
Phi-Bonacci Butterfly Stroke Numbers to Assess Self-Similarity in Elite Swimmers

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
- A stance (swing) duration, (), being close to 62% of gait cycle duration ;
- A swing (stance) duration being close to 38% of gait cycle duration;
- A double support (double float) duration, (), being consequently close to 24% of gait cycle duration.
2. Generalized Fibonacci Sequences in Butterfly Stroke
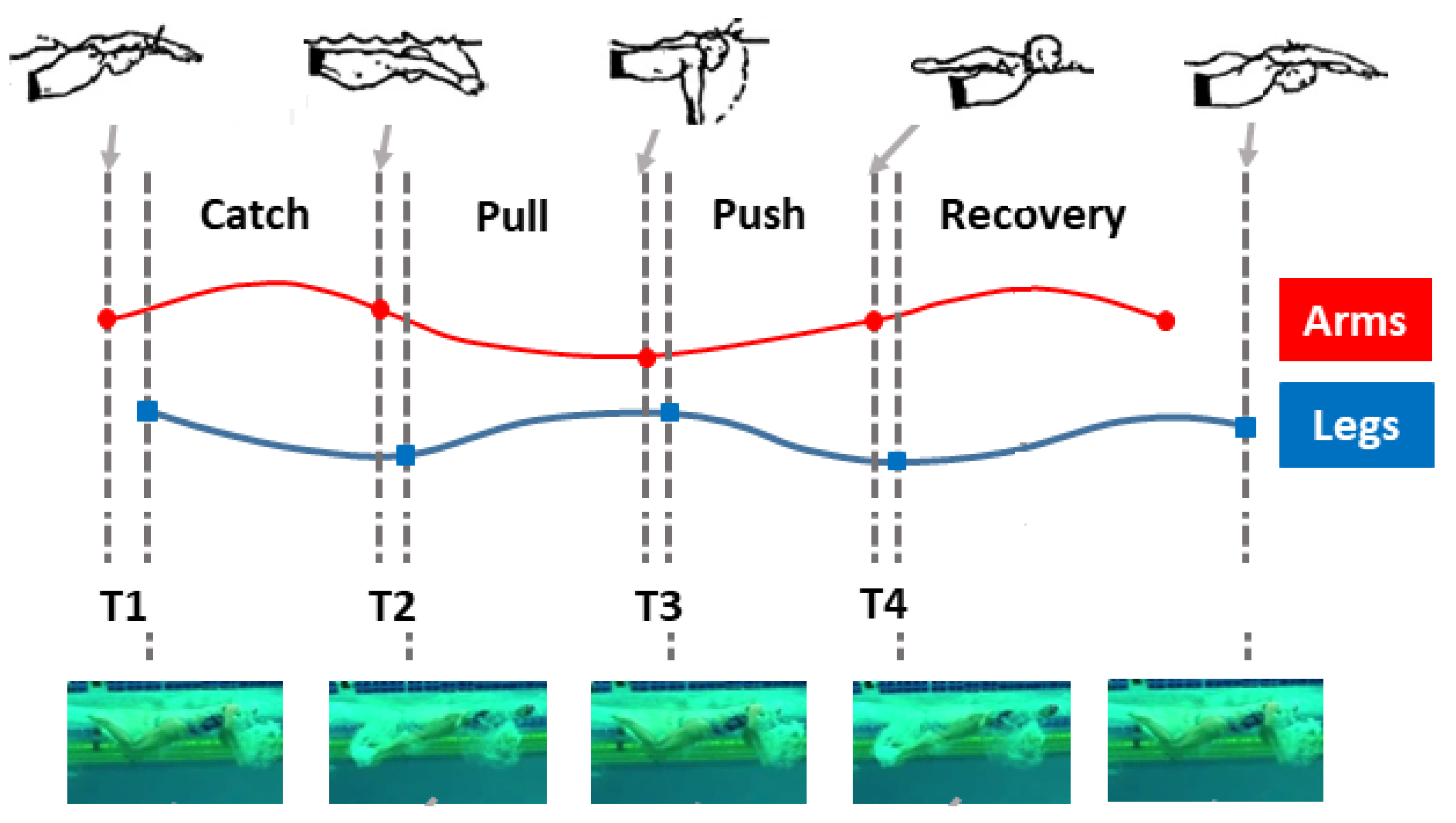
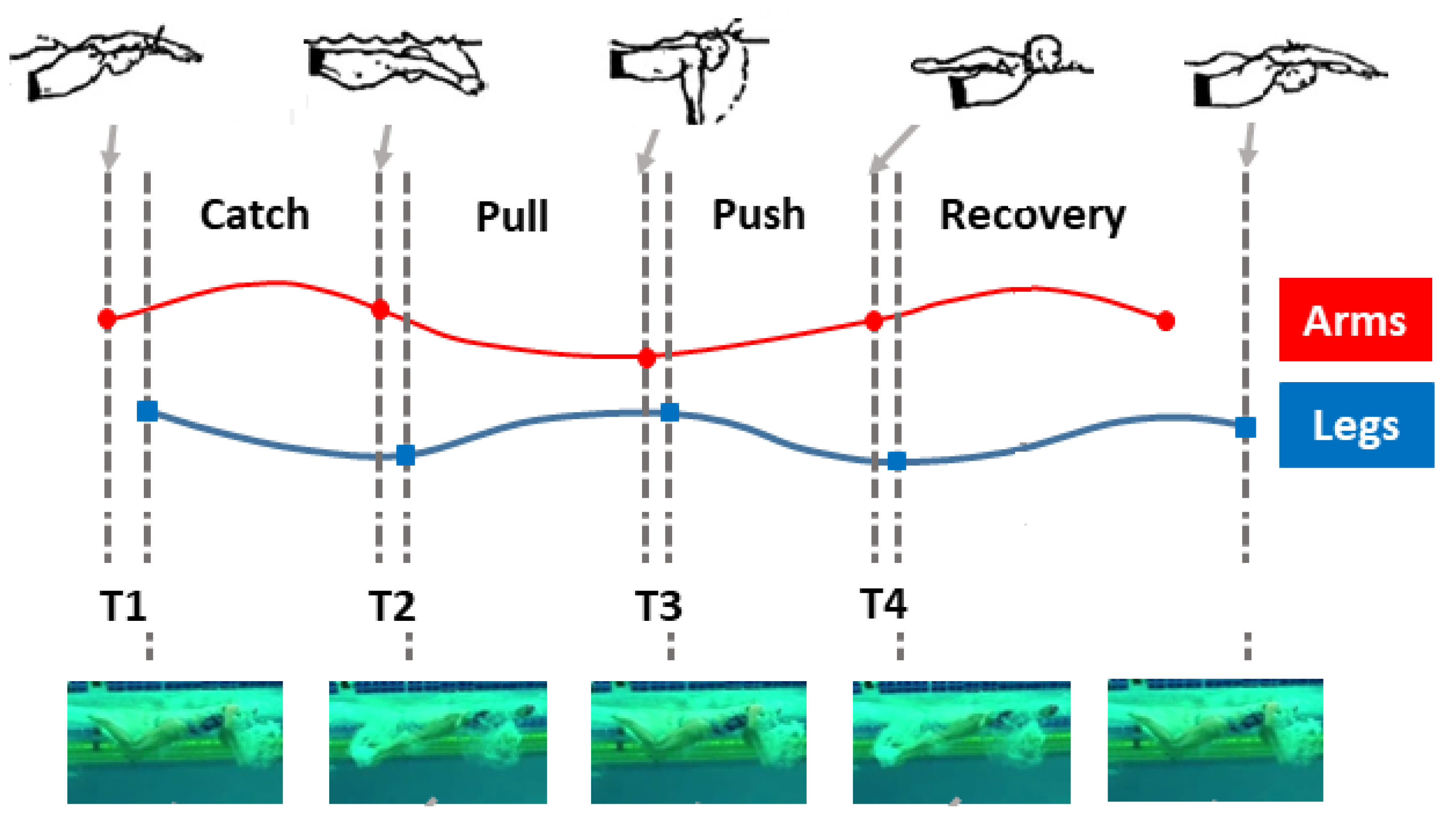
2.1. Butterfly Stroke Phases
- (i)
- Entry and catch phase (between the entry of the hands into the water and the beginning of their backward movement), with duration ;
- (ii)
- Pull phase (between the beginning of the backward movement of the hands and their entry into the plane vertical to the shoulders), with duration ;
- (iii)
- Push phase (between the positioning of the hands below the shoulders and their exit from the water), with duration ;
- (iv)
- Recovery phase (between the arrival of the hands at the water level and their subsequent entry into the water), with duration ,
- (i)
- Downward phase 1 (between the high and low break-even points– at first occurrence–of the feet during the first undulation), with duration ;
- (ii)
- Upward phase 1 (between the low and high break-even points–at first occurrence–of the feet during the first undulation), with duration ;
- (iii)
- Downward phase 2 (between the high and low break-even points–at first occurrence–of the feet during the second undulation), with duration ;
- (iv)
- Upward phase 2 (between the low and high break-even points–at first occurrence–of the feet during the second undulation), with duration .
