


Network Pharmacological Analysis on the Herbal Combinations for Mitigating Inflammation in Respiratory Tracts and Experimental Evaluation
 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Network Pharmacological Analysis
2.2. Preparation of MHRCs
2.3. Cell Culture and Nitrite Assay
2.4. NCI-H292 Cell Culture and PMA-Induced MUC5AC mRNA Expression
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Synergy Scores
2.7. Design of Experiment (DOE) and Design Space (DS)
- Evaluation of the impact between factors and response value; This identifies whether factors and response correlate with each other, and finds out the tendency.
- Identification of major factors affecting response value; The major factors determining the responses are identified through this step and it confirms the degree of influence.
- Creation of a statistical model for the factors and response correlations; This creates an alternative model that approximates the correlation between factors and response.
- An optimal design solution to satisfy responses; The combination of factors is found that obtains the most desirable responses.
2.8. Estimated Regression Equations
3. Results
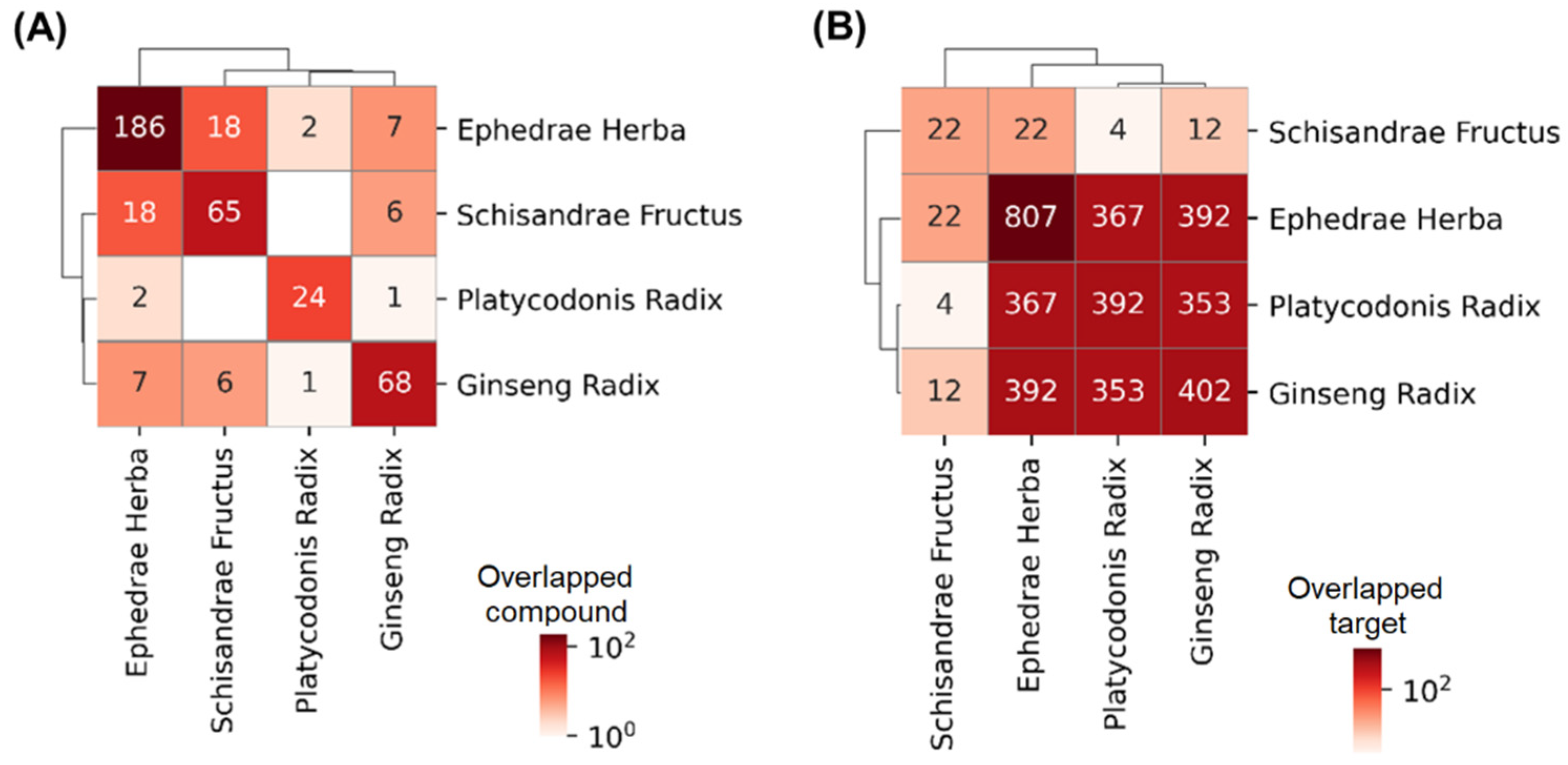
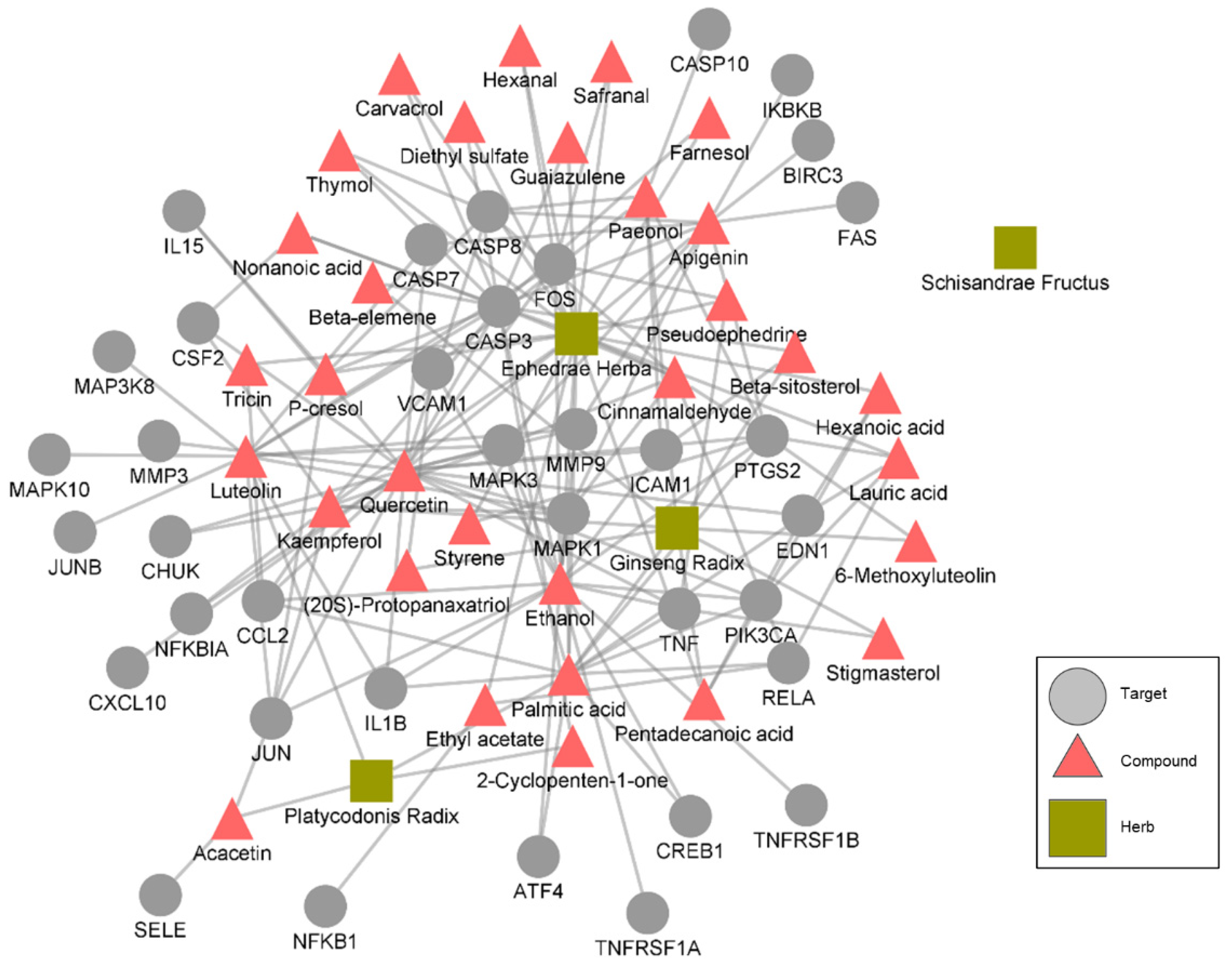
3.1. Association of the Drugs and COVID-19
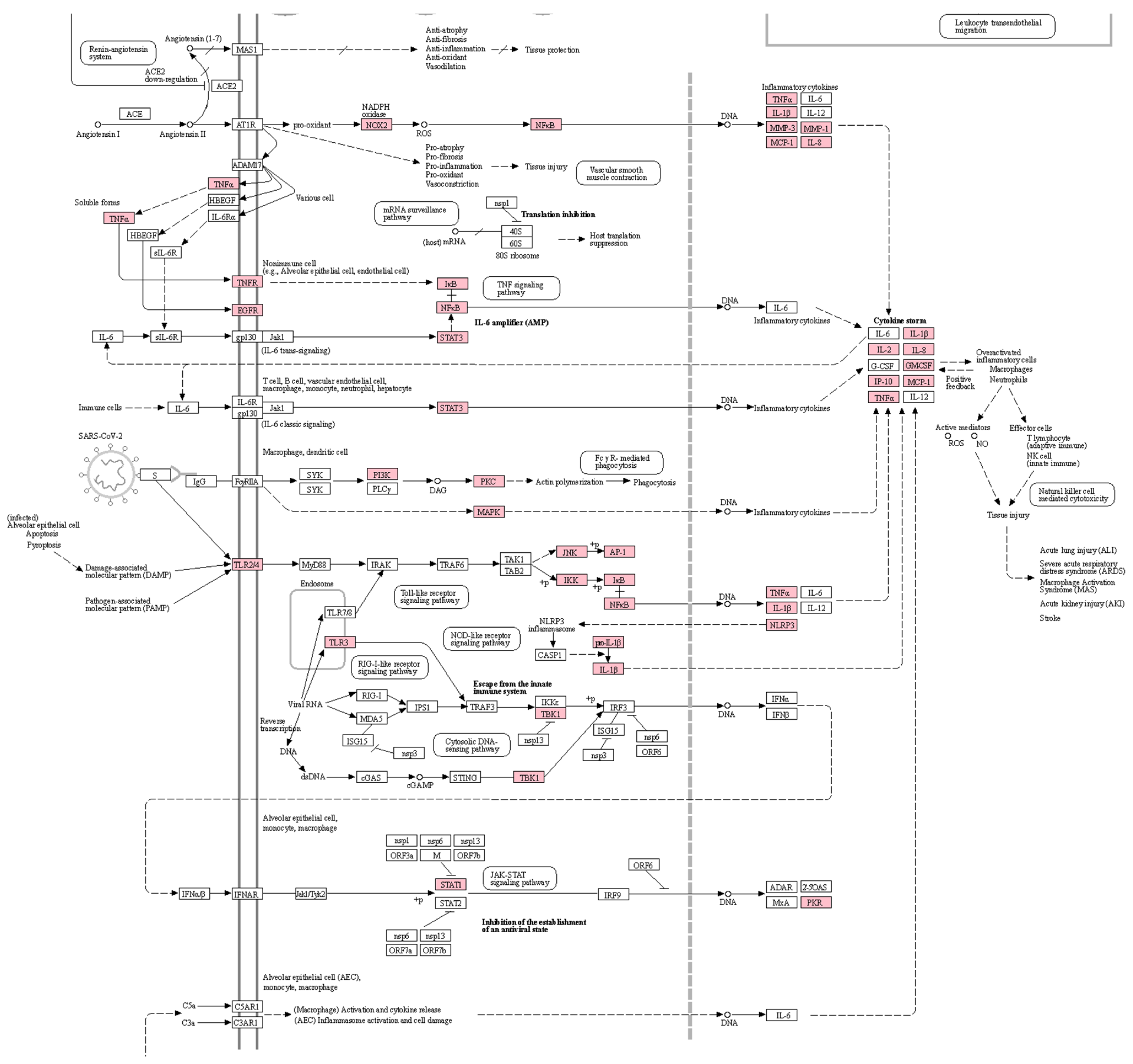
3.2. Enrichment Analysis for COVID-19-Related Tissues and Pathways
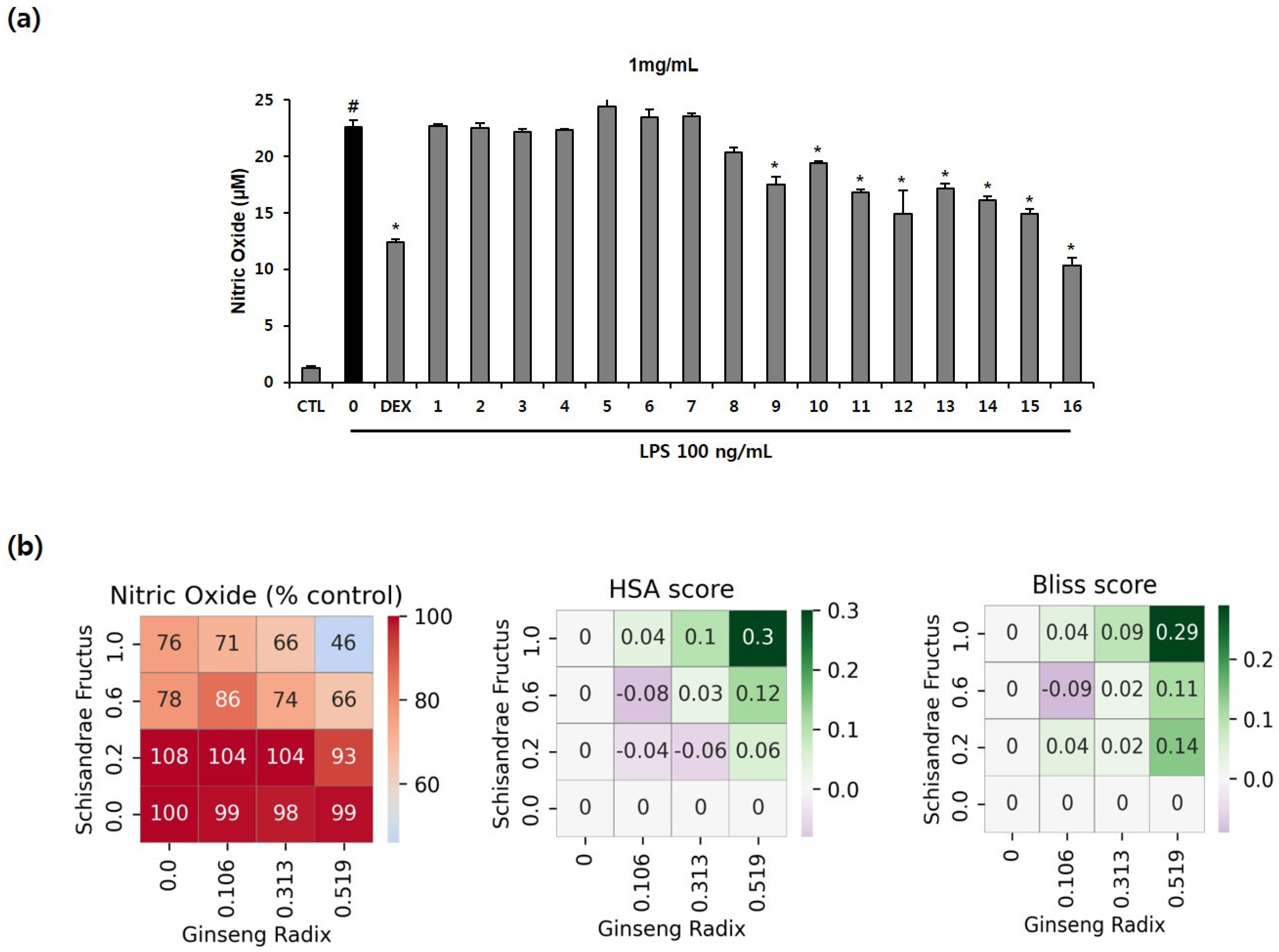
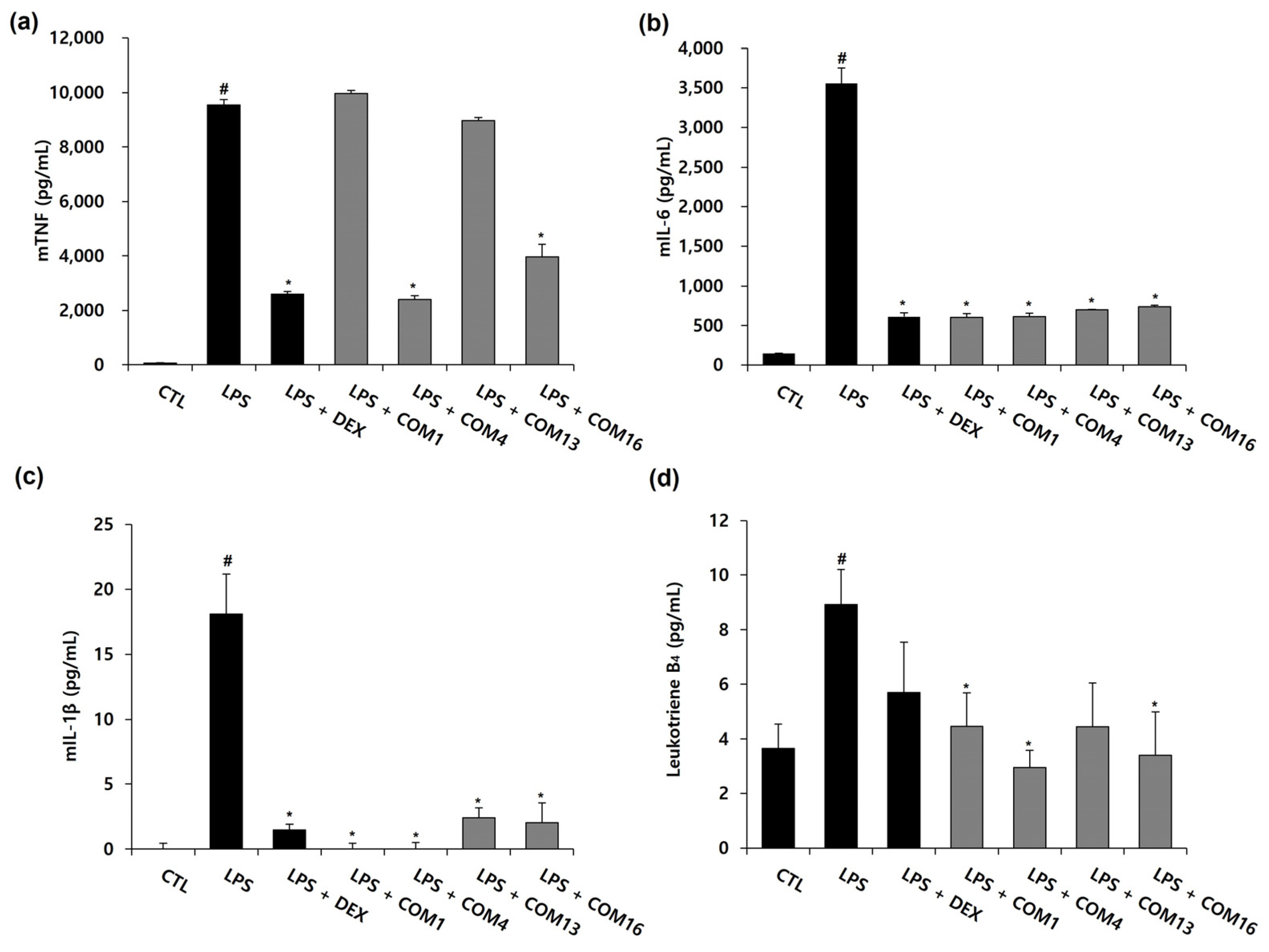
3.3. Effects of the Drug Combinations on LPS-Induced Inflammation in RAW264.7 Cells and Their Synergistic Score
3.4. Effects of the Drug Combinations on LPS-Mediated Pro-Inflammatory Cytokine Production
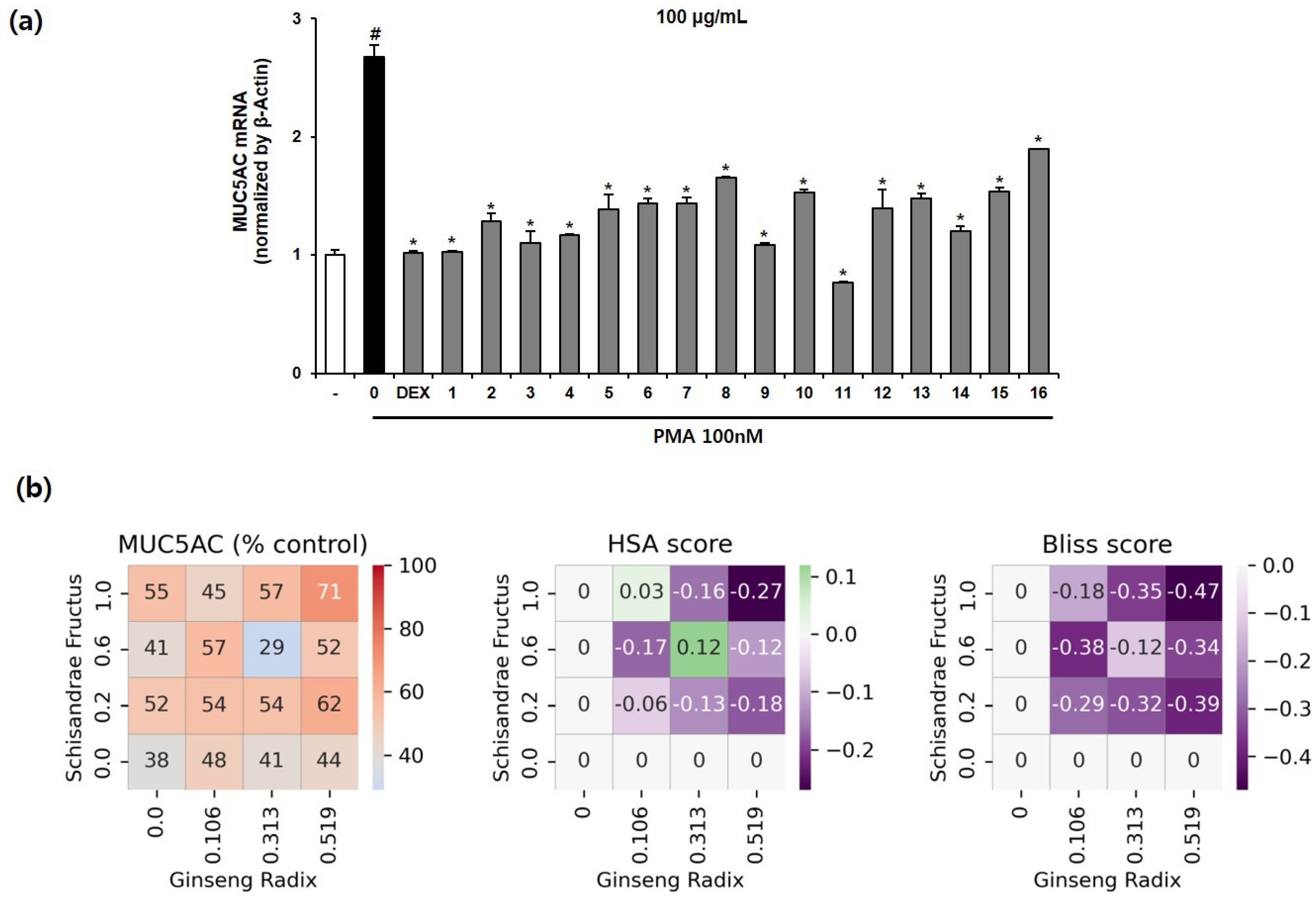
3.5. Effect of the Multi-Herb Ratio Combinations (MHRCs) on PMA-Induced MUC5AC mRNA Expression in NCI-H292 Cells
3.6. DS Analysis for Optimizing the Drug Combination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smale, S.T. Selective transcription in response to an inflammatory stimulus. Cell 2010, 140, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Pierce, G.F. Macrophages: Important physiologic and pathologic sources of polypeptide growth factors. Am. J. Respir. Cell Mol. Biol. 1990, 2, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Crooks, S.W.; Stockley, R.A. Leukotriene B4. Int. J. Biochem. Cell Biol. 1998, 30, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Martin, J.C. Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef]
- Mehandru, S.; Merad, M. Pathological sequelae of long-haul COVID. Nat. Immunol. 2022, 23, 194–202. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Bohnacker, S.; Hartung, F.; Henkel, F.; Quaranta, A.; Kolmert, J.; Priller, A.; Ud-Dean, M.; Giglberger, J.; Kugler, L.M.; Pechtold, L. Mild COVID-19 imprints a long-term inflammatory eicosanoid-and chemokine memory in monocyte-derived macrophages. Mucosal. Immunol. 2022, 15, 515–524. [Google Scholar] [CrossRef]
