Abstract
Lyme disease and human babesiosis are the most common tick-borne zoonoses in the Temperate Zone of North America. The number of infected patients has continued to rise globally, and these zoonoses pose a major healthcare threat. This tick-host-pathogen study was conducted to test for infectious microbes associated with Lyme disease and human babesiosis in Canada. Using the flagellin (flaB) gene, three members of the Borrelia burgdorferi sensu lato (Bbsl) complex were detected, namely a Borrelia lanei-like spirochete, Borrelia burgdorferi sensu stricto (Bbss), and a distinct strain that may represent a separate Bbsl genospecies. This novel Bbsl strain was detected in a mouse tick, Ixodes muris, collected from a House Wren, Troglodytes aedon, in Quebec during the southward fall migration. The presence of Bbsl in bird-feeding larvae of I. muris suggests reservoir competency in three passerines (i.e., Common Yellowthroat, House Wren, Magnolia Warbler). Based on the 18S ribosomal RNA (rRNA) gene, three Babesia species (i.e., Babesia divergens-like, Babesia microti, Babesia odocoilei) were detected in field-collected ticks. Not only was B. odocoilei found in songbird-derived ticks, this piroplasm was apparent in adult questing blacklegged ticks, Ixodes scapularis, in southern Canada. By allowing live, engorged ticks to molt, we confirm the transstadial passage of Bbsl in I. muris and B. odocoilei in I. scapularis. Bbss and Babesia microti were detected concurrently in a groundhog tick, Ixodes cookei, in Western Ontario. In Alberta, a winter tick, Dermacentor albipictus, which was collected from a moose, Alces alces, tested positive for Bbss. Notably, a B. divergens-like piroplasm was detected in a rabbit tick, Haemaphysalis leporispalustris, collected from an eastern cottontail in southern Manitoba; this Babesia species is a first-time discovery in Canada. This rabbit tick was also co-infected with Borrelia lanei-like spirochetes, which constitutes a first in Canada. Overall, five ticks were concurrently infected with Babesia and Bbsl pathogens and, after the molt, could potentially co-infect humans. Notably, we provide the first authentic report of I. scapularis ticks co-infected with Bbsl and B. odocoilei in Canada. The full extent of infectious microorganisms transmitted to humans by ticks is not fully elucidated, and clinicians need to be aware of the complexity of these tick-transmitted enzootic agents on human health. Diagnosis and treatment must be administered by those with accredited medical training in tick-borne zoonosis.
1. Introduction
Lyme disease and human babesiosis are the most frequently reported tick-borne zoonoses in temperate North America [1], and have considerable economic, veterinary, and medical impact [2]. The length of attachment time of ticks and the presence of infectious microbes in human-biting ectoparasites often come into question at medical clinics and emergency departments. Delays in diagnosis and treatment become chronic infections. Based on US findings, approximately 63% of Lyme disease patients develop chronic Lyme disease [3]. With concurrent Lyme disease and human babesiosis, patients frequently have more pronounced symptoms and, in some cases, they can have fatal outcomes [4]. Certain areas in northeastern and north-central North America, such as the eastern part of Long Island, New York State, have endemic areas where 56% of the Lyme disease patients have coexisting human babesiosis [4].
Human babesiosis is a malaria-like zoonosis caused by microscopic parasites belonging to the genus Babesia [5]. This intraerythrocyte piroplasm (Apicomplexa: Piroplasmida: Babesiidae) is commonly carried and transmitted by hard-bodied ticks (Acari: Ixodidae), but has other modes of transmission. The world’s first described human case of babesiosis was a fatal case in an asplenic, male farmer in Croatia [6]. The clinical symptoms are broad-ranging with some patients being asymptomatic while others have a fulminant disease that can result in death. At least 100 Babesia species from around the world have been reported [7], and this apicomplexan pathogen infects multiple vertebrates, including humans.
Lyme disease is caused by members of the Borrelia burgdorferi sensu lato (Bbsl) complex, which consists of at least 23 genospecies, and is typically transmitted by ixodid ticks [8]. Bbsl is normally carried by ixodid ticks; however, this spirochete has other means of transmission. Bbsl is pleomorphic and has diverse forms, and can evade the immune response, and become persistent [9,10,11,12]. If this complex, multisystem zoonosis is not recognized and treated early, it can develop into chronic Lyme disease [12,13].
Each tick species has its own inherent range, hosts, and pathogens. Some ticks, such as the blacklegged tick, Ixodes scapularis, parasitize both birds and mammals, and have both a short- and long-distance range. Based on avian biodiversity, at least 82 species of birds are parasitized by larval and nymphal I. scapularis ticks. Songbirds (order Passeriformes) play an integral role in the wide dispersal of bird-feeding ticks and associated pathogens [14,15,16,17]. Not surprising, migratory passerine birds are able to transport ticks long distances during marathon flights to and from their wintering and breeding grounds each spring and fall [14,18,19,20,21,22]. Some neotropical and southern temperate passerines are known to transport bird-feeding ticks over 600 km/day [23,24,25,26]. Some of these songbird-transported ticks may originate from as far south as Brazil, and be imported into Canada during northward spring migration [27,28,29,30,31]. On the other hand, the groundhog tick, Ixodes cookei, which is not a bird-feeding tick, has a very localized home range on terrestrial mammals.
Ixodes scapularis may carry any combination of nine different polymicrobial pathogens with the potential to cause human and animal diseases [2]. Many etiological microbes are co-transmitted by I. scapularis ticks. As well, the American dog tick, Dermacentor variabilis, can harbour at least three different tick-borne, zoonotic pathogens [2].
Songbird-derived ticks include the blacklegged tick (I. scapularis), mouse tick (Ixodes muris), the rabbit tick (Haemaphysalis leporispalustris), the rabbit-associated tick (Ixodes dentatus). Each of these bird-feeding ticks carry tick-borne pathogens, and the infection prevalence of Bbsl ranges from 15% to 59% in I. scapularis nymphs during spring migration [17,20,21,27,28,29]. Whenever ground-frequenting passerines are heavily infested with ticks, they can initiate new foci of established populations hundreds of kilometres from their original geographic source [14,32].
Documentation of Bbsl-positive I. scapularis ticks within the southernmost part of mainland Ontario [33,34,35,36,37,38] have been ongoing. In contrast, documentation of Babesia-positive I. scapularis ticks have been limited [39,40]. It is noteworthy that Babesia odocoilei has been reported in I. scapularis ticks collected in Indiana, Maine, Massachusetts, Wisconsin [41] and, likewise, in Pennsylvania [42]. The latter account specifically reports a human as the host of a B. odocoilei-positive I. scapularis.
The primary objective of this study was to determine the presence of Babesia species and Bbsl genospecies in ticks collected from avian and mammalian hosts, and ascertain whether there are emerging tick-borne pathogens that have previously gone unnoticed in Canada.
2. Materials and Methods
2.1. Tick Collection
This study represents ixodid ticks collected in Canada during 2018, plus one special tick collected in 2017. Ticks were collected by bird banders, wildlife rehabilitators, road crew workers, Fatal Light Awareness Program staff [43], veterinarians, and the public in five interior Canadian provinces. Some of these ticks were also collected from humans and client-owned companion animals (i.e., feline, canine, equine); these hosts had no history of travel. Any live, fully engorged ticks were held to molt to the next developmental life stage or, in the case of a gravid female, to lay eggs.
Wild-caught ticks were collected from songbirds and mammals using fine-pointed, stainless steel forceps. Live ticks were put in a transparent 8.5 mL polypropylene tube (15.7 × 75 mm, round-bottomed) (Sarstedt, Montreal, Quebec, Canada). The top of the tube was covered with fine tulle netting (3 cm diameter) to allow ventilation for ixodid ticks. A polyethylene push cap with a 7 mm hole was placed into the top of the tube to secure the tulle netting, and prevent ticks from escaping. Each tube, which contained the ticks from one host, was placed in a double-zipped plastic bag with a slightly moistened paper towel to maintain high humidity. All ticks were sent to the lab for identification (J.D.S.). The Amblyomma nymph was tentatively identified using a taxonomic key [44] and, following the nymph–adult molt, Amblyomma taxonomic keys for adults indigenous to the Western Hemisphere were used [45,46]. Likewise, for Ixodes ticks, a larval key [47], a nymphal key [48], and an adult key [49] were used. Ixodes species were exposed to a long-day photoperiod of 16:8 (L:D) h, while Amblyomma ticks from the Neotropics were held at a photoperiod of 12L:12D h daily. Complete records (i.e., geographical location, tick collection date, tick species, developmental life stage, degree of engorgement, host species) were logged for each tick collection. To preserve ticks, they were stored in 2 mL microtubes containing 95% ethyl alcohol.
Adult questing ticks were collected from low-lying vegetation by flagging. The flagging cloth (60 × 70 cm) was made of flannel-backed vinyl, and the aluminum, telescopic pole was 195 cm.
2.2. Bacteria and Piroplasm Detection
Ticks that were stored in 95% ethyl alcohol (ETOH) were initially rinsed in fresh absolute ETOH, and air dried. Each tick was then macerated with a separate, sterile scalpel blade that was first rinsed in 1% sodium hypochlorite followed by two rinses with 70% ETOH. A different scalpel blade was used for each tick. DNA was then extracted from tick tissues using a commercial kit (GeneJET Genomic DNA Purification Kit, ThermoFisher Scientific, Waltham, MA, USA) using the manufacturer’s protocol for tissues. Final elution consisted of 100 µL of TE buffer. PCR testing for pathogen DNA utilized 2.5 µL of eluted DNA sample as the initial template. Each procedural round of 10−12 tick DNA extractions included two negative control extractions with no template, and these extracts were tested along with tick template to ensure no DNA artifact contamination of extraction reagents during the DNA extraction process.
Tick DNA extracts were screened for the presence of Bbsl DNA using a nested PCR that amplifies a portion of the flagellin (flaB) gene of Bbsl, with slight variations from a previously described protocol [50]. The primary PCR assay, which targets a 497 nt fragment of the flaB gene, used the following primers, 271F: 5′-AAG-GAA-TTG-GCA-GTT-CAA-TCA-GG-3′ and 767R: 5′-GCA-TTT-TCT-ATT-TTA-GCA-AGT-GAT-G-3′. The secondary (nested) PCR employed 1 µL of primary amplification product as template with primers that amplify a 437 nt internal fragment, 301F: 5′-ACA-TAT-TCA-GAT-GCA-GAC-AGA-GG-3′ and 737R: 5′-GCA-TCA-ACT-GTA-GTT-GTA-ACA-TTA-ACA-GG-3′.
For Babesia testing and DNA sequencing of ticks, the 18S ribosomal RNA (rRNA) gene primer was applied, and the same protocol was used as previously described by Casati et al. [51]. Along with negative control extraction samples, sterile water was used as additional controls in PCR testing to confirm that PCR reagents were free of DNA artifact contamination.
2.3. DNA Sequence Analysis
PCR products from the Babesia 18S rDNA and the Bbsl flaB positive samples were purified using the Wizard® SV Gel and PCR Clean-Up System (Promega, Madison, WI, USA). DNA templates were sequenced [52] using both the forward and reverse primers. Investigator-derived sequences were aligned using ClustalX [53], and submitted to BLAST (Basic Local Alignment Search Tool) comparison to determine similarity with archived sequences in the GenBank database [54]. A subset of sequences from DNA amplicons representing different tick-host-pathogen associations were accessioned in GenBank.
