1. Introduction
Estimating non-measurable connectivity parameters from experimental measurements is an important challenge to understand brain function. Brain activity is mainly driven by the intrinsic dynamics of neurons and their mutual connections, which are essentially regulated by synapses. A thorough knowledge of individual membrane potentials and synaptic terms is, therefore, crucial to understand how the brain operates. However, we are far from this utopia: current recording methods can only retrieve partial information, such as the time evolution of membrane potential of one or a few neurons. Unfortunately, other quantities that shape the connectivity, like synaptic conductances, are impossible to be directly recorded in experiments.
The time course of synaptic conductances arriving at a single cell not only provides local information about its functional connectivity, but it often constitutes an indicator to validate more general theories such as obtaining information about the architecture of the cortex, discerning between phase insensitive or spatial phase selective coupling [
1], unveiling whether the integrative properties of cortical neurons are altered under high-conductance states [
2], as well as studying many neuronal problems related to the balance between excitation and inhibition [
3]. Unfortunately, there are no direct methods to estimate synaptic conductances, and thus they have to be deduced from experimentally observable quantities. As no direct methods are known to estimate synaptic conductances, they have to be deduced from experimentally observable quantities. This leads to the need of using inverse mathematical methods. As for many inverse problems, it has been proved to be an arduous analytical and computational task for which no general procedures are yet available.
Different strategies have been developed to estimate the time course of the synaptic conductances from time series of the membrane potential. Most strategies are based on deterministic linear estimation procedures (see [
3,
4,
5,
6,
7]). However, researchers are aware of possible misestimations coming from the measurement noise produced during recordings. Therefore, new methods have been also devised to solve this problem from different approaches: stochastic linear procedures (see [
8,
9,
10,
11,
12,
13]), sophisticated filtering techniques (see [
14,
15,
16,
17,
18]), and techniques using the
relation extracted experimentally (see [
19]).
Even though all these strategies provide circumstantial solutions, it is also well known that they can only be applied in the subthreshold regime, where ionic currents are negligible (see [
20,
21]), a situation for which simplified models (close to linear) are representative enough to account for the voltage dynamics. However, it has also been demonstrated that these linear procedures can also provide wrong estimations when ionic currents are active in the subthreshold regime causing non-spikes in the membrane potential of the postsynaptic neuron. This fact motivates the introduction of new strategies to estimate conductances under these nonlinear paradigms (see [
22,
23]). More recently, in [
24], the authors identify that another artifact that can provide wrong estimations is to assume that the synaptic conductances are voltage-independent; to avoid this problem, they provide a new estimation procedure, also in the linear framework. Despite the many efforts in the literature to get rid of artifacts that can contaminate the estimation of conductances (influence of noise, activity of subthreshold ionic currents, variability across trials, etc.), one of the most resilient troubles are the nonlinearities that the
curve exhibits in the presence of spiking mechanisms.
On the other hand, a line of research is devoted to look for possible strategies to estimate conductances in the spiking regime. For instance, in [
25], the authors use a firing clamp technique where a description of the spike is given in terms of the membrane potential threshold and the amplitude of the spike. These traces are approximated by calibrating the membrane potential using dynamic clamp in in vitro experiments of isolated neurons and a quadratic polynomial approximation. More recently, in [
26] we introduce a mathematical approach to estimate conductances by characterizing the period of the membrane potential, and in [
27] estimations of the conductances of spike trains of the membrane potential are performed.
In [
26], we present a first proof-of-concept to estimate synaptic conductances in the spiking regime based on information about the
curve of the neuron. The idea was exemplified with the McKean model, a piecewise linear system that allows for a quasi-analytic expression of the
curve. More specifically, we approximated the firing period,
T, as a function of the parameters of the system and, in particular, of the synaptic conductance parameter,
. Therefore, for a given interspike interval (ISI) of length
, one can numerically solve the implicit equation
to obtain an estimation,
, of the synaptic conductances during that ISI. In order to obtain the time course of the synaptic conductances, the procedure is repeated for each ISI. By interpolation, a continuous estimation of
can be obtained, which is proved to be an accurate estimation of the prescribed conductances provided that they have slower variation (in time) than the membrane potential. Although the results obtained in [
26] show that using the
to estimate synaptic conductances in spiking regimes can be a suitable strategy, there were only applied to the McKean model, a very specific and non-realistic model, which is nothing but a caricature of the Fitzhugh–Nagumo system.
An improvement of this estimation procedure is to consider a different underlying mathematical model that could better fit experimental data. On the one hand, desirable candidates would be
conductance-based models as per the Hodgkin–Huxley formalism but, even though some of them (pyramidal cells [
28], stellate cells [
29], etc.) have been investigated in the context of estimation of conductances in subthreshold regimes, it is extremely difficult to devise a procedure for a generic conductance-based model. On the other hand,
integrate-and-fire (IF) models have also been considered as good approximations to the neuronal activity of different type of neurons (see [
30,
31,
32,
33,
34]) and are simple enough. We found that the
exponential integrate-and-fire (EIF) model represents an optimal balance between adaptability to experimental data and computational
curves, and model simplicity.
In this paper, following the ideas introduced in [
26], we propose a procedure to estimate conductances under a regular spiking regime. This procedure is based on a modification of the standard frequency–input curve, which consists of replacing the input constant current by a constant synaptic conductance (
curve). We aim to estimate the global synaptic conductance time course from a membrane potential time series of a spiking neuron, for which we can have an accurate
curve. For simplicity, here we assume that all synaptic input is excitatory.
The
curve can be provided in several ways: (I) from dynamic clamp experiments, using in vitro recording; (II) approximating the real data by a base model (IIa) for which we can obtain an analytic expression for the
curve, such as the leaky or the quadratic integrate-and-fire models (see [
35,
36]), or (IIb) allowing an approximate expression for the
curve, such as the McKean model (see [
26]) or the exponential integrate-and-fire (see
Section 2.1); finally, (III) using a more detailed model that can be numerically integrated in order to construct a
curve table, that is, providing a set of period-conductance pairs extracted from numerical experiments (see
Section 2.2). Once the
relationship is obtained, we have an expression of the period
depending on the conductances (or vice versa). In cases (I) and (III), where we have a discretization of this relationship, in order to obtain a continuous function
, one can interpolate the data using, for instance, the piecewise cubic Hermite interpolating polynomial (PCHIP). Therefore, given an interspike interval
, the corresponding conductance within this interval can be obtained by solving analytically or numerically the equation
.
Notice that for case (I), which involves dynamic clamp, the method can be considered to extract the
curve in in vitro experiments but not in in vivo as several injected currents would be required in order to have a precise
curve, which could add both variability across trials and noise measurement. Alternatively, we provide different computational strategies (II–III) to obtain the
curve. Even though strategies (IIa) and (IIb) provide good approximations to the
curve, they should not be applied in general to approximate the
curve for a wider range of neurons. For this reason, the computational approach presented in
Section 2.2 focuses on the idea of case (III).
The paper is organized as follows. In
Section 2.1, we present the neuronal model we use to generate data by means of prescribed conductances and its approximation to the EIF model. In
Section 2.2, we present the estimation procedure giving a pseudocode of its implementation sketched in
Figure 1. Next, in
Section 3, we show the results of the estimation procedures. Finally, in
Section 4 and
Section 5 we discuss the results obtained and present our conclusions.