- Wiseman, N. Traditional Chinese medicine: A brief outline. J. Chem. Inf. Comput. Sci. 2002, 42, 445–455. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Z. Prevention and treatment of viral respiratory infections by traditional Chinese herbs. Chin. Med. J. 2014, 127, 1344–1350. [Google Scholar] [CrossRef]
- Huang, K.; Zhang, P.; Zhang, Z.; Youn, J.Y.; Wang, C.; Zhang, H.; Cai, H. Traditional Chinese Medicine (TCM) in the treatment of COVID-19 and other viral infections: Efficacies and mechanisms. Pharmacol. Ther. 2021, 225, 107843. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-I.; Shin, S.; Lee, N.; Lee, B.-J.; Lee, J.; Lee, H. A traditional herbal medication, Maekmoondong-tang, for cough: A systematic review and meta-analysis. J. Ethnopharmacol. 2016, 178, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.P.; Wang, L.; Wang, Z.; Xu, X.R.; Zhou, X.M.; Liu, G.; He, L.Y.; Wang, J.; Hsu, A.; Li, W.M.; et al. Chinese herbal medicine formula for acute asthma: A multi-center, randomized, double-blind, proof-of-concept trial. Respir. Med. 2018, 140, 42–49. [Google Scholar] [CrossRef]
- Liao, H.; Ye, J.; Gao, L.; Liu, Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomed. Pharmacother. 2021, 133, 110917. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wang, Y.; Shi, Y.; Yu, J.; Sun, W.; Zhang, Y. Effect of Traditional Chinese Medicine on Inflammatory Mediators in Pediatric Asthma. Mediat. Inflamm. 2016, 2016, 5143703. [Google Scholar] [CrossRef]
- Kubo, T.; Nishimura, H. Antipyretic effect of Mao-to, a Japanese herbal medicine, for treatment of type A influenza infection in children. Phytomedicine 2007, 14, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Fujikane, A.; Sakamoto, A.; Fujikane, R.; Nishi, A.; Ishino, Y.; Hiromatsu, K.; Nabeshima, S. Ephedrae Herba and Cinnamomi Cortex interactions with G glycoprotein inhibit respiratory syncytial virus infectivity. Commun. Biol. 2022, 5, 94. [Google Scholar] [CrossRef]
- Ma, J.X.; Xiao, X.; Zhou, K.F.; Huang, G.; Ao, B.; Zhang, Y.; Gao, W.J.; Lei, T.; Yang, L.; Fan, X.C.; et al. Herb pair of Ephedrae Herba-Armeniacae Semen Amarum alleviates airway injury in asthmatic rats. J. Ethnopharmacol. 2021, 269, 113745. [Google Scholar] [CrossRef]
- Zhong, S.; Bai, L.P.; Liu, X.D.; Cai, D.Y.; Yau, L.F.; Huang, C.Q.; Zhang, J.Q.; Lai, K.F.; Zhong, N.S. Cough Inhibition Activity of Schisandra chinensis in Guinea Pigs. J. Med. Food. 2021, 24, 348–357. [Google Scholar] [CrossRef]
- Chen, Y.; Kong, Y.; Wang, Q.; Chen, J.; Chen, H.; Xie, H.; Li, L. Schisandrin B Attenuates Airway Inflammation by Regulating the NF-κB/Nrf2 Signaling Pathway in Mouse Models of Asthma. J. Immunol. Res. 2021, 2021, 8029963. [Google Scholar] [CrossRef]
