | Name | Structure a | LOD/nM | Km/μM | Ex/Em b nm/nm | Biological Application |
|---|---|---|---|---|---|---|
| CYP3A4 | NEN Probe 16 [87] | 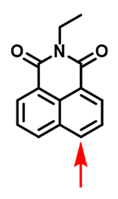 | / | 59.8 | 450/558 | TPM c fluorescence imaging of CYP3A4 in human primary hepatocytes, Fluorescence imaging of CYP3A4 in living zebrafish |
| CYP3A5 | Hcy-Br Probe 17 [91] | 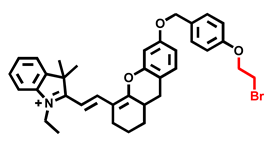 | 0.04903 nM | / | 600/690 | Fluorescence imaging of endogenous CYP1A1 in living cells, in vivo NIR fluorescence imaging of CYP3A5 in tumor-bearing nude mice |
| CYP3A4/ CYP3A5 | BN-1 Probe 18 [95] | 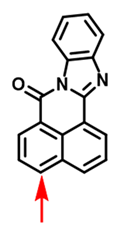 | / | 3.5/10.1 | 470/526 | TPM c imaging of CYP3A activity in the living specimen, High-throughput and visual characterization of mechanism-based CYP3A inactivation of herbal medicines |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Sheng, Y.; Li, W.; Yuan, L. Recent Advances in Molecular Fluorescent Probes for CYP450 Sensing and Imaging. Chemosensors 2022, 10, 304. https://doi.org/10.3390/chemosensors10080304
Li H, Sheng Y, Li W, Yuan L. Recent Advances in Molecular Fluorescent Probes for CYP450 Sensing and Imaging. Chemosensors. 2022; 10(8):304. https://doi.org/10.3390/chemosensors10080304
Chicago/Turabian StyleLi, Haiyan, Yang Sheng, Wei Li, and Lin Yuan. 2022. "Recent Advances in Molecular Fluorescent Probes for CYP450 Sensing and Imaging" Chemosensors 10, no. 8: 304. https://doi.org/10.3390/chemosensors10080304
APA StyleLi, H., Sheng, Y., Li, W., & Yuan, L. (2022). Recent Advances in Molecular Fluorescent Probes for CYP450 Sensing and Imaging. Chemosensors, 10(8), 304. https://doi.org/10.3390/chemosensors10080304