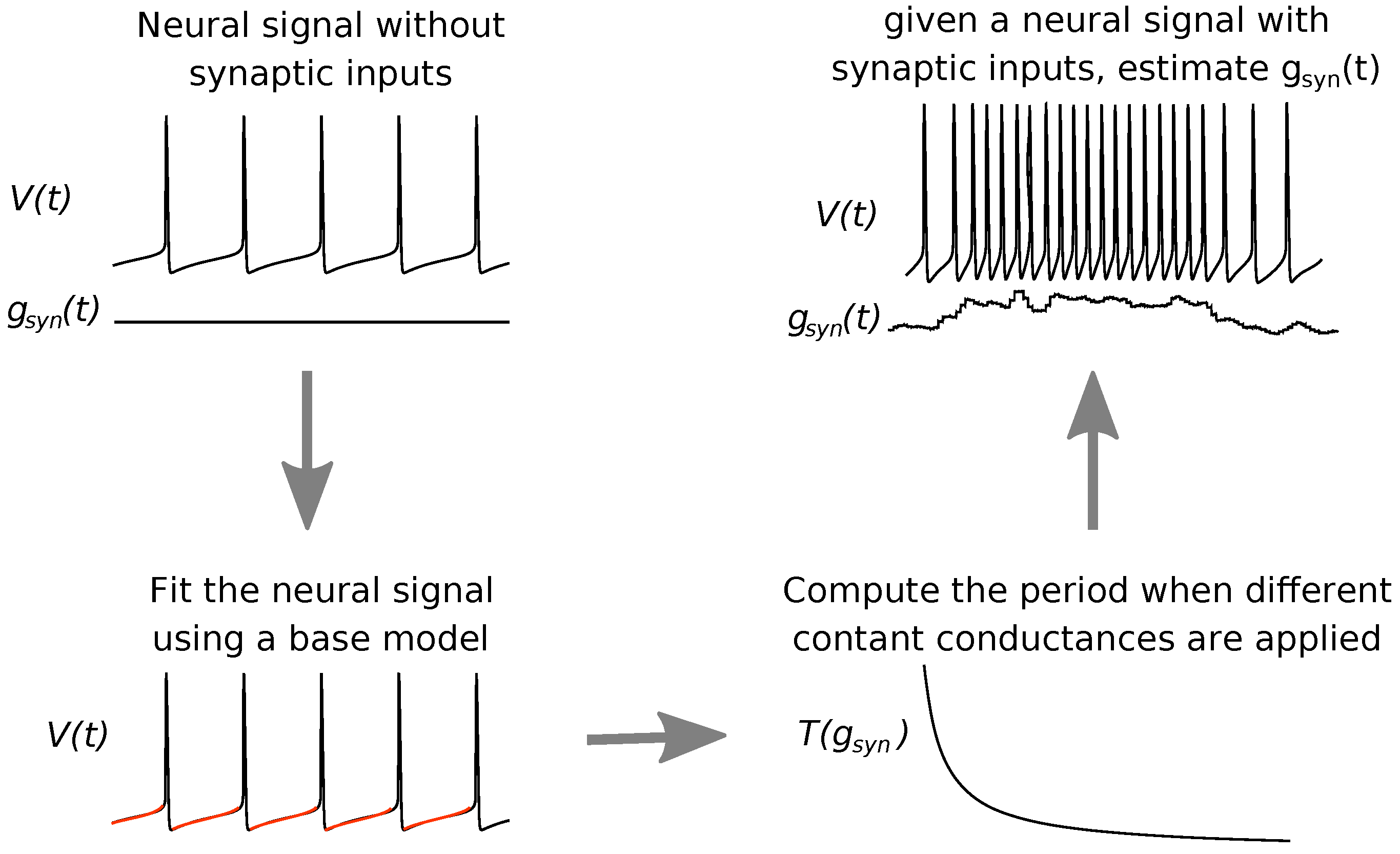
Figure 1.
Scheme of the estimation procedure performed by Algorithm 1. Given a neural signal from a neuron which is not receiving synaptic inputs (; upper left) but exhibiting regular spiking, we fit this signal using a computational model where the synaptic term is added afterwards (bottom left, where the red color stands for the fitting). Next, we use this model to compute the period (or frequency) for different constant synaptic conductances (, bottom right). We finally estimate the synaptic conductance time course from a neural signal subjected to synaptic inputs (upper right).
Figure 1.
Scheme of the estimation procedure performed by Algorithm 1. Given a neural signal from a neuron which is not receiving synaptic inputs (; upper left) but exhibiting regular spiking, we fit this signal using a computational model where the synaptic term is added afterwards (bottom left, where the red color stands for the fitting). Next, we use this model to compute the period (or frequency) for different constant synaptic conductances (, bottom right). We finally estimate the synaptic conductance time course from a neural signal subjected to synaptic inputs (upper right).
| Algorithm 1 Estimation procedure |
Data collection. Let be the membrane potential of a neuron on some time interval J in absence of synaptic inputs. Fitting data to a base model. Fit to a base model (for instance, an integrate-and-fire model). Call the set of parameters of the base model that gives the best fitting. The curve for the base model. Build up the function for the base model with the set . Depending on the base model, one possibility is to find either analytically, or its inverse and then invert it. Otherwise, we will proceed computationally:
- (a)
Consider a discretization of N equally-spaced synaptic conductances, that is, where , for all , where is a constant, assuming that the base model is regularly spiking for . - (b)
By numerical integration, for each with , obtain the period of the corresponding membrane potential. Note that this correspondence could also be obtained using one of the three other options presented in the Introduction (page 3). In all cases, a table containing the relation between and is obtained. Then, interpolate it to obtain a continuous function using, for instance, a piecewise cubic Hermite interpolating polynomial (PCHIP).
Estimation: computing ISIs. Let be the membrane potential trace, for some time interval , of neuron subject to synaptic input bombardments. Assume that emits spikes along the time span and let , for , be the j- interspike interval of . Estimation: inference. For each , , compute to obtain the corresponding estimated conductance . Estimation: interpolation. Interpolate the set , where is a point on the ISI. Here, we take the endpoint of the ISI and use a piecewise cubic Hermite interpolating polynomial (PCHIP).
- *
Note that for those models such that a qualitative expression of the curve can be provided, the interpolation step does not proceed.
|
3. Results
In order to test the estimation procedure, we apply Algorithm 1 (see
Section 2.2) to membrane potential traces, often referred to as
actual membrane potential traces, obtained computationally from the pyramidal cell model (see
Section 2.1.1). This model will be considered either with additive noise (
3), or without it (
1). Moreover, in our simulations we consider the model with different synaptic drives to simulate spiking regimes without (
Section 3.1) and with (
Section 3.2) short subthreshold regimes interspersed (see
Section 2.1.2 for more details in the synaptic drive).