- Li, Y.H.; Zheng, F.J.; Huang, Y.; Zhong, X.G.; Guo, M.Z. Synergistic anti-inflammatory effect of Radix Platycodon in combination with herbs for cleaning-heat and detoxification and its mechanism. Chin. J. Integr. Med. 2013, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Y.; Yang, D.; Zhang, C.; Zhang, N.; Li, M.; Liu, Y. Platycodon grandiflorus—An ethnopharmacological, phytochemical and pharmacological review. J. Ethnopharmacol. 2015, 164, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Lee, H.J.; Park, S.H.; Kim, J.; Lee, D.; Lee, S.K.; Kim, Y.S.; Hong, J.H.; Seok, J.H.; Lee, C.J. Effects of the root of Platycodon grandiflorum on airway mucin hypersecretion in vivo and platycodin D(3) and deapi-platycodin on production and secretion of airway mucin in vitro. Phytomedicine 2014, 21, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Lee, S.-Y.; Jeon, B.-K.; Lee, J.-W.; Kim, Y.-S.; Lee, M.-N.; Lee, C.-J. Effect of platycodin D on airway MUC5AC mucin production and gene expression induced by growth factor and proinflammatory factor. Biomol. Ther. 2010, 18, 294–299. [Google Scholar] [CrossRef][Green Version]
- Phung, H.M.; Jang, D.; Trinh, T.A.; Lee, D.; Nguyen, Q.N.; Kim, C.-E.; Kang, K.S. Regulation of appetite-related neuropeptides by Panax ginseng: A novel approach for obesity treatment. J. Ginseng Res. 2022, 46, 609–619. [Google Scholar] [CrossRef]
- Li, H.; Cui, Q.K.; Li, Z.; Li, J.; Li, F. Clinical observation of the effect of modified Ginseng-Schisandra decoction (MGSD) on trace elements and immune function in children with spleen deficiency syndrome after recurrent respiratory tract infection (RRTI): A randomized controlled trial. Transl. Pediatr. 2021, 10, 1692–1700. [Google Scholar] [CrossRef]
- Meyer, C.T.; Wooten, D.J.; Lopez, C.F.; Quaranta, V. Charting the Fragmented Landscape of Drug Synergy. Trends Pharmacol. Sci. 2020, 41, 266–280. [Google Scholar] [CrossRef]
- Zhou, X.; Seto, S.W.; Chang, D.; Kiat, H.; Razmovski-Naumovski, V.; Chan, K.; Bensoussan, A. Synergistic Effects of Chinese Herbal Medicine: A Comprehensive Review of Methodology and Current Research. Front. Pharmacol. 2016, 7, 201. [Google Scholar] [CrossRef]
- Jang, D.; Lee, E.; Lee, S.; Kwon, Y.; Kang, K.S.; Kim, C.-E.; Kim, D. System-level investigation of anti-obesity effects and the potential pathways of Cordyceps militaris in ovariectomized rats. BMC Complement. Med. Ther. 2022, 22, 132. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, X.; Bai, H.; Ning, K. Network Pharmacology Databases for Traditional Chinese Medicine: Review and Assessment. Front. Pharmacol. 2019, 10, 123. [Google Scholar] [CrossRef]