3. Results
3.1. Tick Collection
This study consists of 311 ixodid ticks from 2018, plus one novel tick from 2017. Specifically, for 2018, we had seven tick species belonging to four genera (Amblyomma, Dermacentor, Haemaphysalis, and Ixodes) collected in five interior provinces (Alberta, n = 16; Manitoba, n = 6; Ontario, n = 229; Quebec, n = 58; and Saskatchewan, n = 2) (Figure 1). Taken as a whole, these ticks consisted of seven species (i.e., Amblyomma inornatum, n = 1; Dermacentor albipictus, n = 16; D. variabilis, n = 88; H. leporispalustris, n = 33; I. cookei, n = 2; I. muris, n = 16; and I. scapularis, n = 155) (Table 1). All ticks collected from mammals had no history of travel.
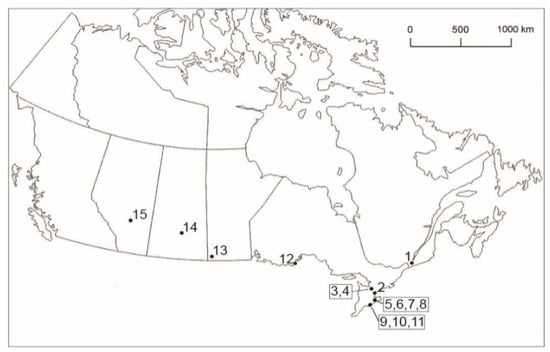
Figure 1.
Geographic locations of sites in Canada where ixodid ticks were collected from avian and mammalian hosts, and by flagging. (1) Ste-Anne-de-Bellevue, Quebec, 45.40° N, 73.95° W; (2) Toronto, Ontario (Fatal Light Awareness Program), 43.74° N, 79.37° W; (3) Barrie, Ontario, 44.39° N, 79.69° W; (4) Elmvale, Ontario, 44.58° N, 79.87° W; (5) Ruthven Park, Ontario (Cayuga), 42.97° N, 79.87° W; (6) Dunnville, Ontario, Property #1 (NT), 42.91° N, 79.61° W; (7) Dunnville, Ontario, Property #1 (NR), 42.90° N, 79.62° W; (8) Dunnville, Ontario, Property #2 (NR), 42.90° N, 79.63° W; (9) Turkey Point Provincial Park, Ontario, 42.70° N, 80.33° W; (10) Turkey Point, Ontario, former Charlotteville landfill, 42.71° N, 80.33° W; (11) Long Point, Ontario, 42.52° N, 80.17° W; (12) McKellar Island, Ontario (Thunder Bay), 48.19° N, 89.13° W; (13) Melita, Manitoba, 49.27° N, 100.99° W; (14) Manitou District and Regional Park, Saskatchewan, 51.68° N, 105.68° W; and (15) Pine Lake, Alberta, 52.11° N, 113.48° W. The locations in parentheses represent mailing addresses.

Table 1.
Presence of Borrelia burgdorferi sensu lato and Babesia spp. in ticks collected from avian and mammalian hosts in five interior provinces in Canada, 2018.
Overall, 174 questing adult ticks (D. variabilis, I. scapularis) were collected by flagging low-level vegetation in southwestern Ontario. At each of the five sites (6,7,8,9,10), D. variabilis and I. scapularis are sympatric.
Of 16 bird species captured, the Common Yellowthroat, a neotropical species, was most frequently parasitized by bird-feeding ticks (Table 1). Two songbirds had co-infestations of two different tick species. Specifically, an I. scapularis nymph and an I. muris nymph were co-feeding on a Common Yellowthroat at Ste-Anne-de-Bellevue, Quebec (Site 1) on 14 August 2018. Additionally, an Amblyomma inornatum nymph and an I. scapularis nymph concurrently parasitized a Veery at Ruthven Park, Ontario (Site 5) on 16 May 2018 [39].
3.2. Pathogen Detection
All 2018 ticks were tested for Babesia species and B. burgdorferi sensu lato. Table 2 and Table 3 list select ticks that were positive for Babesia spp. and Bbsl genospecies. In one Lyme disease endemic area in the Region of Haldimand-Norfolk (Site 10), 11 (34%) of 32 I. scapularis adults were positive for Bbsl; ticks in this established population were also infected with B. odocoilei. In the eastern part of the Region of Haldimand-Norfolk (Site 6), three (37%) of eight questing blacklegged tick adults were positive for Bbsl; likewise, the ticks in this breeding colony contain B. odocoilei. A total of five co-infections of Bbsl and Babesia were detected in ticks (Table 2 and Table 3). These two tables have select representations of ticks with Bbsl and/or Babesia amplicons that have been submitted to GenBank. Certain Bbsl amplicons were not included in Table 2 and Table 3 because we were unable to obtain clean sequence data. Four I. muris larvae were collected from a Magnolia Warbler at Ste-Anne-de Bellevue, Quebec on 18 August 2018, and three of these larvae molted to nymphs; a single larva was positive for Bbsl. This microbial detection suggests that Magnolia Warbler may be a reservoir-competent host. Significantly, this novel collection also provides the first record of enzootic transfer (larva to nymph) of Bbsl in I. muris.
Table 2.
Select tick-host-Babesia associations with corresponding DNA sequences, Canada, 2018.
Table 3.
Select tick-host-pathogen interactions for ticks infected with Borrelia burgdorferi sensu lato collected from birds and mammals, Canada, 2017 and 2018.
In 2017, an I. muris larva collected from a House Wren on 27 August 2017 at Site 1 harboured a unique Bbsl strain. The 367 nt flagellin (flaB) gene sequence that we obtained was 100% identical with that of the Bbsl strain W97F51 (GenBank AY884355) from Wisconsin; the next most similar Bbsl species flaB strains included reference B. lanei strains that shared 362/367 (99%) similarity.
This laboratory (K.L.C.) has never contained any reference strain cultures of W97F51 or Borrelia lanei. Since this laboratory has never detected another strain identical to B. lanei or the W97F51 strain from any source prior to the detection of the unique Bbsl strain in an I. muris larva collected in Canada, it is highly unlikely that this Bbsl finding is the result of any type of PCR error or DNA artifact contamination.
3.2.1. Detection in Bird-derived Ticks
Overall, in 2018, five passerine birds were infested with Babesia-positive I. scapularis nymphs, and six birds were parasitized by Bbsl-infected larvae and nymphs.
Two single I. scapularis nymphs were collected from two individual Gray Catbirds at Site 5 on 24 May 2018. Each of these nymphs was infected with B. odocoilei piroplasms (Figure 2).

Figure 2.
Gray Catbird parasitized by an I. scapularis nymph at Site 5. This nymph was infected with Babesia odocoilei. The white arrow points to the location of an engorged tick (the same below). Photo: Caleb Scholtens.
On 26 May 2018, a fully engorged I. scapularis nymph was collected from a Lincoln’s Sparrow at Site 11; this nymph molted to a female in 39 days, and was infected with B. odocoilei. As well, two I. scapularis nymphs parasitized a Common Yellowthroat at Site 1 on 19 May 2018, and both of these nymphs were infected with Bbss (Table 3).
The GenBank accession numbers (i.e., MK620851 {Bbsl}; MK628544 {Babesia odocoilei}), which pertain to a co-infection of Bbsl and Babesia odocoilei in an I. scapularis nymph parasitizing a Veery [39], were previously published (Table 2 and Table 3). This Veery was concurrently infested by an Amblyomma inornatum nymph and an I. scapularis nymph.
3.2.2. Detection in Mammal-related Ticks
A fully engorged I. cookei nymph was collected from a cat with outdoor exposure on 25 October 2018 (Site 3). This I. cookei tick was co-infected with B. microti and Bbsl [Table 2 and Table 3].
In the present study, two (29%) of the seven I. scapularis females feeding on dogs were positive for Bbsl.
A fully engorged I. scapularis female was collected from a riding horse on 5 November 2018 (Site 4), and this tick tested positive for Bbsl.
In central Canada, a H. leporispalustris (rabbit tick) female was collected from an eastern cottontail on 16 June 2018 (Site 13). This tick was co-infected with a Babesia divergens-like piroplasm and, also, a Borrelia lanei-like spirochetal bacterium.
Notably, 16 winter ticks, D. albipictus, were collected from a moose, Alces alces, on 22 April 2018 (Site 15). A single D. albipictus female tested positive for Bbss: However, none tested positive for Babesia.
None of the I. muris ticks was positive for Babesia spp.
3.2.3. Detection in Questing Ticks
Questing adult I. scapularis (n = 93) were collected by flagging from five sites (i.e., 6, 7, 8, 9, 10) in Haldimand-Norfolk, and the blended infection prevalences were: Bbsl: 24/93 (26%) and Babesia odocoilei: 4/93 (4%). We provide the first account of an I. scapularis tick (female) co-infected with B. odocoilei and Bbsl; it was collected by flagging on 10 May 2018 at Turkey Point Provincial Park (Site 9). Similarly, an I. scapularis female was collected by flagging from low-level vegetation (Site 6) on 25 May 2018, and it was co-infected with B. odocoilei and Bbsl.
In all, 88 adults of the American dog tick, Dermacentor variablis, were collected; seven were removed from humans and 81 collected by flagging. None of the D. variabilis was positive for Babesia spp. or Bbsl.
4. Discussion
In this tick-host-microbe study, we announce the detection of three important Babesia species and three diverse Bbsl genospecies or strains in Canada. The occurrence of Babesia piroplasms in three indigenous tick species (i.e., H. leporispalustris, I. cookei, and I. scapularis) grants substantive proof that these piroplasms are present in the environment. Perhaps most significantly, three Babesia species (i.e., B. divergens-like, B. microti, and B. odocoilei) piroplasms were present in these ixodid ectoparasites. Not only are small and large mammals implicated in the short-distance dissemination of ticks, songbirds are involved in the long-distance dispersal of avian-transported ticks. Furthermore, we verify the presence of three Borrelia groups (i.e., a novel Bbsl strain, B. burgdorferi sensu stricto, and another strain most similar to Borrelia lanei) in Canada. In fact, the flaB gene sequence of the latter Bbsl strain was actually identical to the W97F51 Wisconsin strain [55]. Based on analysis of several different genes, Caporale et al. found that W97F51 to be most similar to Borrelia bissettiae strains [55]. They posit this borrelial microbe might be a unique Bbsl species, but even they did not fully assess that possibility. Due to a shortage of DNA, we did not perform extensive multi-locus sequence typing (MLST) or multi-locus sequence analysis (MLSA). Therefore, we have simply referred to this special Bbsl strain as another unique Bbsl strain. Notably, our DNA findings do not prove reservoir competence of hosts or vector competence of ticks. However, by letting live, engorged ticks molt to the next life stage, we were able to affirm transstadial passage of Bbsl in I. muris and B. odocoilei in I. scapularis. In addition, we have neither proved that hosts are infected nor ticks are competent vectors. Our findings show a diversity of tick-borne, zoonotic pathogens in Canada, and certain pathogens present a public health risk.