For the different synaptic drives, we perform the estimation of conductances using different base models in Algorithm 1, step 3b. First, as we know that, by construction, the actual membrane potential follows the pyramidal neuron model, we consider this model as the base model, so we can test how powerful the estimation procedure is if we know the exact neural model. In this case, we deepen our results by exploring the cases where the data have been obtained with different amounts of noise and where the curve has been constituted using different discretizations.
However, in most of the cases, the specific model that describes the neural data we have in consideration is not always known; therefore, an approximation of it is required. For this reason, second, we use the exponential integrate-and-fire (EIF) model with constant adaptation as a base model and try to approximate the activity of the pyramidal cells to it (as in [
30]; see also
Section 2.1).
Finally, we explore the situation of having a small sample of
pairs which are obtained with some additive noise rather than having a reference model to obtain a more precise
curve. This specific case could be given when trying to obtain the
curve experimentally, where data are obtained with measurement noise and variability across trials. In this case, we first generate a small sample of
pairs where we add a small bias in order to simulate possible errors in the recordings. Then, we use a proper fitting to obtain the
curve to be able to proceed with the estimation procedure (see
Section 3.1.4 for more details on how). We will refer to this curve as the
experimental curve.
In
Table 1, we provide a summary of the results found in the different subsections.
3.1. Estimation in the Spiking Regime
In this section, we present the results obtained from simulations of the noisy pyramidal cell model using the prescribed three-frequency oscillatory drive defined in (
8). This configuration allows us to obtain a membrane potential trace in the spiking regime without interspersed silent periods. The estimation has been performed using, as base model, the three mentioned cases: the pyramidal cell model (
Section 3.1.1), the EIF model (
Section 3.1.3), and the
experimental curve (
Section 3.1.4).
3.1.1. Estimation When the Neural Model Is Known
In this case, we apply the estimation procedure given in Algorithm 1 considering the pyramidal neural model as a base model in step 3b of the algorithm.
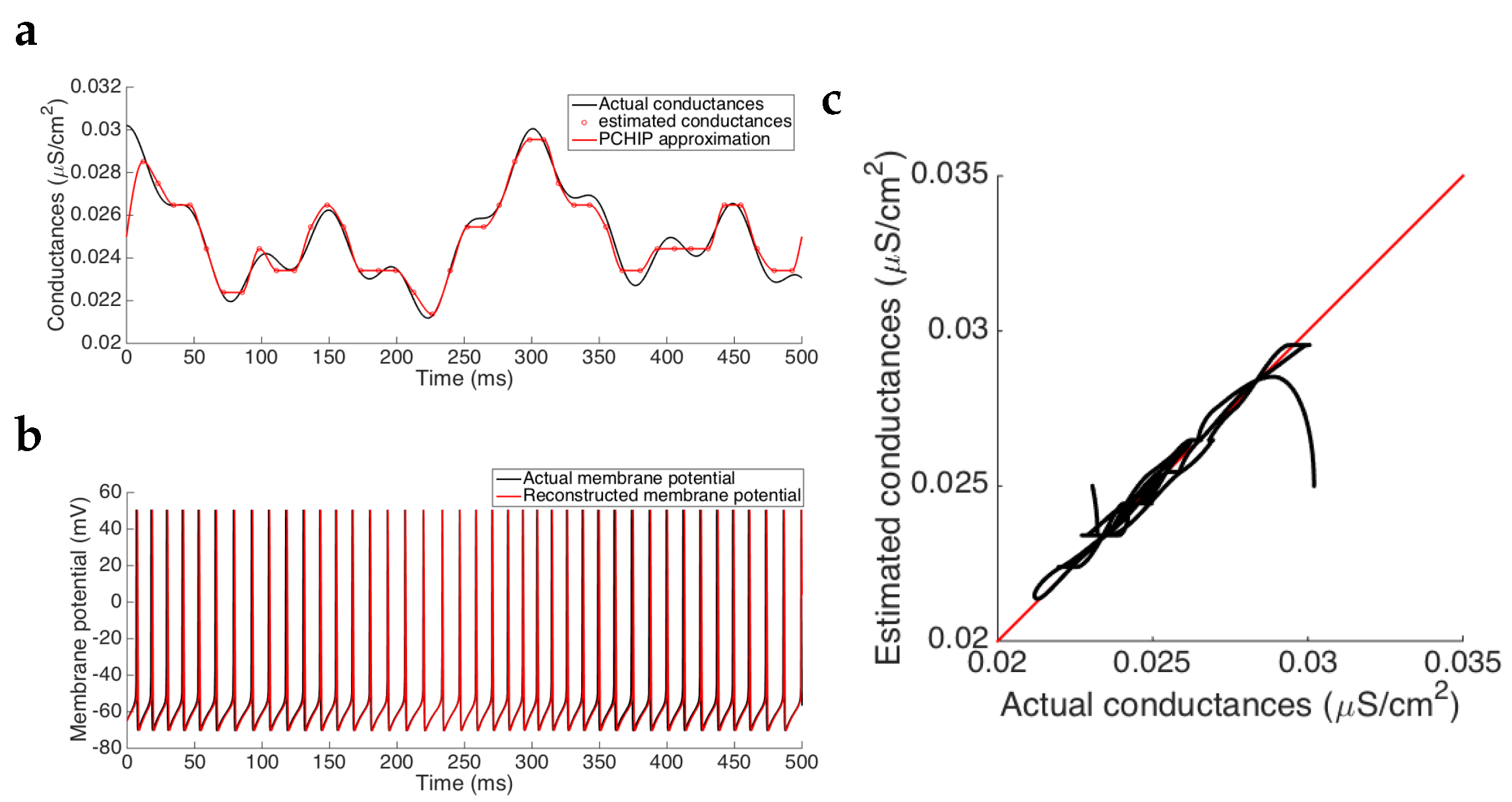
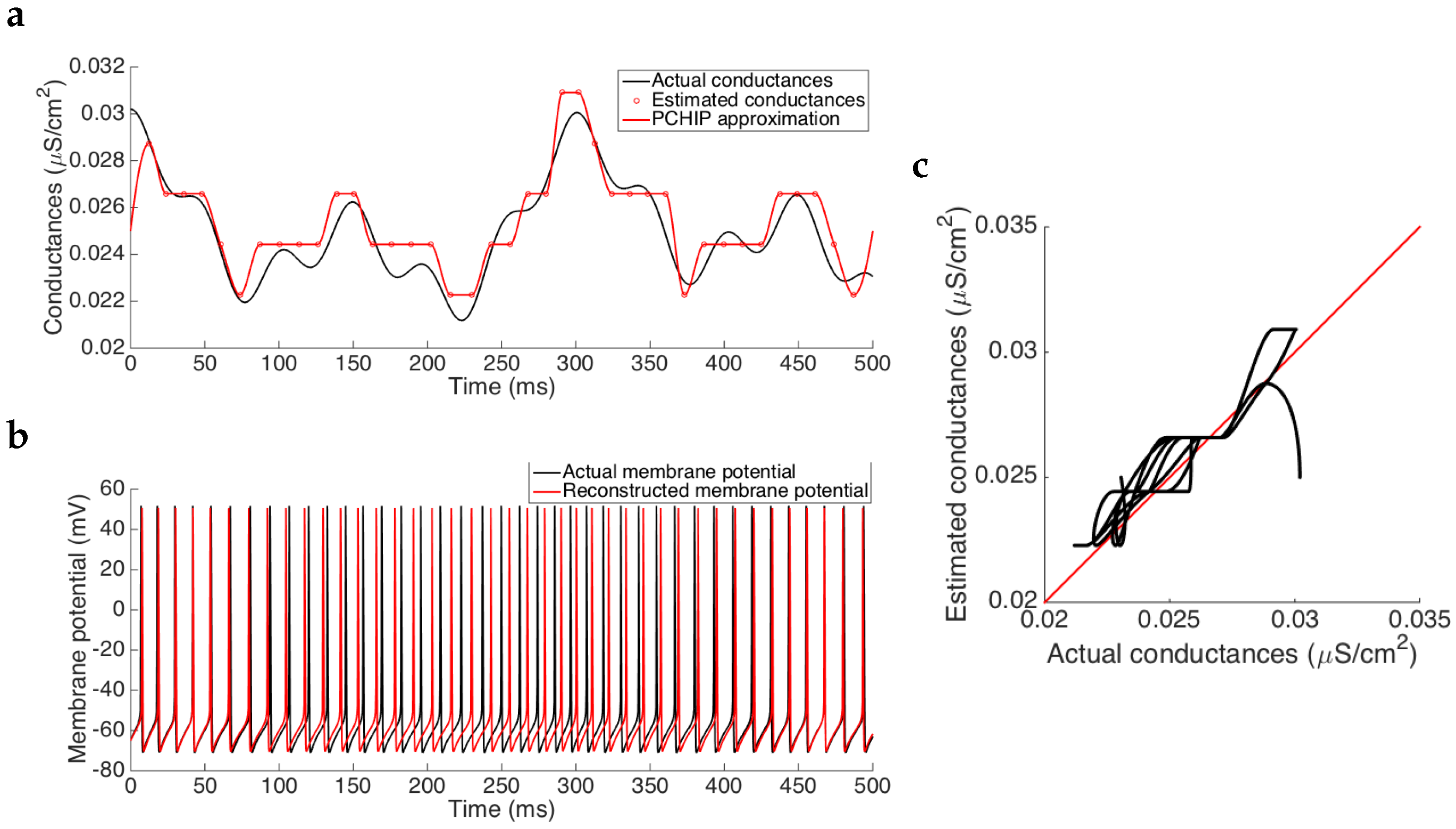
Results show that the estimated synaptic conductance trace follows the actual one before and after interpolating the estimated values (see
Figure 3a). After a short transient discrepancy observed in the first 20 ms, due to the the fact that we cannot have an a priori guess of the actual initial conditions, we observe a strong agreement between the estimation and the actual values, as we can see in
Figure 3c where the actual conductance trace versus the estimated one (after being interpolated) lies in the vicinity of the identity line, meaning a good approximation. In fact, computing the errors between the two traces, we have that the mean squared error is of order
and the relative error is, in mean, of order