- Zheng, W.J.; Yan, Q.; Ni, Y.S.; Zhan, S.F.; Yang, L.L.; Zhuang, H.F.; Liu, X.H.; Jiang, Y. Examining the effector mechanisms of Xuebijing injection on COVID-19 based on network pharmacology. BioData Min. 2020, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wang, X.; Liu, M.; Huang, M.; Shen, Z.; Feng, J.; Yang, H.; Li, Z.; Gao, J.; Ye, X. Exploring the treatment of COVID-19 with Yinqiao powder based on network pharmacology. Phytother. Res. 2021, 35, 2651–2664. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.D.; Xun, Y.; Lu, J.L.; Lu, Y.C.; Yang, Y.Y.; Zhou, P.; Hu, J.; Li, C.; Wang, S.G. Network pharmacology and molecular docking analyses on Lianhua Qingwen capsule indicate Akt1 is a potential target to treat and prevent COVID-19. Cell Prolif. 2020, 53, e12949. [Google Scholar] [CrossRef] [PubMed]
- Tao, Q.; Du, J.; Li, X.; Zeng, J.; Tan, B.; Xu, J.; Lin, W.; Chen, X.L. Network pharmacology and molecular docking analysis on molecular targets and mechanisms of Huashi Baidu formula in the treatment of COVID-19. Drug Dev. Ind. Pharm. 2020, 46, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-Y.; Lee, C.-Y.; Kim, Y.-S.; Kim, C.-E. The methodological trends of traditional herbal medicine employing network pharmacology. Biomolecules 2019, 9, 362. [Google Scholar] [CrossRef]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminformatics 2014, 6, 13. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef]
- Gaulton, A.; Hersey, A.; Nowotka, M.; Bento, A.P.; Chambers, J.; Mendez, D.; Mutowo, P.; Atkinson, F.; Bellis, L.J.; Cibrián-Uhalte, E. The ChEMBL database in 2017. Nucleic Acids Res. 2017, 45, D945–D954. [Google Scholar] [CrossRef]
- Ayers, M. ChemSpider: The free chemical database. Ref. Rev. 2012, 26, 45–46. [Google Scholar] [CrossRef]
- Bickerton, G.R.; Paolini, G.V.; Besnard, J.; Muresan, S.; Hopkins, A.L. Quantifying the chemical beauty of drugs. Nat. Chem. 2012, 4, 90–98. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Santos, A.; Von Mering, C.; Jensen, L.J.; Bork, P.; Kuhn, M. STITCH 5: Augmenting protein–chemical interaction networks with tissue and affinity data. Nucleic Acids Res. 2016, 44, D380–D384. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.; Szklarczyk, D.; Pletscher-Frankild, S.; Blicher, T.H.; von Mering, C.; Jensen, L.J.; Bork, P. STITCH 4: Integration of protein–chemical interactions with user data. Nucleic Acids Res. 2013, 42, D401–D407. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. Comparative toxicogenomics database (CTD): Update 2021. Nucleic Acids Res. 2021, 49, D1138–D1143. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Thul, P.J.; Lindskog, C. The human protein atlas: A spatial map of the human proteome. Protein Sci. 2018, 27, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Berenbaum, M.C. What is synergy? Pharmacol. Rev. 1989, 41, 93–141. [Google Scholar]
- Bliss, C.I. THE TOXICITY OF POISONS APPLIED JOINTLY1. Ann. Appl. Biol. 1939, 26, 585–615. [Google Scholar] [CrossRef]