- , between the start of Entry and catch phase and the start of Downward phase 1;
- , between the start of Pull phase and the start of Upward phase 1;
- , between the start of Push phase, and the start of Downward phase 2;
- , between the start of Recovery phase, and the start of Upward phase 2.
2.2. Self-Similarity and Enhanced Self-Similarity in Butterfly Stroke
2.3. Quantitative Measures of Self-Similarity and Enhanced Self-Similarities
3. Experimental Analysis
3.1. Phase Durations and Interlimb Coordination
3.2. Self-Similarity Analysis
- Rather small values are obtained for IL1–IL7, with IL1’s one being the smallest, owing to the strict closeness of the corresponding phase percentage values to , , , , and ;
- Relatively large reductions in the self-similarity and enhanced self-similarity magnitudes (especially of the latter) appear for the national-level swimmers NL1–NL2 when compared to the international-level swimmers IL1–IL7;
- The - and - values turn out to reproduce the order of physical shape within the two swimmers’ set;
- IL5 even presents an -value () that is close to the one () characterizing the strongly enhanced self-similar structure of Remark ();
- The slight differences in the phase durations of Table 1 and Table 2 (compare, for instance, IL2 to NL1, or IL4 to IL5, or IL2 to IL3), which lead to the differences in self-similarity magnitudes of Table 5 and Table 6, have been successfully identified via the high frame rate analysis used in this paper, with the self-similarity information complementing the delay partition values of Table 3 and Table 4;
- Larger percentage reductions in enhanced self-similarity (with respect to self-similarity) are exhibited by IL2, IL3, IL5—when compared to IL1, IL4, IL5, IL6, NL1, NL2—so that the - values for IL2–IL3 and IL5 tend to thicken (more than the -ones) towards the - values for IL4 and IL6, respectively.
4. Discussion
- Theorem 1 and Theorem 2 provide partition constraints that regard the leg stroke, with the inter-delay composition linking the arm phase partition with the leg phase partition;
- The durations and play the role of and in the front crawl stroke in [6] and of the right and left swing (right and left stance) durations in the asymmetric walking (running) of [2,5], with the involved equality between the durations of such phases simply transposing the swing (stance)-symmetry constraint of symmetric walking (running);
- In light of the twelve-tone equal temperament-based interpretation, constraint imposes that the stroke duration of an -kick-to-kick temporal symmetric repetitive butterfly stroke equals the sum of the durations of eight disjoint sub-phases, three among them with duration and five among them with duration : according to [6], notes D4, E4, G4, C5 correspond, in the (suspended and restored) C-variant Cadd2-chord, to the frequencies: Hz, Hz, Hz, Hz ( Hz), with corresponding to elements of a Fibonacci sequence and with the ratios , , of consecutive elements of such sequence quickly getting close to ;
- The above results—again occurring in elite swimmers—confirm that, in contrast to walking, a precise swimming technique (coming from a considerable amount of practice and instruction repeating specifically precise movements for a long enough time, while producing rhythmic motor patterns through the interrelationship between cortical input, central pattern generator (CPG), and sensory feedback) is relevantly involved (the intra-cyclic variation of the horizontal velocity of the hip is larger in non-expert swimmers than their expert counterparts [20]) as recalled by [6], the fractal dimension is – for highly qualified expert swimmers [20], whereas it is – for on-land walking [21].
5. Practical Implications
6. Strengths and Limits
7. Forecast
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Wang, X. The power sums involving fibonacci polynomials and their applications. Symmetry 2019, 11, 635. [Google Scholar] [CrossRef] [Green Version]
- Verrelli, C.M.; Iosa, M.; Roselli, P.; Pisani, A.; Giannini, F.; Saggio, G. Generalized finite-length Fibonacci sequences in healthy and pathological human walking: Gait recursivity, asymmetry, consistency, self-similarity, and variability. In Frontiers in Human Neuroscience; Special Issue: Rhythmic Patterns in Neuroscience and Human Physiology; to be published.