, indicating that the Algorithm 1 accurately estimates the synaptic conductance time course (see
Table 2). Note that, in this case, as we are using the same model to generate the data and to estimate the conductances, the only source of errors could arise from the process of inversion, which is also related to the discretization provided by the spiking frequency of the neuron. Nevertheless, the magnitude of the errors obtained corroborates that they do not penalize the estimation (see
Figure 3b and the comment about the effects caused by the discretization in the next paragraphs). Moreover, the mean of the conductances after interpolating the data is also well estimated (relative error of
). Therefore, when we reconstruct the membrane potential by using the pyramidal cell model with the estimated synaptic conductances, neither the spike frequency, the spike amplitude nor the spike onset time are modified (see
Figure 3b), thus reinforcing the fact that the small errors caused in the estimation are not substantially important.
Two different sources of errors can arise during the estimation procedure. First, errors coming from the discretization of the conductances used to build up the table that relates spiking periods with constant synaptic conductance values and, second, the measurement noise, which is embedded in the membrane potential obtained from the recordings. Next, we discuss how these errors are reflected in the estimation.
Effects caused by the discretization
We want to analyze the influence on the estimations of the discretization of synaptic conductances in step 3b of Algorithm 1, that is, the effect of parameter
used to build up the
table. For this reason, and to avoid errors coming from additive noise, the actual membrane potential is also generated without considering noise. Then, we explore the errors (mean square error (MSE)) for different discretization steps (see
Table 2). As the range of the actual conductances is approximately
S/cm
(see
Figure 3a), we start using a step size of
. Results show that the discretization has no influence on the goodness of the procedure as increasing the discretization step does not yield a substantial increase of the errors. These results show that, when the base model is exactly the one that the neuron follows, the error of the estimation is given basically by the discretization of the conductances. Therefore, as the error coming from the discretization step size does not improve for values smaller than
and, for this value the precision of the estimation is already acceptable, from now on, we will consider this discretization step size.
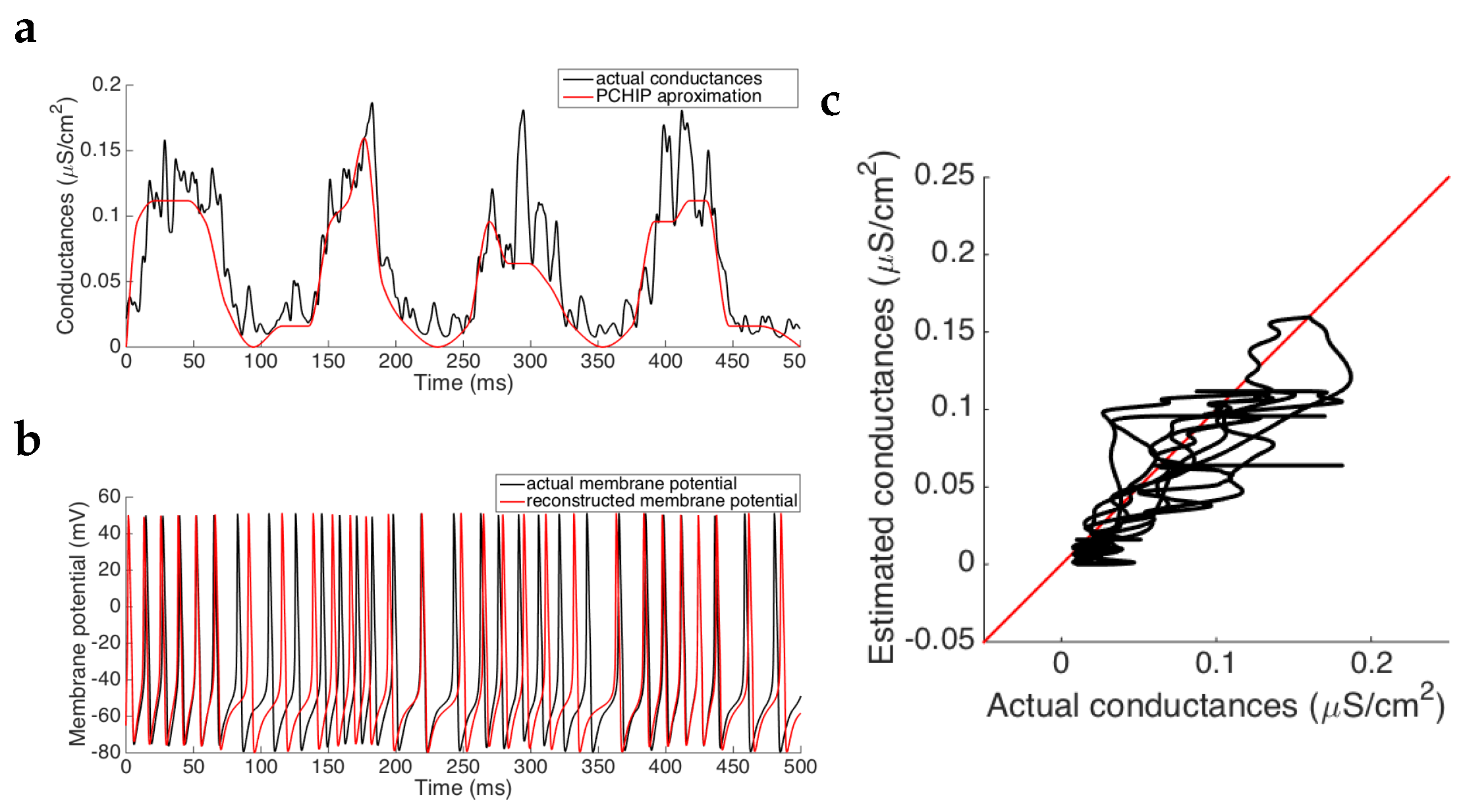
3.1.2. Estimation of Conductances for the Stellate Cell Model
In order to test the robustness of our method, the algorithm has been also applied to a membrane potential trace coming from the stellate cell model (see [
29]), with the synaptic input described in
Section 2.1.2. The dynamics of the system presents oscillations both in the spiking and in the subthreshold regime. The base model considered in this case is the same model used to generate the membrane potential, that is, the stellate cell model. We also obtain good approximations (MSE
), thus reinforcing the goodness of the estimation method.