4.1. Babesia Species in Ticks
In all, nine B. odocoilei PCR amplicons were detected. These apicomplexan amplicons were all associated with I. scapularis ticks (i.e., questing adults, four; bird-derived nymphs, five). Since cervine hosts (i.e., white-tailed deer, Odocoileus virginianus) are reservoirs of B. odocoilei, blacklegged ticks feeding on infected deer can acquire Babesia infection and, following the molt, can subsequently be an enzootic bridge to humans. Certain Babesia spp. (e.g., B. divergens and Babesia sp. EU1) invade the female ticks’ ovaries, and are transmitted transovarially to the next generation [2,56], whereas other Babesia sp. (e.g., B. microti) are not passed via the eggs [2,57]. Enzootically, transovarial transmission (female to eggs) of B. odocoilei takes place in I. scapularis females. Once the eggs are infected, transstadial passage (egg to larva or larva to nymph or nymph to adult) occurs [2]. When B. odocoilei-infected ticks feed on a suitable host, they can promptly transmit babesial sporozoites because the ticks’ salivary glands are infected [58]. These enzootic modes of transmission provide a natural enzootic pathway to perpetuate Babesia in blacklegged ticks, and facilitate transmission to humans during a tick bite. This deer-tick-deer, enzootic cycle of B. odocoilei contributes to the perpetual maintenance, and the dissemination of this piroplasm. Consistent with other researchers [58], we demonstrate in southwestern Ontario that the biogeographic distribution of B. odocoilei coincides with the dispersal of I. scapularis.
4.1.1. Ticks Collected from Songbirds
In the present study, B. odocoilei-positive I. scapularis ticks were collected from five ground-frequenting songbirds (House Wren, Veery, Gray Catbirds (n = 2), Lincoln’s Sparrow) during peak spring migration. Remarkably, two Gray Catbirds were parasitized by B. odocoilei-infected nymphs; both bird parasitisms occurred on the same day and the same location (Site 5). These bird parasitisms are the first report of B. odocoilei-infected ticks on Gray Catbirds (Figure 2). If a human was bitten by either of these B. odocoilei-infected nymphs, it is possible that they could acquire this piroplasm. These collections provide evidence that an endemic area of B. odocoilei may be present in the nearby environs. Since the wild-caught ticks on these passerines are nymphs, we are not able to differentiate whether B. odocoilei was acquired directly from the host birds or derived earlier when I. scapularis larvae parasitized an B. odocoilei-infected host.
Scott et al. published the first report of B. odocoilei in an I. scapularis tick (nymph) collected from a bird (Veery) [39]. Subsequently, Milnes et al. reported B. odocoilei-positive pools of I. scapularis larvae collected from two songbirds [40]. However, there is a paucity of information on how these I. scapularis larvae became infected with B. odocoilei.
Since B. odocoilei is in the same sister clade as other pathogenic Babesia strains (i.e., Babesia sp. EU1; Babesia divergens; Babesia divergens-like species) [6,59,60,61,62], it is possible that B. odocoilei might also be pathogenic to people, especially patients who are concurrently infected with tick-borne, zoonotic pathogens, and are immunologically hampered by these infections.
At Site 11, an I. scapularis nymph was collected from a Lincoln’s Sparrow; this bird parasitism constitutes the first account of a B. odocoilei-positive tick parasitizing a Lincoln’s Sparrow. We held this tick to molt, and during the 39-day transstadial passage, B. odocoilei successfully cleared the nymph–adult molt. This babesial detection provides the first authentic confirmation of transstadial passage of B. odocoilei in I. scapularis. Therefore, unfed I. scapularis larva, nymphs, and females can bite people, and potentially infect them with B. odocoilei.
Two I. scapularis nymphs were collected from a Common Yellowthroat at Site 1, and both of these nymphs were infected with Bbsl. This co-infestation suggests that this bird was spirochetemic with Bbsl. Co-infestations of bird-feeding ticks are frequent when northward-migrating passerines make stopovers at Lyme disease endemic areas en route to breeding grounds or later while these birds are nesting in a Lyme disease endemic area.
4.1.2. Ticks Derived from Mammals
In Saskatchewan, B. odocoilei has been detected in elk (Cervus elaphus canadensis) that had chronic weight loss and unthriftiness and, in the same herd, had sudden deaths [63] Any Babesia-positive ticks collected from mammals were all co-infections, and are addressed under Section 4.3.2.
4.1.3. Questing Ticks
During flagging operations, we collected four field-collected I. scapularis adults that were positive for B. odocoilei. These B. odocoilei-positive, I. scapularis adults were collected in established populations (Sites 6, 9, 10) of I. scapularis ticks on mainland Ontario. Questing ticks are important in this study because they pinpoint the primary vector of B. odocoilei and, also, substantiate transstadial passage of this piroplasm.
4.2. Borrelia burgdorferi Sensu Lato in Ticks
4.2.1. Ticks on Wild-caught Birds
Of special significance, we present the first documentation of a potentially unique Bbsl strain in Canada. This de novo Bbsl strain (GenBank accession number MH290738) was detected in an I. muris larva that was collected from a House Wren (Table 3), and is the first account of this Bbsl strain in this tick species in Canada (Figure 3). Using a portion of the flaB gene, this Bbsl strain is a 100% match to a Wisconsin strain W97F51 obtained in 1997 [55]. Moreover, the flaB fragment sequence is ~99% identical to Borrelia lanei reference strains. This de novo Bbsl strain may possibly represent a distinct and different Bbsl genospecies. Thus, we are simply calling this novel strain B. burgdorferi sensu lato. Moreover, since this I. muris larva was collected during southbound fall migration, this bird parasitism suggests that this unique Bbsl strain may be established in Canada.

Figure 3.
House Wren parasitized by Ixodes scapularis nymphs. While ground-dwelling passerines are foraging for morsels on the forest floor or meadow, they can be parasitized by bird-feeding ticks. Photo: Simon Duval.
Other researchers have previously reported I. muris larvae parasitizing songbirds [17,21,22,28], but this is the first report of a Bbsl-infected I. muris larva parasitizing a bird. The presence of a Bbsl-positive I. muris larva parasitizing the House Wren suggests that this bird species has reservoir competency.
Connecticut researchers have cultured Bbsl from the blood of Common Yellowthroat, Gray Catbird, and American Robin [64]. Moreover, they have isolated Bbsl from I. scapularis larvae collected from songbirds (i.e., Gray Catbird, Brown-headed Cowbird, Field Sparrow, and Common Yellowthroat), and suggest that these ground-foraging songbirds are reservoir-competent hosts [64]. Using spirochete-free, xenodiagnostic larvae, Richter et al. determined that the American Robin is, indeed, a competent reservoir for Bbsl [65]. Since transovarial transmission of Bbsl is not present in wild-caught I. scapularis [66], we extrapolate that I. muris larvae may also acquire Bbsl directly from spirochetemic songbirds.
In the present study, Bbsl-infected I. muris larvae were collected from a Magnolia Warbler and a Common Yellowthroat during southward fall migration, and these novel bird parasitisms suggest that these passerines have reservoir competency. These enzootic results suggest that both the Magnolia Warbler and the Common Yellowthroat were spirochetemic and, during the blood meal, Bbsl was transmitted to these attached larvae. Since these juvenile birds have just fledged the nest, and had scant exposure to ticks, it is possible that the mother birds were spirochetemic, and may have transmitted Bbsl to their offspring. In addition, two I. muris nymphs were collected from a juvenile Common Yellowthroat during southward fall migration, and one of these co-feeding nymphs tested positive for Bbsl (Figure 4). Of epidemiological significance, I. muris is a Lyme disease vector tick that has vector competence for Bbsl, and can transmit Lyme spirochetes to humans [22].

Figure 4.
Common Yellowthroat, adult male, parasitized by nymphal Ixodes scapularis ticks. Since these nymphs were collected during the nesting and fledgling period, this bird parasitism indicates that this location has an established tick population. Photo: Ana Morales.
Bi-directional migration of neotropical and southern temperate songbirds is a natural part of phenology, and wide dispersal of songbird-transported ticks is an ongoing phenomenon. Spring migration of passerine migrants coincides with the peak questing period of I. scapularis nymphs in May and early June [67]. During spring migration, neotropical and southern temperate songbirds, such as the Common Yellowthroat, facilitate the long-distance dispersal of ticks (Figure 4). Passerine migrants transport I. scapularis larvae and nymphs into Canada annually [17,18,20,21,27,28,31], and annual cross-border avian flight provides a perpetual source of pathogen-laden ticks from southern latitudes.
Although we did not sample gallinaceous birds, such as Wild Turkeys (Meleagris galopavo) and Ring-necked Pheasants (Phasianus colchicus), which are native in the Carolinian forest region, we realize that these land-based avifauna do play an important role in the enzootic transmission cycle of Bbsl [68].
During the nesting and fledgling period, ground-foraging passerines are short-distance disseminators of locally acquired ticks. In particular, juvenile (hatch-year) songbirds, which fly south for the winter, have not yet migrated. During this early summer period, a heavily infested juvenile songbird clearly shows that there is an established population of ticks within the nesting area (Figure 5).

Figure 5.
Song Sparrow, a juvenile, parasitized by three Ixodes scapularis nymphs (two are not visible). Since these ticks were acquired in close proximity to the nest, this bird parasitism indicates that an established population of I. scapularis is present within this nesting area. Photo: Ana Morales.
4.2.2. Ticks on Terrestrial Mammals
The predominant borrelial species in this study was Borrelia burgdorferi sensu stricto which is pathogenic to domestic animals (i.e., cats, dogs, horses) and to humans.
In the present study, seven dogs were parasitized by ticks (Table 1), and two dogs had ticks positive for Bbsl. In dogs, symptoms include polyarthritis, stiffness, sore paws, chewing of paws, fatigue, lethargy, depression, anorexia, and reluctance to walk and play [69]. In spite of standard antibiotic treatment, Bbsl can be persistent [70].
We provide the first report of a Bbsl-infected I. scapularis tick parasitizing a horse in Canada (Table 1). Although it was not possible to do a follow-up on this horse, Bbsl causes Lyme disease in horses [71]. The clinical symptoms of Lyme disease in horses include lameness, stiffness, neuroborreliosis, uveitis, and cutaneous pseudolymphoma [71]. Congenital Lyme disease may occur in mares and foals, especially in Lyme disease endemic areas [72]. Cats as mammalian hosts are described under the co-infection section (Section 4.3.2).
The occurrence of a winter tick, D. albipictus, which was infected with Bbsl, is a first-time discovery in western Canada. This Bbsl-positive, D. albipictus female was one of 16 D. albipictus adults collected from a moose, the largest member of the deer family. In Northwestern Ontario a Bbsl-positive D. albipictus was previously collected from an untraveled dog at Kenora, Ontario [73].
Terrestrial mammals provide short-distance dispersal of ticks, and maintain the enzootic transmission cycle of Bbsl within a Lyme disease endemic area. Ticks have an innate ability to avoid premature dislodgement from their hosts. They select secluded attachment sites (e.g., inside ear lobe) that are not subject to grooming or preening (Figure 6). In order to thwart tick dislodgement, ticks will attach beyond the reach of the incisors and the front paws or toes.
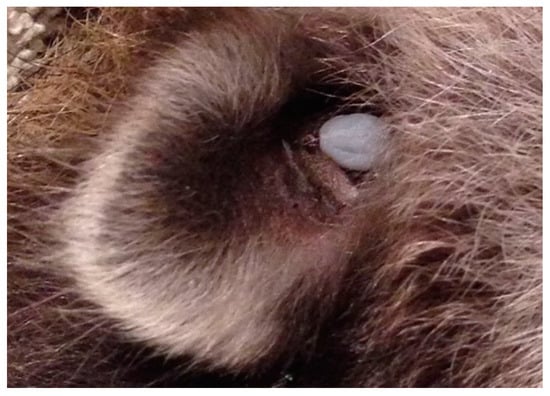
Figure 6.
Engorged Ixodes female parasitizing a medium-sized mammal inside its ear. Ticks select secluded areas of the body to prevent dislodgement during grooming and preening by front paws or incisors. Photo: Christina Carrieres, Wild ARC.