- Horadam, A.F. A generalized Fibonacci sequence. Am. Math. Mon. 1961, 68, 455–459. [Google Scholar] [CrossRef]
- Iosa, M.; Fusco, A.; Marchetti, F.; Morone, G.; Caltagirone, C.; Paolucci, S.; Peppe, A. The golden ratio of gait harmony: Repetitive proportions of repetitive gait phases. Biomed Res. Int. 2013, 2013, 918642. [Google Scholar] [CrossRef] [Green Version]
- Marino, R.; Verrelli, C.M.; Gnucci, M. Synchronicity rectangle for temporal gait analysis: Application to Parkinson’s Disease. Biomed. Signal Process. Control 2020, 62, 102156. [Google Scholar] [CrossRef]
- Verrelli, C.M.; Romagnoli, C.; Jackson, R.R.; Ferretti, I.; Annino, G.; Bonaiuto, V. Front crawl stroke in swimming: Ratios of phase durations and self-similarity. J. Biomech. 2021, 118, 110267. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.K.; Nudds, R.L.; Thomas, A.L.R. Flying and swimming animals cruise at a Strouhal numer tuned for high power efficiency. Lett. Nat. 2003, 425, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Eloy, C. Optimal Strouhal number for swimming animals. J. Fluids Struct. 2012, 30, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, T.M.; Keskinen, K.L.; Fernandes, R.; Colaço, P.; Lima, A.B.; Vilas-Boas, J.P. Energy cost and intracyclic variation of the velocity of the centre of mass in butterfly stroke. Eur. J. Appl. Physiol. 2005, 93, 519–523. [Google Scholar] [CrossRef]
- Seifert, L.; Boulesteix, L.; Chollet, D.; Vilas-Boas, J.P. Differences in spatial-temporal parameters and arm–leg coordination in butterfly stroke as a function of race pace, skill and gender. Hum. Mov. Sci. 2008, 27, 96–111. [Google Scholar] [CrossRef]
- Di Prampero, P.E.; Pendergast, D.R.; Wilson, D.W.; Rennie, D.W. Blood Lactic Acid Concentrations in High Velocity Swimming. In Swimming Medicine IV, Proceedings of the Fourth International Congress on Swimming Medicine, Stockholm, Sweden; Eriksson, B., Furberg, B., Eds.; University Park Press: Baltimore, MD, USA, 1978; pp. 249–261. [Google Scholar]
- Chollet, D.; Seifert, L.; Boulesteix, L.; Carter, M. Arm–leg coordination in elite butterfly swimmers. Int. J. Sport Med. 2006, 27, 322–329. [Google Scholar] [CrossRef]
- Seifert, L.; Delignieres, D.; Chollet, D. Effect of expertise on butterfly stroke coordination. J. Sport. Sci. 2007, 25, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Lanotte, N.; Annino, G.; Bifaretti, S.; Gatta, G.; Romagnoli, C.; Salvucci, A.; Bonaiuto, V. A new device for propulsion analysis in swimming. Proceedings 2018, 2, 285. [Google Scholar] [CrossRef] [Green Version]
- Tourny-Chollet, C.; Chollet, D.; Hogie, S.; Papparodopoulos, C. Kinematic analysis of butterfly turns of international and national swimmers. J. Sport. Sci. 2002, 20, 383–390. [Google Scholar] [CrossRef]
- Mason, B.R.; Tong, Z.; Richards, R.J. A biomechanical analysis of the butterfly stroke. Excel 1991, 7, 6–11. [Google Scholar]
- Chollet, D.; Chalies, S.; Chatard, J.C. A new index of coordination for the crawl: Description and usefulness. Int. J. Sport. Med. 1999, 21, 54–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungerechts, B.E. A comparison of the movements of the rear parts of dolphins and butterfly swimmers. In Biomechanics and Medicine in Swimming; Hollander, A.P., Huijing, P.A., de Groot, G., Eds.; Human Kinetics: Champaign, IL, USA, 1983; pp. 215–221. [Google Scholar]