Figure 4 portrays the results of the estimation procedure.
As we can see in
Figure 4a, the shape of the prescribed synaptic conductances is well approximated by that of the stellate cell model, even though some small fluctuations are missing and the estimation is not as good as the case of the pyramidal neuron. We can also see the goodness by plotting the actual conductances versus the estimated conductances, showing values in the vicinity of the identity line. Moreover, we obtain that the mean squared error is of approximately
, while the mean conductance has been estimated with a relative error of
.
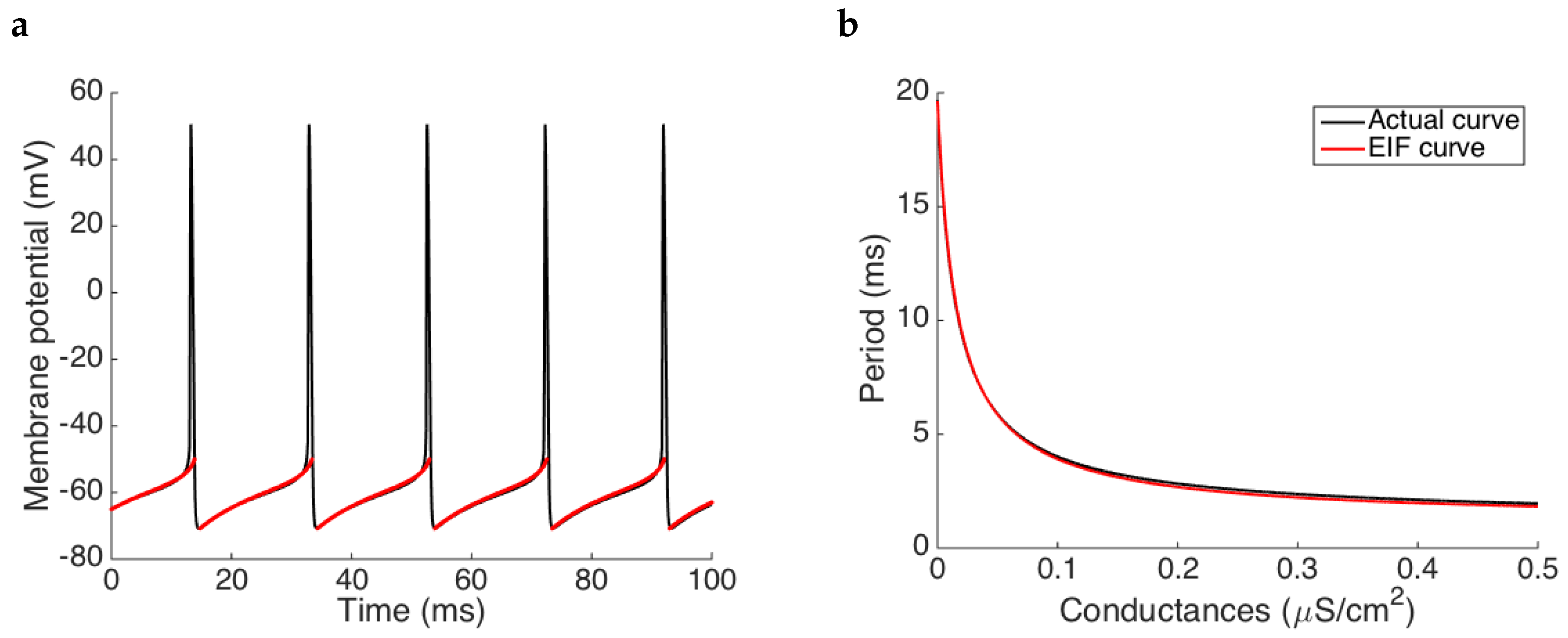
3.1.3. Estimation When the Neural Model Is Approximated by a Base Model
In many experimental cases, the exact model describing the target neuron is not known. One plausible way to apply our methodology is by fitting the measured data to a reference model. In this section, we explore this paradigm and study the errors of the estimation procedure. For this purpose, we generate membrane potential traces from the noisy pyramidal cell model (
3) (see also
Figure 2 in
Section 2) with the synaptic drive shown in
Figure 5a, and use them to fit an exponential integrate-and-fire model (
5). The underlying assumption is that the fitted EIF model provides a good approximation of the original model (see
Figure 2).
In order to challenge our methodology, we consider the worst scenario regarding data discretization and noise levels according to our previous results (see
Table 2 and
Table 3): we assume a discretization step size for the conductances (
) and a high standard deviation for the white noise in the membrane potential (
mV). Even so, results still show a good performance of the estimation procedure, see
Figure 5, unexpectedly similar to the results obtained in
Section 3.1.1.
As we can see in
Figure 5a, some of the small oscillations are tracked properly, yielding to some temporal shifts on the onset of spikes when reconstructing the membrane potential (see
Figure 5b). However, despite of this loss, the estimation keeps capturing the spiking frequency, and the conductances are estimated with a relative error of
(mean over time) whereas the MSE is
. After interpolating the data, the relative error drops to
and the MSE is equal to
. Note that the difference between these errors and those provided by the same case when we use the exact neural model in the estimation procedure does not differ qualitatively (see the first two rows of
Table 4). Therefore, even though we assume that the exact model is not known, we still obtain good estimates of the time course of the synaptic conductance extracting the
relationship from an approximated base model (EIF in this case).