4.2.3. Questing Ticks
During flagging operations, we obtained 21 I. scapularis adults that were positive for Bbsl. These Bbsl-positive I. scapularis are congruent with other tick studies in southwestern Ontario [19,33,34,35,38]. When blacklegged ticks are not conducting host-seeking activities, they descend to the forest floor refuge, re-hydrate, and have a climate-controlled microhabitat. All life stages of blacklegged ticks reside in the cool, moist leaf litter, and are not subject to climate change. Since blacklegged ticks have antifreeze-like compounds (glycoproteins) in their bodies [74], this tick species can survive a significant temperature differential of 80 °C (−44 °C to +36 °C) at Kenora, Ontario [75,76]. When it comes to blacklegged ticks, climate change is a trivial issue [75,76].
4.3. Babesia and Borrelia burgdorferi Sensu Lato Co-infections in Ticks
In this study, we encountered five co-infections in ticks (Table 2 and Table 3). Co-infections were detected in three tick species (H. leporispalustris, I. cookei, I. scapularis) involving three vertebrate hosts (i.e., eastern cottontail, domestic cat, and Veery), respectively. These zoonotic microorganisms comprise: A) spirochetes: Borrelia lanei-like spirochete, Borrelia burgdorferi sensu stricto, and an unique Bbsl strain and B) piroplasms: Babesia divergens-like, Babesia microti, and Babesia odocoilei.
4.3.1. Co-infected Ticks on Birds
During spring and fall migrations, ground-foraging migrants make stopovers at select meadows and sylvatic areas to consume seeds, berries, and invertebrates. These energy-laden morsels include spent gravid I. scapularis females that have laid eggs, and have died. These tick habitats are also commonly inhabited with small mammals (i.e., deer mice, meadow voles, eastern chipmunk, shrews) that act as hosts for immature life stages of blacklegged ticks and I. muris ticks [14,48,61,77]. Several researchers indicate that I. scapularis are directly connected to B. odocoilei [39,40,41,78], and denote that B. odocoilei overlaps with the distribution range of I. scapularis and white-tailed deer. Meadows and wooded areas are community-centered foci where deer, small mammals, ground-dwelling songbirds congregate, and form enzootic hubs for the enzootic transmission cycle of Bbsl and B. odocoilei. Within these tick-conducive habitats, I. scapularis ticks and white-tailed deer play a pivotal role in perpetuating B. odocoilei.
A heavily infested songbird can initiate an established population of blacklegged ticks [32]. Whenever juvenile songbirds are infested with I. scapularis ticks, these tick collections clearly indicate that an established population is present. For example, ground-frequenting songbirds, such as the Rose-breasted Grosbeak, provide short-distance dispersal of ticks during the nesting and fledgling period (Figure 7).

Figure 7.
Rose-breasted Grosbeak, adult male, parasitized by Ixodes scapularis nymphs. Since this bird parasitism occurred during the nesting and fledgling period, these attached nymphs denote an established tick population in this locale. Photo: Ana Morales.
4.3.2. Co-infected Ticks on Terrestrial Mammals
The co-infection of B. microti and Bbsl in an I. cookei nymph collected from a cat at Site 3 is a first-time event. Not only is B. microti reported for the first time in I. cookei, it is the initial documentation of B. microti in Western Ontario. Of note, these two zoonotic pathogens are typically reported in blacklegged ticks [79], but not in I. cookei. Importantly, I. cookei bites humans [22,73,80,81,82], and this present study signifies that this cat-derived I. cookei could have simultaneously transmit these two tick-borne, zoonotic pathogens (e.g., B. microti and Bbsl) to companion animals or people [2,4,69,83]. Often domestic cats will have a subclinical Bbsl infection; however, they may have various symptoms including lethargy, lameness, irregular gait, pain on manipulation of hips and tail (hip and/or tail pain). They may also be subdued, depressed, and have inappetence (lack of desire or appetite), and/or have severe ataxia of hind legs [83].
The B. microti sequence detected in a cat-derived I. cookei nymph matches closely with a B. microti amplicon (GenBank accession number AF5446902) from a skunk in Massachusetts. Based on phylogenetic analysis, this strain is a carnivore-associated B. microti, and not a rodent-associated B. microti strain [84]. Even though Barrie, Ontario is 690 km from Massachusetts, the two related B. microti strains are congruent with each other. Not only are there carnivore- and rodent-associate strains, there are several raccoon-associated strains [84]. Although B. microti is widely reported in blacklegged ticks in the USA, it was previously not reported in I. cookei in Canada. Most notably, B. microti is reported in I. cookei which suggests that this piroplasm is cycling enzootically with groundhogs (woodchucks), Marmota monax. Ecologically, B. microti has been isolated from white-footed mice (Peromyscus leucopus) captured in Connecticut [85]. All three motile life stages of I. cookei feed on groundhogs, and are likely a reservoir host of B. microti. After the nymph–adult molt, this female could have transmitted Bbsl and B. microti to a human. Not only do I. cookei ticks carry and transmit deer tick virus (Powassan group virus) [86], they also harbour Babesia microti and Bbsl. Since I. cookei is a human-biting tick, it can act as an ecological bridge for B. microti between reservoir hosts (i.e., groundhogs, coyotes, skunks, raccoons) to humans and, therefore, this tick species is of epidemiological significance [80,82].
In North America, B. odocoilei is commonly associated with I. scapularis ticks [57] and, also, white-tailed deer [57,78]. White-tailed deer are hosts of all three motile life stages (larvae, nymphs, adults) of I. scapularis, and support the reproduction of I. scapularis. In contrast to Bbsl spirochetes, I. scapularis and cervine hosts both facilitate the enzootic transmission cycle of B. odocoilei. White-tailed deer are reservoir hosts of B. odocoilei; however, they are refractory to Lyme disease spirochetes [87].
In southern Manitoba, we report a H. leporispalustris tick infected with both a B. divergens-like piroplasm and, also, a Borrelia lanei-like spirochete (Table 2 and Table 3). This discovery marks the first report of a Babesia divergens-like piroplasm in Canada. Although H. leporispalustris ticks rarely bite humans [88], this tick species can transmit this piroplasm to lagomorphs and domestic animals, such as cats and dogs. Banerjee et al. documented Bbsl in H. leporispalustris ticks that were collected from a snowshoe hare (Lepus americanus) in northern Alberta [89]. In addition, Scott et al. reported Bbsl in H. leporispalustris collected from songbirds [22]. Reports of human cases with high levels of parasitemia caused by B. divergens-like microorganisms include residents of Missouri, Kentucky, Washington, Arkansas, Massachusetts, and Michigan [90]. In the latter case, Herc et al. reported an asplenic Michigan patient infected with a B. divergens-like/MO-1 piroplasm, and this 60-year-old lady experienced fatigue, nausea, and hemolytic febrile symptoms [90]. Not only have B. divergens-like infections been identified in the blood and spleen of eastern cottontail rabbits, they have also been detected in rabbit-associated ticks, I. dentatus, on Nantucket Island, Massachusetts, USA [91]. Both immature stages of I. dentatus and H. leporispalustris feed on migratory birds, and facilitate the wide dispersal of infected ticks across North America. Based on DNA sequence assessment, B. odocoilei and B. divergens-like piroplasms are closely related to B. divergens in the Babesia sensu stricto clade. In Europe, B. divergens is noted as the most common cause of human babesiosis, and can be fatal [6,57].
Both B. divergens-like species and B. lanei-like strains have a direct connection to lagomorphs. In fact, B. lanei (formerly Borrelia genomospecies 2) was detected in Ixodes spinipalpis and Ixodes pacificus (western blacklegged tick) ticks collected from eastern cottontails (Sylvilagus floridanus) and snowshoe hares, respectively, in southwestern British Columbia [92]. Since H. leporispalustris larvae and nymphs parasitize migratory songbirds, B. lanei-like spirochetes and B. divergens-like piroplasms could have been transported by songbird-transported ticks across the US-Canada border during northbound migratory flights. Biogeographically, the B. lanei-like spirochete is documented for the first time in Canada east of the Rocky Mountains.
4.3.3. Co-infected Questing Ticks
Of epidemiological significance, two I. scapularis females harboured co-existent Babesia and Bbsl (Table 1 and Table 2). If a person was bitten by either of these ticks, they could become concurrently infected by these potentially pathogenic microorganisms. A host-seeking I. scapularis female was collected by flagging at Turkey Point Provincial Park (Site 9), and this tick was co-infected with B. odocoilei and Bbsl. Similarly, an I. scapularis female was concurrently infected with B. odocoilei and Bbsl collected in the eastern part of Region of Haldimand-Norfolk (Site 6). If a companion animal or person had been bitten by either of these unfed females, it is theoretically possible that they could become infected with both B. odocoilei and Bbsl.
None of the adult D. variabilis was positive for B. odocoilei or Bbsl, which indicates that this tick species is neither a Lyme disease vector tick nor a vector of B. odocoilei. However, the American dog tick is known to harbour at least three different tick-borne, zoonotic pathogens, and an engorged female can cause tick paralysis [93].
4.4. Impact of Babesia and Bbsl on Humans
Canadian patients are testing positive for Lyme disease and human babesiosis [22,94]. Patients with these zoonoses often exhibit unusual symptoms, such as summer flu, and clinicians have trouble diagnosing these tick-borne diseases accurately. Pathologically, these co-infections typically cause greater disease severity, and have longer duration than either pathogens alone [95,96,97,98,99]. During a tick bite, these polymicrobial infections may be co-transmitted to their hosts. Symptoms from co-infections are typically more severe, and harder to treat with antimicrobials. In some coexisting Lyme disease and human babesiosis cases, patients die [4,62,98,99,100].
Babesiosis is a potentially life-threatening, zoonotic infection that can affect a variety of vertebrates, including cats, dogs, horses, cattle, and humans [2,42]. Pathologically, this piroplasm lives and multiples in erythrocytes, and is typically transmitted by ixodid ticks. Alternately, this intraerythrocytic hemoparasite can also be transmitted by blood transfusion [101,102,103,104] and transplacental passage [105,106,107,108]. When sporozoites invade red blood cells, symptoms range from a silent, subclinical infection to a fulminant, malaria-like disease that can result in death [6,57,60,61,62,95,96]. Some of the more common symptoms include sweats (particularly night sweats), chills, profound fatigue, malaise, weakness, increased thirst, fever, body aches, thrombocytopenia (decreased blood platelets), and a sense of ‘air hunger,’ especially those who are immunocompromised (i.e., 55 years and up; splenectomized; infected with two or more zoonotic pathogens) [95,96,97]. Once established in the human body, this babesial piroplasm is refractory, and recalcitrant to treat with standard antimicrobials. When human babesiosis is advanced, this zoonosis is commonly recrudescent, and often associated with the presence of severe anemia and persistent parasitemia [99,109,110,111].
Lyme disease is a zoonosis with multisystemic clinical manifestations in humans. Bbsl is pleomorphic with diverse forms (i.e., spirochetes, spherocytes, blebs, granules) and, collectively, as dormant biofilms [9,12,112,113,114]. Lyme disease spirochetes have an affinity for immune privileged sites, and side-step the immune response, and lodge in niche reservoirs including bone [115], brain [116,117,118], eye [119], muscle [120], collagenous tissues (ligaments, tendons) [121,122], glial and neuronal cells [123,124,125], and fibroblasts/scar tissue [126]. Left untreated or inadequately treated, this insidious spirochetosis can be persistent [9,10,11,113,116,127,128,129], and develop into chronic Lyme disease [12,13]. Often, patients advance to chronic Lyme disease before they get diagnosed and treated. Psychiatric illness, caused by Lyme disease, may include violence, substance abuse, and developmental disabilities [130,131,132]. Lyme disease may cause severe and potentially fatal central nervous system complications. Although Lyme carditis is known to be fatal in Lyme disease patients, there are multiple other causes of death. Fatal neurological impairments include seizures, grand mal seizures, chronic meningoencephalomyelitis, massive hypocephalus, epilepticus, ependymitis, progressive encephalitis, cerebral atrophy, periventricular white matter disease, and irreversible brain injury [133,134,135]. When the pathologies of neuroborreliosis are unrelenting, the pain in musculoskeletal tissues is unbearable, and somnolence is unending, Lyme disease patients sometimes resort to suicide [130,131,132]. Ultimately, this severely debilitating illness can be fatal [12,116,117,119,133,134,135].