- Hewson, C.A.; Edbrooke, M.R.; Johnston, S.L. PMA induces the MUC5AC respiratory mucin in human bronchial epithelial cells, via PKC, EGF/TGF-α, Ras/Raf, MEK, ERK and Sp1-dependent mechanisms. J. Mol. Biol. 2004, 344, 683–695. [Google Scholar] [CrossRef]
- Malik, P.; Patel, K.; Pinto, C.; Jaiswal, R.; Tirupathi, R.; Pillai, S.; Patel, U. Post-acute COVID-19 syndrome (PCS) and health-related quality of life (HRQoL)-A systematic review and meta-analysis. J. Med. Virol. 2022, 94, 253–262. [Google Scholar] [CrossRef]
- Song, W.J.; Hui, C.K.M.; Hull, J.H.; Birring, S.S.; McGarvey, L.; Mazzone, S.B.; Chung, K.F. Confronting COVID-19-associated cough and the post-COVID syndrome: Role of viral neurotropism, neuroinflammation, and neuroimmune responses. Lancet Respir. Med. 2021, 9, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Kang, K.; Lan, X.; Fei, D.; Wang, Q.; Li, X.; Chong, Y.; Gao, Y.; Wang, H.; Li, X.; et al. Cytokine levels in sputum, not serum, may be more helpful for indicating the damage in the lung and the prognosis of severe COVID-19—A case series. J. Infect. 2021, 83, e6–e9. [Google Scholar] [CrossRef] [PubMed]
- Jatakanon, A.; Lalloo, U.G.; Lim, S.; Chung, K.F.; Barnes, P.J. Increased neutrophils and cytokines, TNF-α and IL-8, in induced sputum of non-asthmatic patients with chronic dry cough. Thorax 1999, 54, 234. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.G.; Zlatic, K.; Scott, J.; Sewell, W.; Woolley, K.; Saltos, N. Chronic cough resembles asthma with IL-5 and granulocyte-macrophage colony-stimulating factor gene expression in bronchoalveolar cells. J. Allergy Clin. Immunol. 1998, 101, 320–326. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Ryter, S.W.; Kim, H.P.; Hoetzel, A.; Park, J.W.; Nakahira, K.; Wang, X.; Choi, A.M. Mechanisms of cell death in oxidative stress. Antioxid. Redox Signal. 2007, 9, 49–89. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Okuda, K.; Chen, G.; Subramani, D.B.; Wolf, M.; Gilmore, R.C.; Kato, T.; Radicioni, G.; Kesimer, M.; Chua, M.; Dang, H.; et al. Localization of Secretory Mucins MUC5AC and MUC5B in Normal/Healthy Human Airways. Am. J. Respir. Crit. Care Med. 2019, 199, 715–727. [Google Scholar] [CrossRef]
- Koeppen, M.; McNamee, E.N.; Brodsky, K.S.; Aherne, C.M.; Faigle, M.; Downey, G.P.; Colgan, S.P.; Evans, C.M.; Schwartz, D.A.; Eltzschig, H.K. Detrimental role of the airway mucin Muc5ac during ventilator-induced lung injury. Mucosal Immunol. 2013, 6, 762–775. [Google Scholar] [CrossRef]
- Johnson, L.; Shah, I.; Loh, A.X.; Vinall, L.E.; Teixeira, A.S.; Rousseau, K.; Holloway, J.W.; Hardy, R.; Swallow, D.M. MUC5AC and inflammatory mediators associated with respiratory outcomes in the British 1946 birth cohort. Respirology 2013, 18, 1003–1010. [Google Scholar] [CrossRef]
- Fonseca, S.G.; Romão, P.R.; Figueiredo, F.; Morais, R.H.; Lima, H.C.; Ferreira, S.H.; Cunha, F.Q. TNF-alpha mediates the induction of nitric oxide synthase in macrophages but not in neutrophils in experimental cutaneous leishmaniasis. Eur. J. Immunol. 2003, 33, 2297–2306. [Google Scholar] [CrossRef]