In the estimation procedure we have built a conductances’ table to be able to make the estimation. However, note that for this specific case, and when the conductances are supposed to be constant, there exists a closed expression for the EIF period,
, see Equation (
6). This fact allows us to replace the table of the estimation procedure for the solution
, where
is the specific interspike considered in each iteration of the procedure. Knowledge of this expression allows us to estimate the conductances more efficiently and with less numerical errors (see third row in
Table 4).
3.1.4. Estimation without a Reference Base Model
To fully explore all possibilities of how to extract a relation between the oscillatory period and a constant value of the conductance, apart from considering an approximate model from which we can build up a table or find an explicit expression, we also consider the possibility of having the relationship experimentally. This relationship could be directly obtained by using the dynamic clamp technique or indirectly through the curve by using other techniques (such as current clamp). Indeed, if the current I only contains synaptic currents, then the curve can be obtained by computing and extracting the period T from the membrane potential trance.
Note that, if the
curve can be obtained with high accuracy (with a sufficiently fine discretization of points), then the estimation will present similar errors than those presented in
Section 3.1 (see
Table 2). However, if the discretization is poor, errors could be magnified.
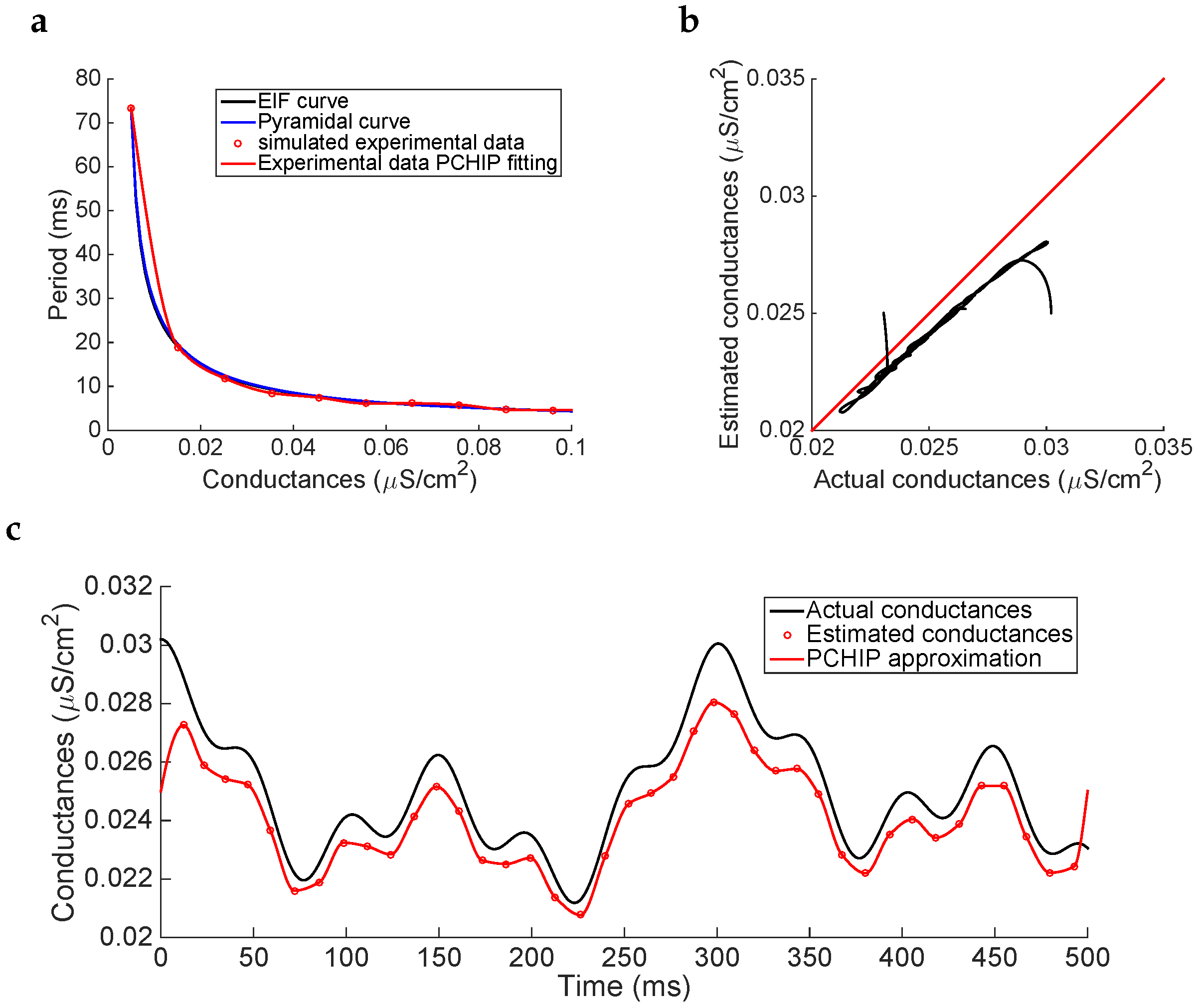
In this section, we investigate the case where only a small sample of
pairs are obtained. To computationally simulate this specific situation, we assume that the data set is generated from the noisy pyramidal cell model. By using this same model, we generate a small sample of ten
pairs to construct the
curve. Moreover, to consider possible errors in the recordings (measurement noise and variability across trials), we have added noise to the resulting set of points. This bias has been included by adding a random normal distributed value with 0 mean and 0.5 standard deviation to the period component. Finally, to estimate the conductance for any given period, and as we are supposing that no extra information is known about the actual data, we have interpolated these data using the PCHIP method (see
Figure 6a).
Applying now the estimation procedure to the membrane potential shown in
Figure 3b (black trace), using the simulated experimental curve instead of an approximate base model to generate the
curve, the results show how the estimated conductances are clearly underestimated (see
Figure 6b,c) and, even though the frequency of oscillation of conductances is maintained, the amplitude of the oscillation is reduced. This fact is mainly due to misestimations that arise in the
experimentally fitted curve (with respect to that obtained with the actual model), specially the underestimation obtained for those values of conductances between
and
, which is in fact the interval where the actual conductances lie.
However, in view of the magnitude of the conductances, these underestimations are not substantially significant, providing similar errors to the cases where the
curve is numerically computed from a base model (see fourth row in
Table 4).