In a study by Fallon et al. [136], the two-tier Lyme disease serological testing had a sensitivity of 49% for patients with persistent symptoms following Lyme disease treatment. Lyme disease patients who use the two-tiered serology testing will often be seronegative, but still have active Bbsl infection [11,12,116,119,121,127,134,135,136,137,138]. Stricker and Johnson also encountered low sensitivity exhibited as false negatives [139]. Since Bbsl biofilms have mechanisms to resist antibiotic challenge, especially in immune-privileged niche tissue, it is adventitious to use a biofilm disruptor (e.g., biofilm buster) to stimulate an immune response prior to blood draw for Lyme disease serology testing [140].
Borrelia burgdorferi sensu lato may be transmitted by congenital passage [141,142,143,144,145] or by blood transfusion [146,147]. Similar to syphilis [148], Bbsl transmission could potentially occur during intimate relationships [11,149].
5. Conclusions
This study highlights three dissimilar Babesia species and three diverse Bbsl genospecies/strains in ticks collected in centralized provinces of Canada. Of epidemiological significance, we detected Borrelia burgdorferi sensu stricto, Babesia divergens-like piroplasm and Babesia microti, and all of these three tick-borne zoonotic microorganisms are pathogenic to humans. Even though Babesia odocoilei was found in several engorged and questing I. scapularis ticks, we cannot decipher at this point if this babesial species is pathogenic to humans. We detected co-infections in ticks, and suggest that more than one infectious microbe can be transmitted simultaneously to the host during a blood meal. To our knowledge, we provide the first enzootic study reporting blacklegged ticks concurrently infected with B. odocoilei and Bbsl. Additionally, we report the first evidence of established populations of I. scapularis on mainland Ontario infected with B. odocoilei. In view of the current findings, we advise that I. scapularis ticks play a pivotal role in the transmission dynamics of B. odocoilei and Bbsl spirochetes. Not only are I. scapularis vectors for multiple tick-borne pathogens, they have the potential to be a bridge vector of B. odocoilei between white-tailed deer and humans and domestic animals. By holding fully engorged ticks to molt, we confirm that Bbsl in I. muris and B. odocoilei in I. scapularis successfully undergo transstadial passage. The detection of B. microti in a groundhog tick constitutes a landmark Babesia discovery for this tick species. We provide the first-ever study that documents a B. divergens-like piroplasm in Canada, and this particular strain is known to be pathogenic to humans. Within the Lyme disease genospecies complex, a Borrelia lanei-like bacterium is unveiled for the first time in Canada east of the Rocky Mountains. Furthermore, we report a unique Bbsl bacterium that may constitute a new genospecies which may be potentially pathogenic to humans.
Of medical importance, not only are Haldimand-Norfolk residents testing positive for human babesiosis and Lyme disease, they are dwelling in environmental strongholds with I. scapularis ticks infected with B. odocoilei and Bbsl. Further etiological research is needed to determine whether B. odocoilei is pathogenic to humans. Such research is essential to explain how some individuals are sick, even gravely sick, but test negative for piroplasms or strains of Bbsl. Healthcare practitioners must have the freedom to use clinical judgment, based on empirical evidence, to treat patients with tick-borne, zoonotic diseases. Even though diagnostics may currently not be available, public health authorities, medical societies, and regulatory colleges need to protect the autonomy of first-line clinicians to utilize their diagnostic skills and clinical acumen for tick-borne zoonoses in Canada. Since bird-feeding ticks are harbouring infectious microbes, our findings suggest that these songbird-transported ticks are widespread. Our data indicate that ticks harbour pathogens associated with Lyme disease and human babesiosis are host-seeking in the Canadian outdoors. Healthcare practitioners must include these zoonoses in their differential diagnoses, and treat them in a forthright manner and with due diligence.
Author Contributions
J.D.S. was responsible for study design, coordinating this tick–host-microbe project, and writing the manuscript. K.L.C., N.M.C., and T.R.B. conducted molecular testing of ticks and analysis on PCR amplicons. All authors read and approved the final manuscript.
Funding
Funding was provided in part by the Mary Alice Holmes Foundation.
Acknowledgments
We thank bird banders, veterinarians, wildlife rehabilitators, road crew workers, Fatal Light Awareness Program staff, and the public for collecting ticks. We are indebted to Amanda Green for computer graphics.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Johnson, L.; Shapiro, M.; Mankoff, J. Removing the mask of average treatment effects in chronic Lyme disease research using Big Data and subgroup analysis. Healthcare 2018, 6, 124. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, W.A.; Sonenshine, D.E.; Noden, B.H. Ticks (Ixodida). In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Academic Press/Elsevier: London, UK, 2019; pp. 603–672. ISBN 978-0-12-814043-7. [Google Scholar]
- Davidsson, M. The financial implications of a well-hidden and ignored chronic Lyme disease pandemic. Healthcare 2018, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Benach, J.L.; Coleman, J.L.; Habicht, G.S.; MacDonald, A.; Grunwaldt, E.; Giron, J.A. Serological evidence for simultaneous occurrences of Lyme disease and babesiosis. J. Infect. Dis. 1985, 152, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Babes, V. Sur l’hemoglobinurie bactérienne du boeuf. C. R. Acad. Sci. Ser. III Sci. Vie 1888, 107, 692–694. [Google Scholar]
- Škrabalo, Z.; Deanović, Z. Piroplasmosis in man: Report on a case. Doc. Med. Geogr. Trop. 1957, 9, 11–16. [Google Scholar]
- Akel, T.; Mobarakai, N. Hematologic manifestations of babesiosis. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 6. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Barbour, A.G.; Hayes, S.F.; Benach, J.L.; Grunwaldt, E.; Davis, J.P. Lyme disease—A tick-borne spirochetosis? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef]
- Miklossy, J.; Kasas, S.; Zurn, A.D.; McCall, S.; Yu, S.; McGeer, P.L. Persisting atypical and cystic forms of Borrelia burgdorferi and local inflammation in Lyme neuroborreiosis. J. Neuroinflamm. 2008, 5, 40. [Google Scholar] [CrossRef]
- Embers, M.E.; Hasenkampf, N.R.; Jacobs, M.B.; Tardo, A.C.; Doyle-Myers, L.A.; Philipp, M.T.; Hodzic, E. Variable manifestations, diverse seroreactivity and post-treatment persistence in non-human primates exposed to Borrelia burgdorferi by tick feeding. PLoS ONE 2017, 12, e0189071. [Google Scholar] [CrossRef]
- Middelveen, M.J.; Sapi, E.; Burke, J.; Filush, K.R.; Franco, A.; Fesler, M.C.; Stricker, R.B. Persistent Borrelia infection in patients with ongoing symptoms of Lyme disease. Healthcare 2018, 6, 33. [Google Scholar] [CrossRef]
- Sapi, E.; Kasliwala, R.S.; Ismail, H.; Torres, J.P.; Oldakowski, M.; Markland, S.; Gaur, G.; Melillo, A.; Eisendle, K.; Liegner, K.B.; et al. The long-term persistence of Borrelia burgdorferi antigens and DNA in the tissues of a patient with Lyme disease. Antibiotics 2019, 8, 183. [Google Scholar] [CrossRef] [PubMed]
- Stricker, R.B.; Fesler, M.C. Chronic Lyme disease: A working case definition. Am. J. Infect. Dis. 2018, 14, 44. [Google Scholar] [CrossRef]
- Anderson, J.F.; Magnarelli, L.A. Avian and mammalian hosts for spirochete-infected ticks and insects in a Lyme disease focus in Connecticut. Yale J. Biol. Med. 1984, 57, 627–641. [Google Scholar] [PubMed]
- Reed, K.D.; Meece, J.K.; Henkel, J.S.; Shukla, S.K. Birds, migration and emerging zoonoses: West Nile virus, Lyme disease, influenza A and enteropathogens. Clin. Med. Res. 2003, 1, 5–12. [Google Scholar] [CrossRef]
- Morshed, M.G.; Scott, J.D.; Fernando, K.; Beati, L.; Mazerolle, D.F.; Geddes, G.; Durden, L.A. Migratory songbirds disperse ticks across Canada, and first isolation of the Lyme disease spirochete, Borrelia burgdorferi, from the avian tick, Ixodes auritulus. J. Parasitol. 2005, 91, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Clark, K.L.; Foley, J.E.; Bierman, B.C.; Durden, L.A. Far-reaching dispersal of Borrelia burgdorferi sensu lato-infected blacklegged ticks by migratory songbirds in Canada. Healthcare 2018, 6, 89. [Google Scholar] [CrossRef]
- Scott, J.D.; Fernando, K.; Banerjee, S.N.; Durden, L.A.; Byrne, S.K.; Banerjee, M.; Mann, R.B.; Morshed, M.G. Birds disperse ixodid (Acari: Ixodidae) and Borrelia burgdorferi-infected ticks in Canada. J. Med. Entomol. 2001, 38, 493–500. [Google Scholar] [CrossRef]
- Scott, J.D.; Durden, L.A. First isolation of Lyme disease spirochete, Borrelia burgdorferi, from ticks collected from songbirds in Ontario, Canada. N. Am. Bird Bander 2009, 34, 97–101. [Google Scholar]
- Scott, J.D.; Lee, M.-K.; Fernando, K.; Durden, L.A.; Jorgensen, D.R.; Mak, S.; Morshed, M.G. Detection of Lyme disease spirochete, Borrelia burgdorferi sensu lato, including three novel genotypes in ticks (Acari: Ixodidae) collected from songbirds (Passeriformes) across Canada. J. Vector Ecol. 2010, 35, 124–139. [Google Scholar] [CrossRef]
- Scott, J.D.; Anderson, J.F.; Durden, L.A. Widespread dispersal of Borrelia burgdorferi-infected ticks collected from songbirds across Canada. J. Parasitol. 2012, 98, 49–59. [Google Scholar] [CrossRef]
- Scott, J.D.; Clark, K.L.; Foley, J.E.; Anderson, J.F.; Bierman, B.C.; Durden, L.A. Extensive distribution of the Lyme disease bacterium, Borrelia burgdorferi sensu lato, in multiple tick species parasitizing avian and mammalian hosts across Canada. Healthcare 2018, 6, 131. [Google Scholar] [CrossRef] [PubMed]
- Brewer, D.A.; Diamond, A.; Woodsworth, E.J.; Collins, B.T.; Dunn, E.H. Doves, cuckoos, and hummingbirds through passerines. In Canadian Atlas of Bird Banding, 1921–1995; Canadian Wildlife Service, Environment Canada: Hull, UK, 2000; Volume 1, pp. 1–395. ISBN 0-662-28946-3. [Google Scholar]
- DeLuca, W.V.; Woodworth, B.K.; Rimmer, C.C.; Marra, P.P.; Taylor, P.D.; McFarland, K.P.; Mackenzie, S.A.; Norris, D.R. Transoceanic migration by a 12 g songbird. Biol. Lett. 2015, 11, 20141045. [Google Scholar] [CrossRef] [PubMed]
- Dunson, W. The Incredible Flight of a Willow Flycatcher. 2014. Available online: http://lemonbayconservancy.org/incredible-flight-willow-flycatcher/ (accessed on 5 November 2018).