- Arias-Salvatierra, D.; Silbergeld, E.K.; Acosta-Saavedra, L.C.; Calderon-Aranda, E.S. Role of nitric oxide produced by iNOS through NF-κB pathway in migration of cerebellar granule neurons induced by Lipopolysaccharide. Cell Signal. 2011, 23, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, M.G.; Birchall, J.P.; Pearson, J.P. TNF-alpha in the regulation of MUC5AC secretion: Some aspects of cytokine-induced mucin hypersecretion on the in vitro model. Cytokine 2000, 12, 1732–1736. [Google Scholar] [CrossRef] [PubMed]
- Sikder, M.A.; Lee, H.J.; Mia, M.Z.; Park, S.H.; Ryu, J.; Kim, J.H.; Min, S.Y.; Hong, J.H.; Seok, J.H.; Lee, C.J. Inhibition of TNF-α-induced MUC5AC mucin gene expression and production by wogonin through the inactivation of NF-κB signaling in airway epithelial cells. Phytother. Res. 2014, 28, 62–68. [Google Scholar] [CrossRef]
- Kang, Y.S.; Han, M.H.; Hong, S.H.; Park, C.; Hwang, H.J.; Kim, B.W.; Kyoung, K.H.; Choi, Y.W.; Kim, C.M.; Choi, Y.H. Anti-inflammatory Effects of Schisandra chinensis (Turcz.) Baill Fruit Through the Inactivation of Nuclear Factor-κB and Mitogen-activated Protein Kinases Signaling Pathways in Lipopolysaccharide-stimulated Murine Macrophages. J. Cancer Prev. 2014, 19, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Kim, Y.K. New approach for herbal formula research: Network pharmacology. J. Physiol. Pathol. Korean Med. 2016, 30, 385–396. [Google Scholar] [CrossRef]
- Chung, K.; McCusker, M.; Page, C.; Dent, G.; Guinot, P.; Barnes, P. Effect of a ginkgolide mixture (BN 52063) in antagonising skin and platelet responses to platelet activating factor in man. Lancet 1987, 329, 248–251. [Google Scholar] [CrossRef]
- Han, Y. Synergic anticandidal effect of epigallocatechin-O-gallate combined with amphotericin B in a murine model of disseminated candidiasis and its anticandidal mechanism. Biol. Pharm. Bull. 2007, 30, 1693–1696. [Google Scholar] [CrossRef]
- Chen, L.; Sun, X.; Wang, Q.; Wu, W.; Tian, H.; Liu, L.; Jiang, Y.; Bei, C.; Chen, N.; Chen, J. Potential synergy mechanism of processing methods for the basic remedies of Qixue Shuangbu prescription based on integrated metabolomics strategy and network pharmacology study. J. Chromatogr. Sci. 2021, 59, 983–993. [Google Scholar] [CrossRef]
- Zhao, N.; Kong, H.; Liu, H.; Shi, Q.; Qi, X.; Chen, Q. A network pharmacology approach to evaluate the synergistic effect of dihydromyricetin and myricitrin in vine tea on the proliferation of B16F10 cells. Front. Nutr. 2022, 9, 993133. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EH (g) | PR (g) | SF (g) | GR (g) | |
|---|---|---|---|---|
| 1 | 0.273 | 0.263 | 0.000 | 0.000 |
| 2 | 0.273 | 0.263 | 0.106 | 0.000 |
| 3 | 0.273 | 0.263 | 0.313 | 0.000 |