- Sanders, R.H.; Cappaert, J.M.; Devlin, R.K. Wave Characteristics of butterfly swimming. J. Biomech. 1995, 28, 9–16. [Google Scholar] [CrossRef]
- Barbosa, T.M.; Goh, W.X.; Morais, J.E.; Costa, M.J. Variation of linear and nonlinear parameters in the swim strokes according to the level of expertise. Mot. Control 2017, 21, 312–326. [Google Scholar] [CrossRef]
- Schiffman, J.M.; Chelidze, D.; Adams, A.; Segala, D.B.; Hasselquist, L. Nonlinear analysis of gait kinematics to track changes in oxygen consumption in prolonged load carriage walking: A pilot study. J. Biomech. 2009, 42, 2196–2199. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, T.M.; Goh, W.X.; Morais, J.E.; Costa, M.J. Comparison of classical kinematics, entropy, and fractal properties as measures of complexity of the motor system in swimming. Front. Psychol. 2016, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Takagi, H.; Sugimoto, S.; Nishijima, N.; Wilson, B. Differences in stroke phases, arm-leg coordination and velocity fluctuation due to event, gender and performance level in breaststroke. Sport. Biomech. 2004, 3, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Ualí, I.; Herrero, A.J.; Garatachea, N.; Marín, P.J.; Alvear-Ordenes, I.; García-López, D. Maximal strength on different resistance training rowing exercises predicts start phase performance in elite kayakers. J. Strenght Cond. Res. 2012, 26, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Landlinger, J.; Lindinger, S.; Stöggl, T.; Wagner, H.; Müller, E. Key factors and timing patterns in the tennis forehand of different skill levels. J. Sport. Sci. Med. 2010, 9, 643–651. [Google Scholar]

{kind=link}
| IL1 | |||||
| IL2 | |||||
| IL3 | |||||
| IL4 | |||||
| IL5 | |||||
| IL6 | |||||
| IL7 | |||||
| NL1 | |||||
| NL2 |
| IL1 | |||||
| IL2 | |||||
| IL3 | |||||
| IL4 | |||||
| IL5 | |||||
| IL6 | |||||
| IL7 | |||||
| NL1 | |||||
| NL2 |
| NTD | |||||
|---|---|---|---|---|---|
| IL1 | |||||
| IL2 | |||||
| IL3 | |||||
| IL4 | |||||
| IL5 | |||||
| IL6 | |||||
| IL7 | |||||
| NL1 | |||||
| NL2 |
| a | b | c | d | |
|---|---|---|---|---|
| IL1 | ||||
| IL2 | ||||
| IL3 | ||||
| IL4 | ||||
| IL5 | ||||
| IL6 | ||||
| IL7 | ||||
| NL1 | ||||
| NL2 |
| IL1 | |||||
| IL2 | |||||
| IL3 | |||||
| IL4 | |||||
| IL5 | |||||
| IL6 | |||||
| IL7 | |||||
| NL1 | |||||
| NL2 |
| IL1 | ||
| IL2 | ||
| IL3 | ||
| IL4 | ||
| IL5 | ||
| IL6 | ||
| IL7 | ||
| NL1 | ||
| NL2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verrelli, C.M.; Romagnoli, C.; Jackson, R.; Ferretti, I.; Annino, G.; Bonaiuto, V. Phi-Bonacci Butterfly Stroke Numbers to Assess Self-Similarity in Elite Swimmers. Mathematics 2021, 9, 1545. https://doi.org/10.3390/math9131545
Verrelli CM, Romagnoli C, Jackson R, Ferretti I, Annino G, Bonaiuto V. Phi-Bonacci Butterfly Stroke Numbers to Assess Self-Similarity in Elite Swimmers. Mathematics. 2021; 9(13):1545. https://doi.org/10.3390/math9131545
Chicago/Turabian StyleVerrelli, Cristiano Maria, Cristian Romagnoli, Roxanne Jackson, Ivo Ferretti, Giuseppe Annino, and Vincenzo Bonaiuto. 2021. "Phi-Bonacci Butterfly Stroke Numbers to Assess Self-Similarity in Elite Swimmers" Mathematics 9, no. 13: 1545. https://doi.org/10.3390/math9131545
APA StyleVerrelli, C. M., Romagnoli, C., Jackson, R., Ferretti, I., Annino, G., & Bonaiuto, V. (2021). Phi-Bonacci Butterfly Stroke Numbers to Assess Self-Similarity in Elite Swimmers. Mathematics, 9(13), 1545. https://doi.org/10.3390/math9131545