- Stutchbury, B.J.M.; Tarof, S.A.; Done, T.; Gow, E.; Kramer, P.M.; Tautin, J.; Fox, J.W.; Afanasyev, V. Tracking long-distance songbird migration by using geolocators. Science 2009, 323, 896. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Lindsay, L.R.; Hanincová, K.; Barker, I.K.; Bigras-Poulin, M.; Charron, D.F.; Heagy, A.; Francis, C.M.; O’Callaghan, C.J.; Schwartz, I.; et al. Role of migratory birds in introduction and range expansion of I. scapularis ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada. Appl. Environ. Microbiol. 2008, 74, 1780–1790. [Google Scholar] [CrossRef]
- Scott, J.D.; Durden, L.A. New records of the Lyme disease bacterium in ticks collected from songbirds in central and eastern Canada. Int. J. Acarol. 2015, 41, 241–249. [Google Scholar] [CrossRef]
- Scott, J.D.; Durden, L.A. Amblyomma dissimile Koch (Acari: Ixodidae) parasitizes bird captured in Canada. Syst. Appl. Acarol. 2015, 20, 854–860. [Google Scholar]
- Scott, J.D.; Durden, L.A. First record of Amblyomma rotundatum tick (Acari: Ixodidae) parasitizing a bird collected in Canada. Syst. Appl. Acarol. 2015, 20, 155–161. [Google Scholar]
- Scott, J.D. Birds widely disperse pathogen-infected ticks. In Seabirds and Songbirds: Habitat Preferences, Conservation, Migratory Behavior; Mahala, G., Ed.; Nova Publishers, Inc.: New York, NY, USA, 2015; pp. 1–22. ISBN 978-1-63463-496-0. [Google Scholar]
- Scott, J.D.; Scott, C.M.; Anderson, J.F. The establishment of a blacklegged tick population by migratory songbirds in Ontario, Canada. J. Vet. Sci. Med. 2014, 2, 5. [Google Scholar] [CrossRef]
- Morshed, M.G.; Scott, J.D.; Fernando, K.; Geddes, G.; McNabb, A.; Mak, S.; Durden, L.A. Distribution and characterization of Borrelia burdgorferi isolates from Ixodes scapularis and presence in mammalian hosts in Ontario, Canada. J. Med. Entomol. 2006, 43, 762–773. [Google Scholar] [CrossRef]
- Scott, J.D.; Fernando, K.; Durden, L.A.; Morshed, M.G. Lyme disease spirochete, Borrelia burgdorferi, endemic in epicenter at Turkey Point, Ontario. J. Med. Entomol. 2004, 41, 226–230. [Google Scholar] [CrossRef]
- Scott, J.D.; Lee, M.-K.; Fernando, K.; Jorgensen, D.R.; Durden, L.A.; Morshed, M.G. Rapid introduction of Lyme disease spirochete, Borrelia burgdorferi sensu stricto, in Ixodes scapularis (Acari: Ixodidae) established at Turkey Point Provincial Park, Ontario, Canada. J. Vector Ecol. 2008, 33, 64–69. [Google Scholar] [CrossRef]
- Morshed, M.G.; Scott, J.D.; Banerjee, S.N.; Fernando, K.; Mann, R.; Isaac-Renton, J. First isolation of Lyme disease spirochete, Borrelia burgdorferi from blacklegged tick, Ixodes scapularis, collected at Rondeau Provincial Park, Ontario. Can. Com. Dis. Rep. 2000, 26, 42–44. [Google Scholar]
- Morshed, M.G.; Scott, J.D.; Fernando, K.; Mann, R.B.; Durden, L.A. Lyme disease spirochete, Borrelia burgdorferi endemic at epicenter in Rondeau Provincial Park, Ontario. J. Med. Entomol. 2003, 40, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Anderson, J.F.; Durden, L.A.; Smith, M.L.; Manord, J.M.; Clark, K.L. Prevalence of the Lyme disease spirochete, Borrelia burgdorferi, in blacklegged ticks, Ixodes scapularis at Hamilton-Wentworth, Ontario. Int. J. Med. Sci. 2016, 13, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Clark, K.L.; Coble, N.M.; Ballantyne, T.R. Presence of Babesia odocoilei and Borrelia burgdorferi sensu stricto in a tick and dual parasitism of Amblyomma inornatum and Ixodes scapularis on a bird in Canada. Healthcare 2019, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Milnes, E.L.; Thornton, G.; Léveillé, A.N.; Delinatte, P.; Barta, J.R.; Smith, D.A.; Nemeth, N. Babesia odocoilei and zoonotic pathogens identified from Ixodes scapularis ticks in southern Ontario, Canada. Ticks Tick-Borne Dis. 2019, 10, 670–676. [Google Scholar] [CrossRef]
- Steiner, F.E.; Pinger, R.R.; Vann, C.N.; Abley, M.J.; Sullivan, B.; Grindle, N.; Clay, K.; Fuqua, C. Detection of Anaplasma phagocytophilum and Babesia odocoilei DNA in Ixodes scapularis (Acari: Ixodidae) collected in Indiana. J. Med. Entomol. 2006, 43, 437–442. [Google Scholar] [CrossRef][Green Version]
- Shock, B.C.; Moncayo, A.; Cohen, S.; Mitchell, E.A.; Williamson, P.C.; Lopez, G.; Garrison, L.E.; Yabsley, M.J. Diversity of piroplasms detected in blood-fed and questing ticks from several states in the United States. Ticks Tick-Borne Dis. 2014, 5, 373–380. [Google Scholar] [CrossRef]
- Available online: https://flap.org (accessed on 18 November 2019).
- Keirans, J.E.; Durden, L.A. Illustrated key to nymphs of the tick genus Amblyomma (Acari: Ixodidae) found in the United States. J. Med. Entomol. 1998, 35, 489–495. [Google Scholar] [CrossRef]
- Guzmán–Cornejo, C.; Robbins, R.G.; Guglielmone, A.A.; Montiel–Parra, G.; Pérez, M. The Amblyomma (Acari: Ixodida: Ixodidae) of Mexico: Identification keys, distribution and hosts. Zootaxa 2011, 2998, 16–38. [Google Scholar]
- Jones, E.K.; Clifford, C.M.; Keirans, J.E.; Kohls, G.M. Ticks of Venezuela (Acarina: Ixodoidea) with a Key to the Species of Amblyomma in the Western Hemisphere; Biological Series; Brigham Young University Science Bulletin: Provo, Utah, 1972; Volume VXII, pp. 1–40. [Google Scholar]
- Clifford, C.M.; Anastos, G.; Elbl, A. The larval ixodid ticks of the eastern United States. Misc. Publ. Entomol. Soc. Am. 1961, 2, 213–237. [Google Scholar]
- Durden, L.A.; Keirans, J.E. Nymphs of the Genus Ixodes (Acari: Ixodidae) of the United States: Taxonomy, Identification Key, Distribution, Hosts, and Medical/Veterinary Importance. Monographs; Thomas Say Publications in Entomology, Entomological Society of America: Lanham, MD, USA, 1996; p. 95. ISBN 0-938522-57. [Google Scholar]
- Keirans, J.E.; Clifford, C.M. The genus Ixodes in the United States: A scanning electron microscope study and key to the adults. J. Med. Entomol. 1978, 15 (Suppl. S2), 1–38. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Hendricks, A.; Burge, D. Molecular identification and analysis of Borrelia burgdorferi sensu lato in lizards in the southeastern United States. Appl. Environ. Microbiol. 2005, 71, 2616–2625. [Google Scholar] [CrossRef] [PubMed]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.-C. Presence of potentially pathogenic Babesia sp. for human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2016, 13, 65–70. [Google Scholar]
- McCombie, W.R.; Heiner, C.; Kelly, J.M.; Fitzgerald, M.G.; Gocayne, J.D. Rapid and reliable fluorescent cycle sequencing of double stranded templates. DNA Seq. 1992, 2, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX–Windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tools. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Caporale, D.A.; Johnson, C.M.; Millard, B.J. Presence of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae) in southern Kettle Moraine State Forest, Wisconsin, and characterization of strain W97F51. J. Med. Entomol. 2005, 42, 457–472. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Shein, E. The piroplasms: Life cycle and sexual stages. Adv. Parasitol. 1984, 23, 37–103. [Google Scholar]
- Telford, S.R.; Gorenflot, A.; Brasseur, P.; Spielman, A. Babesial infections in humans and wildlife. In Parasitic Protozoa; Kresier, J.P., Ed.; Academic Press: San Diego, CA, USA, 1993; Volume 5, pp. 1–47. [Google Scholar]
- Holman, P.J.; Madeley, J.; Craig, T.M.; Allsopp, B.A.; Allsopp, M.T.; Petrini, K.R.; Waghela, S.D.; Wagner, G.G. Antigenic, phenotypic and molecular characterization confirms Babesia odocoilei isolated from three cervids. J. Wildl. Dis. 2000, 36, 518–530. [Google Scholar] [CrossRef]
- Gorenflot, A.; Moubri, K.; Percigout, E.; Carey, B.; Schetters, T.P. Human babesiois. Ann. Trop. Med. Parasitol. 1998, 92, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; de Bruyn, G.; Pieniazek, N.J.; Homer, M.; Lofy, K.H.; Shemenda, S.B.; Fritsche, T.R.; Persing, D.H.; Limaye, A.P. Babesia divergens-like infection, Washington state. Emerg. Infect. Dis. 2004, 10, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Kjemtrup, A.M.; Conrad, P.A. Human babesiosis: An emerging tick-borne disease. Int. J. Parasitol. 2000, 30, 1323–1337. [Google Scholar] [CrossRef]
- Herwaldt, B.L.; Cacciò, S.; Gherlinzoni, F.; Aspöck, H.; Siemenda, S.B.; Piccaluga, P.; Martinelli, G.; Edelhofer, R.; Hollenstein, U.; Poletti, G.; et al. Molecular characterization of a non-Babesia divergens organism causing zoonotic babesiosis in Europe. Emerg. Infect. Dis. 2003, 9, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Pattullo, K.M.; Wobeser, G.; Lockerbie, B.P.; Burgess, H.J. Babesia odocoilei infection in a Sakatchewan elk (Cervus elaphus canadensis) herd. J. Vet. Diagn. Investig 2013, 25, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.F. Mammalian and avian reservoirs for Borrelia burgdorferi. Ann. N. Y. Acad. Sci. 1988, 539, 180–191. [Google Scholar] [CrossRef]
- Richter, D.; Spielman, A.; Komar, N.; Matuschka, F.-R. Competence of American Robins as reservoir hosts for Lyme disease spirochetes. Emerg. Infect. Dis. 2000, 6, 133–138. [Google Scholar] [CrossRef]
- Rollend, L.; Fish, D.; Childs, J.E. Transovarial transmission of Borrelia spirochetes by Ixodes scapularis: A summary of the literature and recent observations. Ticks Tick-Borne Dis. 2013, 4, 46–51. [Google Scholar] [CrossRef]
- Mannelli, A.; Kitron, U.; Jones, C.J.; Slajchert, T.L. Influence of season and habitat on Ixodes scapularis infestation on white-footed mice in northeastern Illinois. J. Parasitol. 1994, 80, 1038–1043. [Google Scholar] [CrossRef]