| 4 | 0.273 | 0.263 | 0.519 | 0.000 |
| 5 | 0.273 | 0.263 | 0.000 | 0.200 |
| 6 | 0.273 | 0.263 | 0.106 | 0.200 |
| 7 | 0.273 | 0.263 | 0.313 | 0.200 |
| 8 | 0.273 | 0.263 | 0.519 | 0.200 |
| 9 | 0.273 | 0.263 | 0.000 | 0.600 |
| 10 | 0.273 | 0.263 | 0.106 | 0.600 |
| 11 | 0.273 | 0.263 | 0.313 | 0.600 |
| 12 | 0.273 | 0.263 | 0.519 | 0.600 |
| 13 | 0.273 | 0.263 | 0.000 | 1.000 |
| 14 | 0.273 | 0.263 | 0.106 | 1.000 |
| 15 | 0.273 | 0.263 | 0.313 | 1.000 |
| 16 | 0.273 | 0.263 | 0.519 | 1.000 |
| Gene | Sense Primer Sequence (5′-3′) | Antisense Primer Sequence (5′-3′) |
|---|---|---|
| MUC5AC | TCCACCATATACCGCCACAGA | TGGACGGACAGTCACTGTCAAC |
| β-actin | AGGAGAAGCTGTGCTACGTC | GGATGTCCACGTCACACTTC |
| Database | Drug | Overlap | p-Value (Adjusted) | Odds Ratio | Combined Score |
|---|---|---|---|---|---|
| CTD database | Drug combination | 11/37 | 5.72 × 10−5 | 6.33 | 72.02 |
| Ephedrae Herba | 9/37 | 0.00142 | 5.02 | 41.00 | |
| Platycodonis Radix | 7/37 | 4.37 × 10−4 | 7.80 | 72.90 | |
| Ginseng Radix | 8/37 | 5.81 × 10−5 | 9.01 | 102.38 | |
| Schisandrae Fructus | 0/37 | - | - | - | |
| KEGG | Drug combination | 37/232 | 8.11 × 10−12 | 4.47 | 119.04 |
| Ephedrae Herba | 35/232 | 4.64 × 10−11 | 4.37 | 108.57 | |
| Platycodonis Radix | 20/232 | 1.14 × 10−7 | 4.92 | 84.43 | |
| Ginseng Radix | 20/232 | 1.61 × 10−7 | 4.79 | 80.20 | |
| Schisandrae Fructus | 1/232 | 0.24 | 4.07 | 6.05 |
| Tissue | Drug | Overlap | p-Value (Adjusted) | Odds Ratio | Combined Score |
|---|---|---|---|---|---|
| Bronchial epithelial cells | Drug combination | 30/280 | 5.67 × 10−5 | 2.79 | 35.73 |
| Ephedrae Herba | 30/280 | 2.40 × 10−5 | 2.93 | 39.92 | |
| Platycodonis Radix | 9/280 | 0.45 | 1.68 | 3.84 | |
| Ginseng Radix | 11/280 | 0.26 | 2.02 | 7.32 | |
| Schisandrae Fructus | 1/280 | 0.40 | 3.36 | 4.44 | |
| Lung | Drug combination | 22/299 | 0.047 | 1.83 | 8.78 |
| Ephedrae Herba | 21/299 | 0.063 | 1.82 | 8.35 | |
| Platycodonis Radix | 12/299 | 0.13 | 2.13 | 8.87 | |
| Ginseng Radix | 11/299 | 0.28 | 1.89 | 6.07 | |
| Schisandrae Fructus | 0/299 | - | - | - |
| Pathway | Drug | Overlap | p-Value (Adjusted) | Odds Ratio | Combined Score |
|---|---|---|---|---|---|
|
TNF signaling pathway | Drug combination | 36/112 | 2.68 × 10−21 | 11.21 | 560.76 |
| Ephedrae Herba | 31/112 | 4.54 × 10−17 | 9.43 | 373.21 | |
| Platycodonis Radix | 20/112 | 8.17 × 10−13 | 11.40 | 348.36 | |
| Ginseng Radix | 20/112 | 1.12 × 10−12 | 11.10 | 333.83 | |
| Schisandrae Fructus | 0/112 | - | - | - | |
|
NF-κB signaling pathway | Drug combination | 23/104 | 1.13 × 10−10 | 6.61 | 157.46 |
| Ephedrae Herba | 21/104 | 2.05 × 10−9 | 6.15 | 128.27 | |
| Platycodonis Radix | 9/104 | 3.69 × 10−4 | 4.83 | 40.93 | |
| Ginseng Radix | 11/104 | 1.76 × 10−5 | 5.90 | 69.33 | |
| Schisandrae Fructus | 0/104 | - | - | - | |