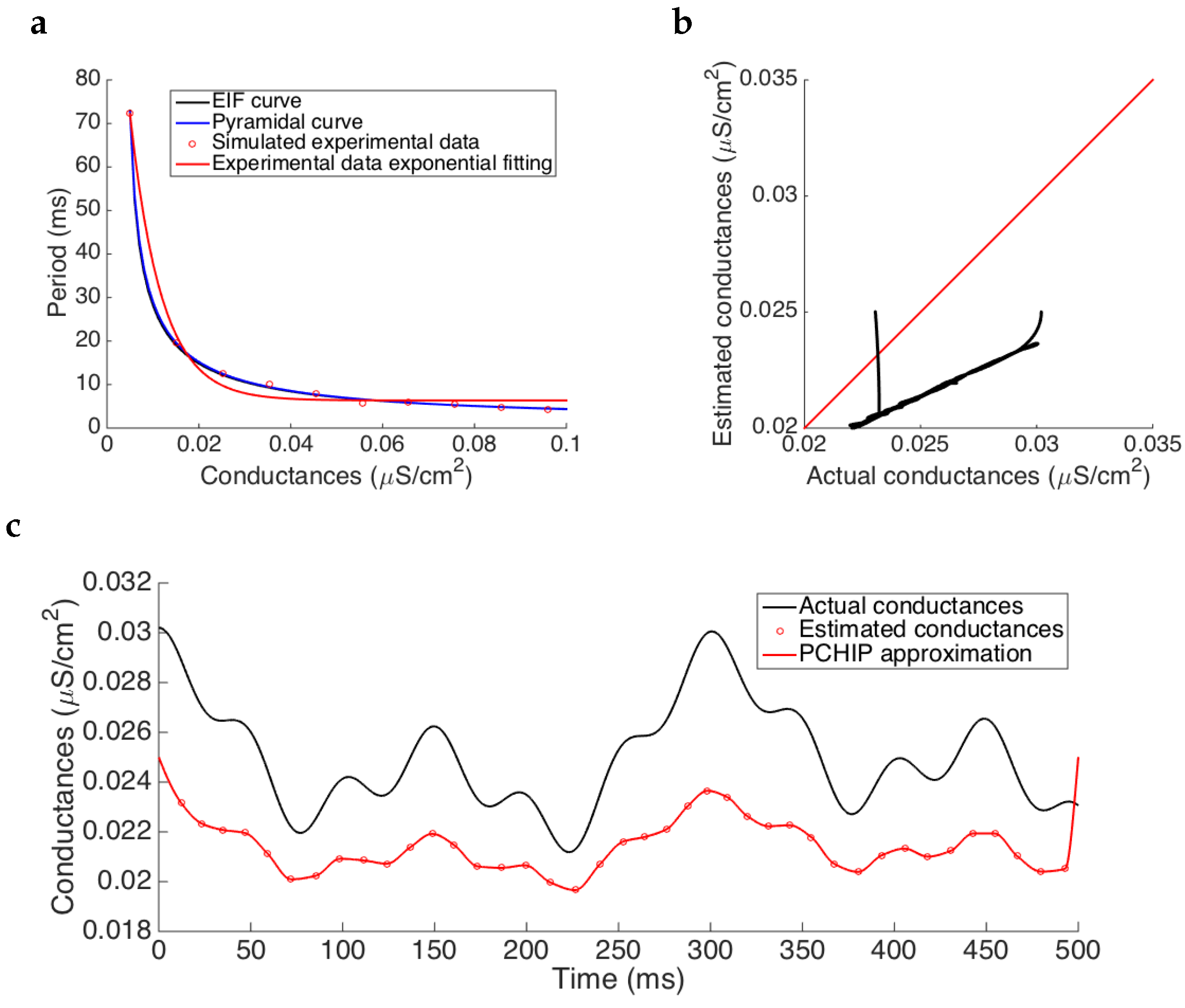
Note that the errors in the
curve can alter consistently the overall estimation causing an important increase of the errors in the estimation. One source of these misestimations is the interpolation method. We also tested an exponential fitting, instead of the PCHIP interpolation, which gave us higher misestimations of the
curve in the working interval, see
Figure 7. Results show, as previously, that the frequency of oscillation is maintained but the amplitude is worst underestimated providing an average relative error of approximately
. Therefore, the errors are substantially magnified showing the importance of having a good approximation of the
curve to be able to properly estimate the conductances.
3.2. Estimation in the Spiking Regime with Interspersed Short Subthreshold Periods
In this section, we aim to explore whether the estimation procedure can be also used when short periods of subthreshold activity interrupt the spiking regime. Note that, if those silent periods are sufficiently large (for instance, greater than 100 ms), one could use a combination of the spiking estimation procedure developed in
Section 2.2 and a subthreshold estimation procedure (for instance, the so-called
QIF method presented in [
23], which takes into account the spike initialization and possible subthreshold activity). However, for shorter subthreshold regimes, this combination could be not possible (as some methods require a certain amount of data to be used) or could cause a lot of misestimations induced by recent spike activity. For these reasons, we consider a synaptic conductance trace (see
Section 2.1.2), depicted in
Figure 8a (black trace), which causes spiking regimes interleaved with subthreshold regimes (of 50 ms of duration, approximately) when this is used into the pyramidal cell model (see
Figure 8b (black trace)).
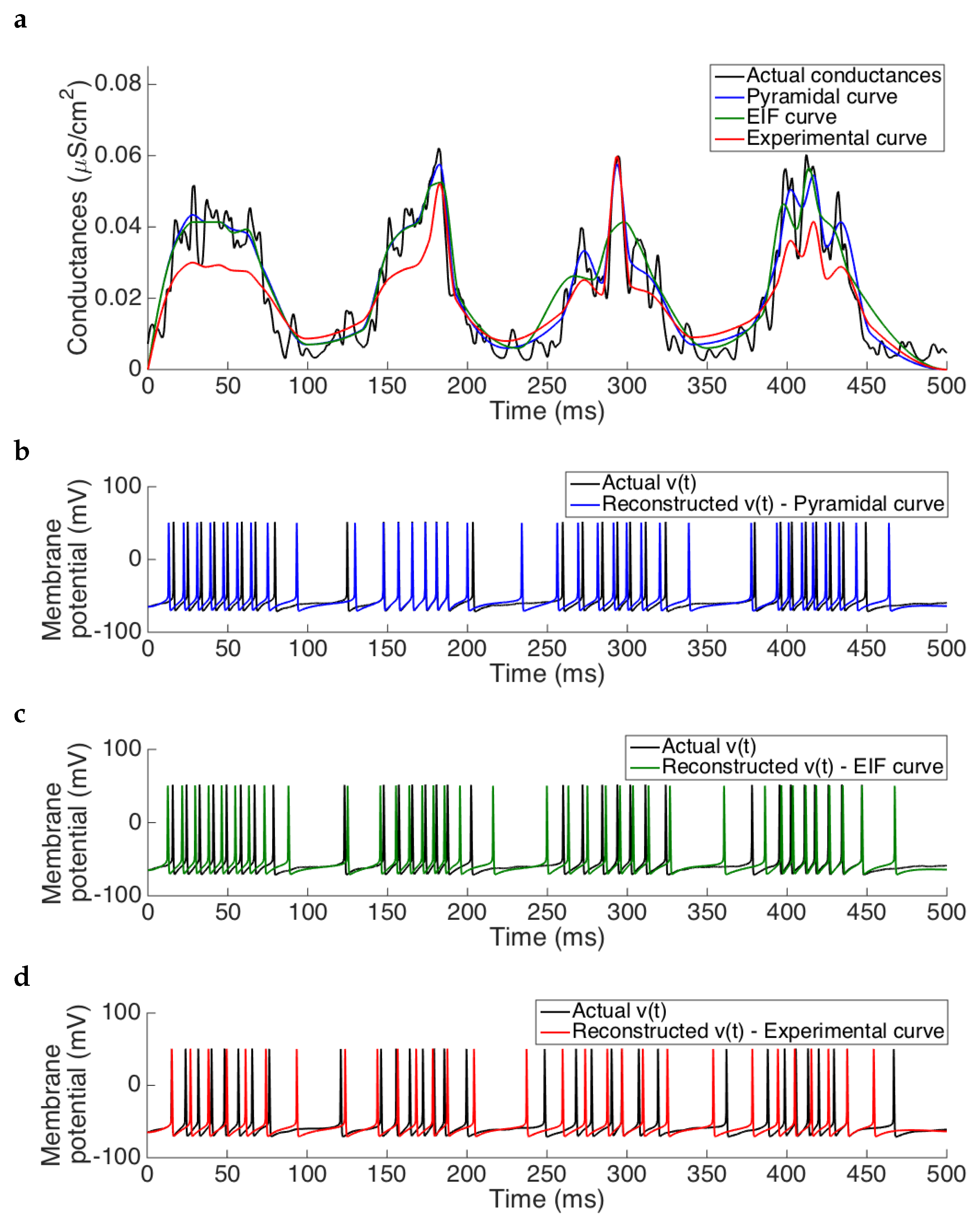
Results presented in
Figure 8a show that the estimation procedure provided in Algorithm 1 is able to reproduce the synaptic drive, either if we use as a base model to generate the
table the pyramidal cell model (blue trace) or the EIF model (green curve). In both cases, the reconstruction of the membrane potential using the estimated conductances maintains the spike frequency, thus keeping unchanged the firing rate (
Figure 8b,c, respectively). On the other hand, when the simulated experimental curve is used, conductances around
are not well estimated, as it happened when considering just the spiking regime, see
Section 3.1.4. These misestimates alter the spike frequency in the membrane potential reconstructed using the estimated conductances as synaptic inputs (see
Figure 8b), by adding extra spikes in the silent regime and displacing the spiking time-onset in the spiking regime.