- Kurtenbach, K.; Carey, D.; Hoodless, A.N.; Nuttall, P.A.; Randolph, S.E. Competence of pheasants as reservoirs for Lyme disease spirochetes. J. Med. Entomol. 1998, 35, 77–81. [Google Scholar] [CrossRef]
- Littman, M.P.; Goldstein, R.E.; Labato, M.A.; Lappin, M.R.; Moore, G.E. ACVIM small animal consensus statement on Lyme disease in dogs: Diagnosis, treatment, and prevention. J. Vet. Intern. Med. 2006, 20, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Straubinger, R.K.; Summers, B.A.; Chang, Y.F.; Appel, M.J. Persistence of Borrelia burgdorferi in experimentally infected dogs after antibiotic treatment. J. Clin. Microbiol. 1997, 35, 111–116. [Google Scholar] [PubMed]
- Divers, T.J.; Gardner, R.B.; Madigan, J.E.; Witonsky, S.G.; Bertone, J.J.; Swinebroad, E.L.; Schutzer, S.E.; Johnson, A.L. Borrelia burgdorferi infection and Lyme disease in North American horses: A consensus statement. J. Vet. Intern. Med. 2018, 32, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Burgess, E.C.; Gendron-Fitpatrick, A.; Mattison, M. Foal mortality associated with natural infection of pregnant mares with Borrelia burgdorferi. In Proceedings of the Fifth International Conference of Equine Infectious Diseases; Press of Kentucky: Lexington, KY, USA, 1988; pp. 217–220. [Google Scholar]
- Scott, J.D.; Clark, K.L.; Anderson, J.F.; Foley, J.E.; Young, M.R.; Durden, L.A. Lyme disease bacterium, Borrelia burgdorferi sensu lato, detected in multiple tick species at Kenora, Ontario, Canada. J. Bacteriol. Parasitol. 2017, 8, 304. [Google Scholar] [CrossRef]
- Heisig, M.; Abraham, N.M.; Liu, L.; Neelakanta, G.; Mattessich, S.; Sultana, H.; Shang, Z.; Anari, J.M.; Killiam, C.; Walker, W.; et al. Anti-virulence properties of an antifreeze protein. Cell Rep. 2014, 9, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Scott, C.M. Lyme disease propelled by Borrelia burgdorferi-infected blacklegged ticks, wild birds and public awareness—Not climate change. J. Vet. Sci. Med. 2018, 6, 8. [Google Scholar]
- Scott, J.D.; Schillberg, E.; Lunny, D.; Lindsay, L.; Nelder, M.; Russell, C.; Mackie, M.; Coats, D.; Berry, A.; Young Hoon, K. Distribution of Ixodes scapularis in Northwestern Ontario: Results from active and passive activities in the Northwestern Health Unit catchment area. Int. J. Environ. Res. Public Health 2019, 16, 1939. [Google Scholar] [CrossRef]
- McLean, R.G.; Ubico, S.R.; Cooksey, L.M. Experimental infection of the eastern chipmunk (Tamias striatus) with the Lyme disease spirochete (Borrelia burgdorferi). J. Wildl. Dis. 1993, 29, 527–532. [Google Scholar] [CrossRef]
- Holman, P.J.; Waldrup, K.A.; Wagner, G.G. In vitro cultivation of a Babesia isolated from a white–tailed deer (Odocoileus virginianus). J. Parasitol. 1988, 74, 111–115. [Google Scholar] [CrossRef]
- Hersh, M.H.; Osfeld, R.S.; McHenry, D.J.; Tibbetts, M.; Brunner, J.L.; Killilea, M.E.; LoGiudice, K.; Schmidt, K.A.; Keesing, F. Co-infestation of blacklegged ticks with Babesia microti and Borrelia burgdorferi is higher than expected and acquired from small mammal hosts. PLoS ONE 2014, 9, e99348. [Google Scholar] [CrossRef]
- Hall, J.E.; Amrine, J.W., Jr.; Gais, R.D.; Kolanko, V.P.; Hagenbuch, B.E.; Gerencser, V.F.; Clark, S.M. Parasitization of humans in West Virginia by Ixodes cookei (Acari: Ixodidae), a potential vector of Lyme borreliosis. J. Med. Entomol. 1991, 28, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Merten, H.A.; Durden, L.A. A state-by-state survey of ticks recorded from humans in the United States. J. Vect. Ecol. 2000, 25, 102–113. [Google Scholar]
- Scott, J.D.; Foley, J.E.; Anderson, J.F.; Clark, K.L.; Durden, L.A. Detection of Lyme disease bacterium, Borrelia burgdorferi sensu lato, in blacklegged ticks collected in the Grand River Valley, Ontario, Canada. Int. J. Med. Sci. 2017, 14, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, K.; Chandrashekar, R.; Beall, M.; Leutenegger, C.; Lappin, M.R. Evidence for clinical anaplasmosis and borreliosis in cats in Maine. Top. Companion Anim. Med. 2018, 33, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Savick, K.; Butler, J. Babesia microti in rodents and raccoons from northeast Florida. J. Parasitol. 2012, 98, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.F.; Mintz, E.D.; Gadbaw, J.J.; Magnarelli, L.A. Babesia microti, human babesiosis, and Borrelia burgdorferi in Connecticut. J. Clin. Microbiol. 1991, 29, 2779–2783. [Google Scholar]
- Anderson, J.F.; Armstrong, P.M. Prevalence and genetic characterization of Powassan virus strains infecting Ixodes scapularis in Connecticut. Am. J. Trop. Med. Hyg. 2012, 87, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Telford, S.R., III; Mather, T.N.; Moore, S.I.; Wilson, M.L.; Spielman, A. Incompetence of deer as reservoirs of the Lyme disease spirochete. Am. J. Trop. Med. Hyg. 1988, 39, 105–109. [Google Scholar] [CrossRef]
- Brown, J.H. The rabbit tick Haemaphysalis leporispalustris (Pack.) as an ectoparasite on man. Can. Entomol. 1945, 77, 176. [Google Scholar] [CrossRef]
- Banerjee, S.N.; Banerjee, M.; Fernando, K.; Dong, M.Y.; Smith, J.A.; Cook, D. Isolation of Borrelia burgdorferi, the Lyme disease spirochete from rabbit ticks, Haemaphysalis leporispalustris from Alberta. J. Spir. Tick-Borne Dis. 1995, 2, 23–24. [Google Scholar]
- Herc, E.; Pritt, B.; Huizenga, T.; Douce, R.; Hysell, M.; Newton, D.; Sidge, J.; Losman, E.; Sherbeck, J.; Kaul, D.R. Probable locally acquired Babesia divergens-like infection in woman, Michigan, USA. Emerg. Infect. Dis. 2018, 24, 1558–1560. [Google Scholar] [CrossRef] [PubMed]
- Goethert, H.K.; Telford, S.R. Enzootic transmission of Babesia divergens among cottontail rabbits on Nantucket Island, Massachusetts. Am. J. Trop. Med. Hyg. 2003, 69, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Clark, K.L.; Foley, J.E.; Anderson, J.F.; Durden, L.A.; Manord, J.M.; Smith, M.L. Detection of Borrelia genomospecies 2 in Ixodes spinipalpis ticks collected from a rabbit in Canada. J. Parasitol. 2017, 103, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Chagnon, S.L.; Naik, M.; Abdel-Hamid, H. Child Neurology: Tick paralysis: A diagnosis not to miss. Neurology 2014, 82, e91–e93. [Google Scholar] [CrossRef]
- Scott, J.D. First record of locally acquired human babesiosis in Canada caused by Babesia duncani: A case report. SAGE Open Med. Case Rep. 2017, 5, 2050313X17725645. [Google Scholar]
- Benach, J.L.; Habicht, G.S. Clinical characteristics of human babesiosis. J. Infect. Dis. 1981, 144, 481. [Google Scholar] [CrossRef]
- Homer, M.J.; Aquilar-Delfin, I.; Telford, S.R., III; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef]
- Ruebush, T.K.; Cassaday, P.B.; Marsh, H.J.; Lisker, S.A.; Voorhees, D.B.; Mahoney, E.B.; Healy, G.R. Human babesiosis on Nantucket Island. Ann. Intern. Med. 1977, 86, 6–9. [Google Scholar] [CrossRef]
- Mayne, P.J. Clinical determinants of Lyme borreliosis, babesiosis, bartonellosis, anaplasmosis, and ehrlichiosis in an Australian cohort. Int. J. Gen. Med. 2014, 8, 15–26. [Google Scholar] [CrossRef]
- Hatcher, J.C.; Greenberg, P.D.; Antique, J.; Jimenez-Lucho, V.E. Severe babesiosis in Long Island: Review of 34 cases and their complications. Clin. Infect. Dis. 2001, 32, 1117. [Google Scholar] [CrossRef]
- Marcus, L.C.; Steere, A.C.; Duray, P.H.; Anderson, A.E.; Mahoney, E.B. Fatal pancarditis in a patient with coexistent Lyme disease and babesiosis: Demonstration of spirochete in myocardium. Ann. Intern. Med. 1985, 103, 374–376. [Google Scholar] [CrossRef] [PubMed]
- LeBel, D.P.; Moritz, E.D.; O’Brien, J.J.; Lazarchick, J.; Tormos, L.M.; Duong, A.; Fontaine, M.J.; Squires, J.E.; Stramer, S.L. Cases of transfusion-transmitted babesiosis occurring in nonendemic areas: A diagnostic dilemma. Transfusion 2017, 57, 2348–2354. [Google Scholar] [CrossRef] [PubMed]
- Bloch, E.M.; Levin, A.E.; Williamson, P.C.; Cyrus, S.; Shaz, B.H.; Kessler, D.; Gorlin, J.; Bruhn, R.; Lee, T.-H.; Montalvo, L.; et al. A prospective evaluation of chronic Babesia microti infection in seroreactive blood donors. Transfusion 2016, 56, 1875–1882. [Google Scholar] [CrossRef] [PubMed]
- Villatoro, T.; Karp, J.K. Transfusion-transmitted babesiosis. Arch. Pathol. Lab. Med. 2019, 143, 130–134. [Google Scholar] [CrossRef]
- Klevens, R.M.; Cumming, M.A.; Caten, E.; Stramer, S.L.; Townsend, R.L.; Tonnetti, L.; Rios, J.; Young, C.T.; Soliva, S.; DeMaria, A., Jr. Transfusion-transmitted babesiosis: One state’s experience. Transfusion 2018, 58, 2611–2616. [Google Scholar] [CrossRef] [PubMed]
- Fox, L.M.; Winger, S.; Ahmed, A.; Arnold, A.; Chou, J.; Rhein, L.; Levy, O. Neonatal babesiosis: Case report and review of the literature. Pediatr. Infect. Dis. J. 2006, 25, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Cornett, J.K.; Malhotra, A.; Hart, D. Vertical transmission of babesiosis from a pregnant, splenectomized mother to her neonate. Infect. Dis. Clin. Pract. 2012, 20, 408–410. [Google Scholar] [CrossRef]
- Iyer, S.; Goodman, K. Congenital babesiosis from maternal exposure: A case report. J. Emerg. Med. 2009, 56, e39–e41. [Google Scholar] [CrossRef]
- Khangura, R.K.; Williams, N.; Cooper, S.; Prabulos, A.M. Babesiosis in pregnancy: An imitator of HELLP syndrome. AJP Rep. 2019, 9, e147–e152. [Google Scholar] [CrossRef]