| Calcium signaling pathway | Drug combination | 66/240 | 7.96 × 10−34 | 9.28 | 743.30 |
| Ephedrae Herba | 66/240 | 7.03 × 10−35 | 9.74 | 805.21 | |
| Platycodonis Radix | 37/240 | 5.74 × 10−21 | 9.96 | 500.62 | |
| Ginseng Radix | 38/240 | 1.55 × 10−21 | 10.02 | 517.97 | |
| Schisandrae Fructus | 1/240 | 0.25 | 3.93 | 5.72 |
| Drug | Compounds |
|---|---|
| Ephedrae Herba | hexanoic acid, carvacrol, pentadecanoic acid, tricin, ethyl acetate, cinnamaldehyde, tetradecanoic acid, guaiazulene, ethanol, diethyl sulfate, p-cresol, dibutyl phthalate, apigenin, gamma-butyrolactone, safranal, 2,3,5,6-tetramethylpyrazine, pseudoephedrine, farnesol, palmitic acid, quercetin, nonanoic acid, kaempferol, lauric acid, decanoic acid, thymol, styrene, hexanal, phenanthrene, toluene, ephedrine |
| Platycodonis Radix | palmitic acid, luteolin, 2-cyclopenten-1-one, acacetin |
| Ginseng Radix | Stigmasterol, paeonol, protopanaxatriol, pentadecanoic acid, palmitic acid, octane, kaempferol, beta-elemene, 6-methoxyluteolin, beta-sitosterol, dibutyl phthalate, (20s)-protopanaxatriol |
| Schisandrae Fructus | dibutyl phthalate |
| Responses | Target | Upper Limit | Criteria for Setting the Upper Limit |
|---|---|---|---|
| Nitric Oxide | Minimum | 18.73 | 10% improvement compared to LPS |
| TNF-α | Minimum | 6682.51 | 30% improvement compared to LPS |
| IL-6 | Minimum | 1777.42 | 50% improvement compared to LPS |
| IL-1β | Minimum | 9.06 | 50% improvement compared to LPS |
| LTB4 | Minimum | 4.47 | 50% improvement compared to LPS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, D.; Lee, M.J.; Kim, K.S.; Kim, C.-E.; Jung, J.H.; Cho, M.; Hong, B.-H.; Park, S.J.; Kang, K.S. Network Pharmacological Analysis on the Herbal Combinations for Mitigating Inflammation in Respiratory Tracts and Experimental Evaluation. Healthcare 2023, 11, 143. https://doi.org/10.3390/healthcare11010143
Jang D, Lee MJ, Kim KS, Kim C-E, Jung JH, Cho M, Hong B-H, Park SJ, Kang KS. Network Pharmacological Analysis on the Herbal Combinations for Mitigating Inflammation in Respiratory Tracts and Experimental Evaluation. Healthcare. 2023; 11(1):143. https://doi.org/10.3390/healthcare11010143
Chicago/Turabian StyleJang, Dongyeop, Myong Jin Lee, Kang Sub Kim, Chang-Eop Kim, Jong Ho Jung, Minkwan Cho, Bo-Hee Hong, Shin Jung Park, and Ki Sung Kang. 2023. "Network Pharmacological Analysis on the Herbal Combinations for Mitigating Inflammation in Respiratory Tracts and Experimental Evaluation" Healthcare 11, no. 1: 143. https://doi.org/10.3390/healthcare11010143
APA StyleJang, D., Lee, M. J., Kim, K. S., Kim, C.-E., Jung, J. H., Cho, M., Hong, B.-H., Park, S. J., & Kang, K. S. (2023). Network Pharmacological Analysis on the Herbal Combinations for Mitigating Inflammation in Respiratory Tracts and Experimental Evaluation. Healthcare, 11(1), 143. https://doi.org/10.3390/healthcare11010143