Errors of the estimation in the different cases are presented in
Table 5, where we can see also that both the pyramidal cell model and its EIF approximation lead to similar errors, while the experimental curve leads to higher errors.
4. Discussion
Estimations of the time course of synaptic conductances arriving at a single cell (or a population) reveal relevant information to fully understand the functional connectivity of the brain. Several strategies have been developed for the estimation during subthreshold activity regimes [
3,
4,
5,
6,
7,
8,
9,
10,
12,
13,
14,
15,
16,
17,
18,
19,
21,
22,
23,
25]. However, the estimation during oscillatory (spiking) regimes has received less attention [
25,
26,
27,
40]. The main drawback is the predominance of ionic currents over synaptic ones in this regime. Nevertheless, as we already showed in a caricature model (see [
26]), if one gets the main information about the contribution of ionic currents and is able to track their activity during the spiking regime, then it is possible to infer the synaptic activity. In this paper, we extend this idea to more realistic spiking models and provide a systematic procedure that can be applied to a variety of neural data, coming both from computational simulations and from experiments. The idea is to exploit the input-response properties of the cell; more precisely, the relation between constant synaptic conductances and the spiking frequency (the
curve).
In the spiking regime, the most natural measurable quantity is the oscillation frequency, which constitutes a valuable piece of information for our purpose. The frequency–input relationship encompasses the average response of the neuron, which includes both the direct effect of the input stimulus and the resulting activation of ionic currents. The fact that
curves are usually monotone implies the one-to-one correspondence between frequency and input, and favors the resolution of the inverse problem. However, while in the case of a steady input (
I constant) that induces a constant-frequency repetitive firing, it is straightforward to infer the input current from the output frequency; the presence of the neuron’s voltage in the synaptic current (
in their minimal expression) makes the estimation of the total input current highly sensitive to the fluctuations in
. Here, we propose to consider an alternative frequency–input relationship, the
curve, which is more appropriate for the estimation problem since it implicitly incorporates the voltage dependency of the synaptic current and mitigates the effects of the fluctuations. In a mathematical model, we can obtain the
or
curve,
, numerically (in some cases, even analytically or quasi-analytically, see [
26] and
Section 2.1.1). For experimental studies, retrieving the
curve requires the use of dynamic clamp techniques on the isolated cell for fixed
values within a physiologically plausible interval. In Algorithm 1, we summarize the steps to perform the estimation of synaptic conductances from the interspike intervals of the voltage traces and the knowledge of the
curve.
The second ingredient of the method presented in this paper is the choice of a reference/base model, which must be both flexible to fit to a variety of neural data (choosing a suitable parameter set in each case) and simple enough to be able to compute the
curve in a precise way. We found the exponential integrate-and-fire model (EIF) to be a convenient base model: on the one hand, its oscillation period can be well approximated by means of the semi-analytical formula (
6); on the other, there are examples in the literature showing that its
curve can fit
curves of neural data.
Our procedure has been applied first to a representative computational model driven by a prescribed input conductance trace. We verify that the use of the
curve brings excellent predictions of the
fluctuations along the prescribed trace, see
Figure 3. We also observe better estimations when the synaptic conductances change on a time-scale longer than the mean interspike interval. We also analyze the effects of the discretization and the measurement noise, see
Table 2 and
Table 3. In particular, we observe that levels of noise below 1 mV do not compromise the estimations in a substantial way since the MSE of the estimations is very low (MSE
) in all cases. In
Section 3.1.3, we explore the goodness of the estimations under the assumption that the EIF (base) model provides a good fit to the neural data. By using a model for a noisy pyramidal cell to generate the data, we again observe excellent estimations of the synaptic conductance (MSE
), see
Figure 5. We conclude that we could reasonably estimate the synaptic conductances from other neural data to which the EIF could be fitted.
We next study potential difficulties that can be found in more realistic scenarios. First, in
Section 3.1.4, we cope with the problem of estimating synaptic conductances when there is no base model that decently fits the data. We propose a procedure to come up with an approximation of the
curve and apply it to data generated by a computational model. Again, we achieve good estimations (MSE
), see
Table 4. For real cells, the dynamic-clamp technique allows obtaining the period
T from a
value, and therefore it would be interesting to design a protocol so that it was feasible to obtain the
curve. As our method is mainly addressed to the spiking regime, the second potential difficulty is the presence of interspersed short subthreshold periods. We show (see
Figure 8) that these short subthreshold periods do not contaminate the estimation of synaptic conductances and provide results with small errors.
In order to check the robustness of the estimation procedure for other types of neurons, we have also considered the stellate cell model (see
Section 3.1.2), to which the quadratic integrate and fire model (QIF; see [
30]) has been fitted. Results (not shown) reveal that the estimation procedure can also be used in cases where the fitting is not as perfect as those obtained for the pyramidal cell model.
Our method is related in spirit to the firing-clamp method presented in [
25], where the authors also consider a point neuron and, by calibrating its activity, they experimentally build up an approximated function to describe the changes in both the membrane potential threshold and the amplitude of the spiking regime (instead of the spiking frequency). Their model is able to discern between excitatory and inhibitory conductances; however, as their calibration is purely experimental, errors could arise from the measurement noise because of repetitive trials.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}