- Krause, P.J.; Spielman, A.; Telford, S.R., III; Sikand, V.K.; McKay, K.; Christianson, D.; Pollack, R.J.; Brassard, P.; Magera, J.; Ryan, R.; et al. Persistent parasitemia after acute babesiosis. N. Engl. J. Med. 1998, 339, 160–165. [Google Scholar] [CrossRef]
- Abraham, A.; Brasov, I.; Thekkiniath, J.; Kilian, N.; Lawres, L.; Gao, R.; DeBus, K.; He, L.; Yu, X.; Zhu, G.; et al. Establishment of a continuous in vitro culture of Babesia duncani in human erythrocytes reveals unusually high tolerance to recommended therapies. J. Biol. Chem. 2018, 293, 19974–19981. [Google Scholar] [CrossRef] [PubMed]
- Liegner, K.B. Disulfiram (tetraethylthiuram disulfide) in the treatment of Lyme disease and babesiosis: Report of experience in three cases. Healthcare 2019, 8, 72. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, A.B. Concurrent neotropical borreliosis and Alzheimer’s Disease. Ann. N. Y. Acad. Sci. 1988, 539, 468–470. [Google Scholar] [CrossRef]
- Sapi, E.; MacDonald, A. Biofilms of Borrelia burgdorferi in chronic cutaneous borreliosis. Am. J. Clin. Pathol. 2008, 129, 988–989. [Google Scholar]
- Meriläinen, L.; Herranen, A.; Schwarzbach, A.; Gilbert, L. Morphological and biochemical features of Borrelia burgdorferi pleomorphic forms. Microbiology 2015, 161, 516–527. [Google Scholar] [CrossRef]
- Oksi, J.; Mertsola, J.; Reunanen, M.; Marjamäki, M.; Viljanen, M.K. Subacute multiple-site osteomyelitis caused by Borrelia burgdorferi. Clin. Infect. Dis. 1994, 19, 891–896. [Google Scholar] [CrossRef]
- Oksi, J.; Kalimo, H.; Marttila, R.J.; Marjamäki, M.; Sonninen, P.; Nikoskelainen, J.; Viljanen, M.K. Inflammatory brain changes in Lyme borreliosis: A report on three patients and review of literature. Brain 1996, 119, 2143–2154. [Google Scholar] [CrossRef]
- MacDonald, A.B. Alzheimer’s neuroborreliosis and trans-synaptic spread of infection and neurofibrillary tangles derived from intraneuronal spirochetes. Med. Hypotheses 2007, 68, 822–825. [Google Scholar] [CrossRef]
- Miklossy, J. Alzheimer’s disease—A neurospirochetosis. Analysis of the evidence following Koch’s and Hill’s criteria. J. Neuroinflamm. 2011, 8, 90. [Google Scholar] [CrossRef]
- Preac-Mursic, V.; Pfister, H.W.; Spiegel, H.; Burk, R.; Wilske, B.; Reinhardt, S.; Böhmer, R. First isolation of Borreia burgdorferi from an iris biopsy. J. Clin. Neuroophthalmol. 1993, 13, 155–161. [Google Scholar]
- Frey, M.; Jaulhac, B.; Piemont, Y.; Marcellin, L.; Boohs, P.M.; Vautravers, P.; Jesel, M.; Kuntz, J.L.; Monteil, H.; Sibilia, J. Detection of Borrelia burgdorferi DNA in muscle of patients with chronic myalgia related to Lyme disease. Am. J. Med. 1998, 104, 591–594. [Google Scholar] [CrossRef]
- Häupl, T.; Hahn, G.; Rittig, M.; Krause, A.; Schoerner, C.; Schönherr, U.; Kalden, J.R.; Burmester, G.R. Persistence of Borrelia burgdorferi in ligamentous tissue from a patient with chronic Lyme borreliosis. Arthritis Rheum. 1993, 36, 1621–1626. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.E. Damage of collagen and elastic fibres by Borrelia burgdorferi—Known and new clinical histopathogical aspects. Open Neurol. J 2012, 6 (Suppl. S1-M11), S179–S186. [Google Scholar]
- Livengood, J.A.; Gilmore, R.D., Jr. Invasion of human neuronal and glial cells by an infectious strain of Borrelia burgdorferi. Microbes Infect. 2006, 8, 2832–2840. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, G.; Borda, J.T.; Dufour, J.; Kaushal, D.; Ramamoorthy, R.; Lackner, A.A.; Philipp, M.T. Interaction of the Lyme disease spirochete Borrelia burgdorferi with brain parenchyma elicits inflammatory mediators from glial cells as well as glial and neuronal apoptosis. Am. J. Pathol. 2008, 173, 1415–1427. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, G.; Santana-Gould, L.; Inglis, F.M.; England, J.D.; Philipp, M.T. The Lyme disease spirochete Borrelia burgdorferi induces inflammation and apoptosis in cells from dorsal root ganglia. J. Neuroinflamm. 2013, 10, 88. [Google Scholar] [CrossRef]
- Klempner, M.S.; Noring, R.; Rogers, R.A. Invasion of human skin fibroblasts by the Lyme disease spirochete, Borrelia burgdorferi. J. Infect. Dis. 1993, 167, 1074–1081. [Google Scholar] [CrossRef]
- Stricker, R.B. Counterpoint: Long-term antibiotic therapy improves persistent symptoms associated with Lyme disease. Clin. Infect. Dis. 2007, 45, 149–157. [Google Scholar] [CrossRef]
- Hodzic, E.; Feng, S.; Holden, K.; Freet, K.J.; Barthold, S.W. Persistence of Borrelia burgdorferi following antibiotic treatment in mice. Antimicrob. Agents Chemother. 2008, 52, 1728–1736. [Google Scholar] [CrossRef]
- Embers, M.E.; Barthold, S.W.; Borda, J.T.; Bowers, L.; Doyle, L.; Hodzic, E.; Jacobs, M.B.; Hasenkampf, N.R.; Martin, D.S.; Narasimhan, S.; et al. Persistence of Borrelia burgdorferi in rhesus macaques following antibiotic treatment of disseminated infection. PLoS ONE 2012, 7, e29914. [Google Scholar] [CrossRef]
- Bransfield, R.C. Suicide and Lyme and associated diseases. Neuropsychiatr. Dis. Treat. 2017, 13, 1575–1587. [Google Scholar] [CrossRef] [PubMed]
- Bransfield, R.C. Aggressiveness, violence, homocidality, homicide, and Lyme disease. Neuropsychiatr. Dis. Treat. 2018, 14, 693–713. [Google Scholar] [CrossRef] [PubMed]
- Bransfield, R.C.; Cook, M.J.; Bransfield, D.R. Proposed Lyme disease guidelines and psychiatric illnesses. Healthcare 2019, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Liegner, K.B.; Ziska, M.; Agricola, M.D.; Hubbard, J.D.; Klempner, M.S.; Coyle, P.K.; Bayer, M.E.; Duray, P.H. Fatal chronic meningoencephalomyelitis with massive hydocephalus, in a New York state patient with evidence of Borrelia burgdorferi (Bb) exposure. In Proceedings of the Sixth International Conference on Lyme Borreliosis, Bologna, Italy, 19–22 June 1994. Abstract P041T. [Google Scholar]
- Liegner, K.B.; Duray, P.; Agricola, M.; Rosenkilde, C.; Yannuzzi, L.A.; Ziska, M.; Tilton, R.C.; Hulinska, D.; Hubbard, J.; Fallon, B.A. Lyme disease and the clinical spectrum of antibiotic responsive chronic meningoencephalomyelitides. J. Spir. Tick-Borne Dis. 1997, 4, 61–73. [Google Scholar]
- Liegner, K.B.; Jones, C.R. Fatal progressive encephalitis following an untreated deer tick attachment in a 7 year-old Fairfield County, Connecticut child. In Proceedings of the VIII International Conference on Lyme Borreliosis, Abstract P380, Munich, Germany, 20–24 June 1999. [Google Scholar]
- Fallon, B.A.; Pavlicova, M.; Coffino, S.W.; Brenner, C. A comparison of Lyme disease serologic test results from four laboratories in patients with persistent symptoms after antibiotic treatment. Clin. Infect. Dis. 2014, 59, 1705–1710. [Google Scholar] [CrossRef] [PubMed]
- Paparone, P.W.; Paparone, P.A. Lyme disease in the elderly. J. Spir. Tick-Borne Dis. 1995, 2, 14–18. [Google Scholar]
- Schutzer, S.E.; Coyle, P.K.; Reid, P.; Holland, B. Borrelia burgdorferi-specific immune complexes in acute Lyme disease. JAMA 1999, 282, 1942–1945. [Google Scholar] [CrossRef]
- Stricker, R.B.; Johnson, L. Lyme disease diagnosis and treatment: Lessions from the AIDS epidemic. Minerva Med. 2010, 101, 419–425. [Google Scholar]
- Berndtson, K. Review of evidence from immune evasion and persistent infection in Lyme disease. Int. J. Gen. Med. 2013, 6, 291–306. [Google Scholar] [CrossRef]
- Feder, H. Borrelia infections: Lyme disease. In Remington and Klein’s Infectious Diseases of the Fetus and Newborn Infant, 8th ed.; Wilson, C.B., Nizet, V., Maldonado, Y., Remington, J.S., Klein, J.O., Eds.; Elsevier: Philadelphia, PA, USA, 2016; pp. 544–557. ISBN 9780321472. [Google Scholar]
- MacDonald, A.B. Gestational Lyme borreliosis: Implications for the fetus. In Rheumatic Disease Clinics of North America; Johnson, R.C., Ed.; Saunders: Philadelphia, PA, USA, 1989; Volume 15, pp. 657–677. Available online: http://www.molecularalzheimer.org/files/Gestational_Lyme_Borreliosis-Annotated_1989.pdf (accessed on 14 October 2017).
- Horowitz, R.I. Lyme disease and pregnancy: Implications of chronic infection, PCR testing, and prenatal treatment. In Proceedings of the 16th International Scientific Conference on Lyme Disease & Other Tick-Borne Disorders, Hartford, CT, USA, 7–8 June 2003. [Google Scholar]
- Trevisan, G.; Stinco, G.; Cinco, M. Neonatal skin lesions due to spirochetal infection: A case of congenital Lyme bororeliosis? Int. J. Dermatol. 1997, 36, 677–680. [Google Scholar]
- Lavoie, P.E.; Lattner, B.P.; Duray, P.H.; Barbour, A.G.; Johnson, P.C. Culture positive seronegative transplacental Lyme borreliosis infant mortality. Arthritis Rheum. 1987, 30, S50. [Google Scholar]
- Burrascano, J.J. Transmission of Borrelia burgdorferi by blood transfusion. In Proceedings of the V International Conference on Lyme Borreliosis, Biology of Borrelia Burgdorferi II, Abstract 265A, Arlington, VA, USA, 30 May–2 June 1992. [Google Scholar]
- Sapi, E.; Pabbati, N.; Datar, A.; Davies, E.M.; Kuo, B.A. Improved culture conditions for the growth and detection of Borrelia from human serum. Int. J. Med. Sci 2013, 10, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Schaudinn, F.R.; Hoffmann, E. VorläuFigureer Bericht über das Vorkommen von Spirochaeten in syphilitischen Krankheitsprodukten und bei Papillomen. Arb. Kais. Gesundh. 1905, 22, 527–534. [Google Scholar]
- Fesler, M.C.; Middelveen, M.J.; Burke, J.M.; Stricker, R.B. Erosive vulvovaginitis associated with Borrelia burgdorferi infection. J. Investig. Med. High Impact Case Rep. 2019, 7, 2324709619842901. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).