
A Model for Brucellosis Disease Incorporating Age of Infection and Waning Immunity



Abstract
:1. Introduction
Mathematical Model
- with ([30]);
- The functions , belong to and for almost every and for some ;
- The function is positive, bounded, and uniformly continuous on .
- The initial conditions are such that , .
2. Well-Posedness Results
3. Basic Reproduction Number
4. Endemic Equilibria and Their Stability
5. Numerical Experiments
5.1. Model Parameters
5.2. Numerical Simulation
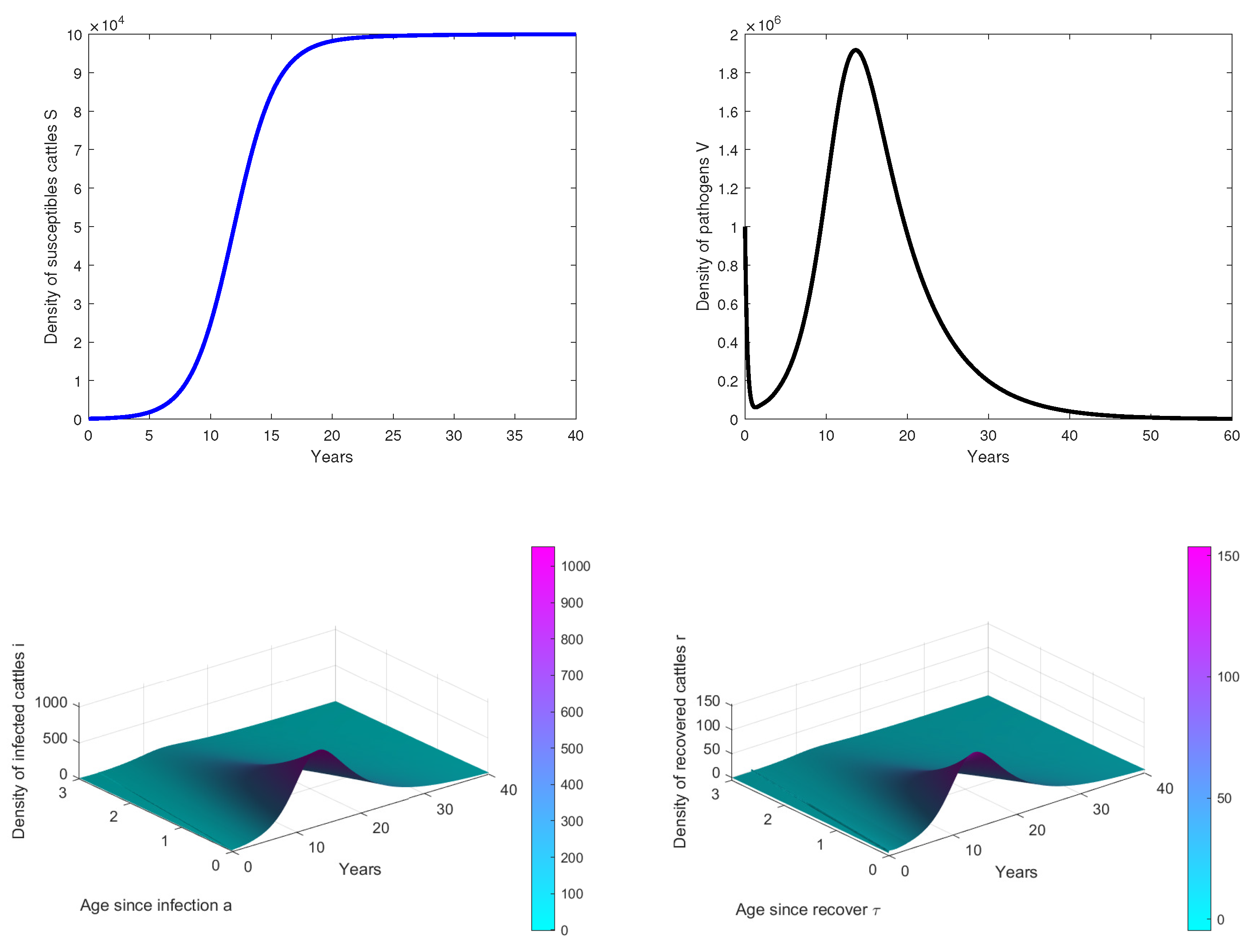
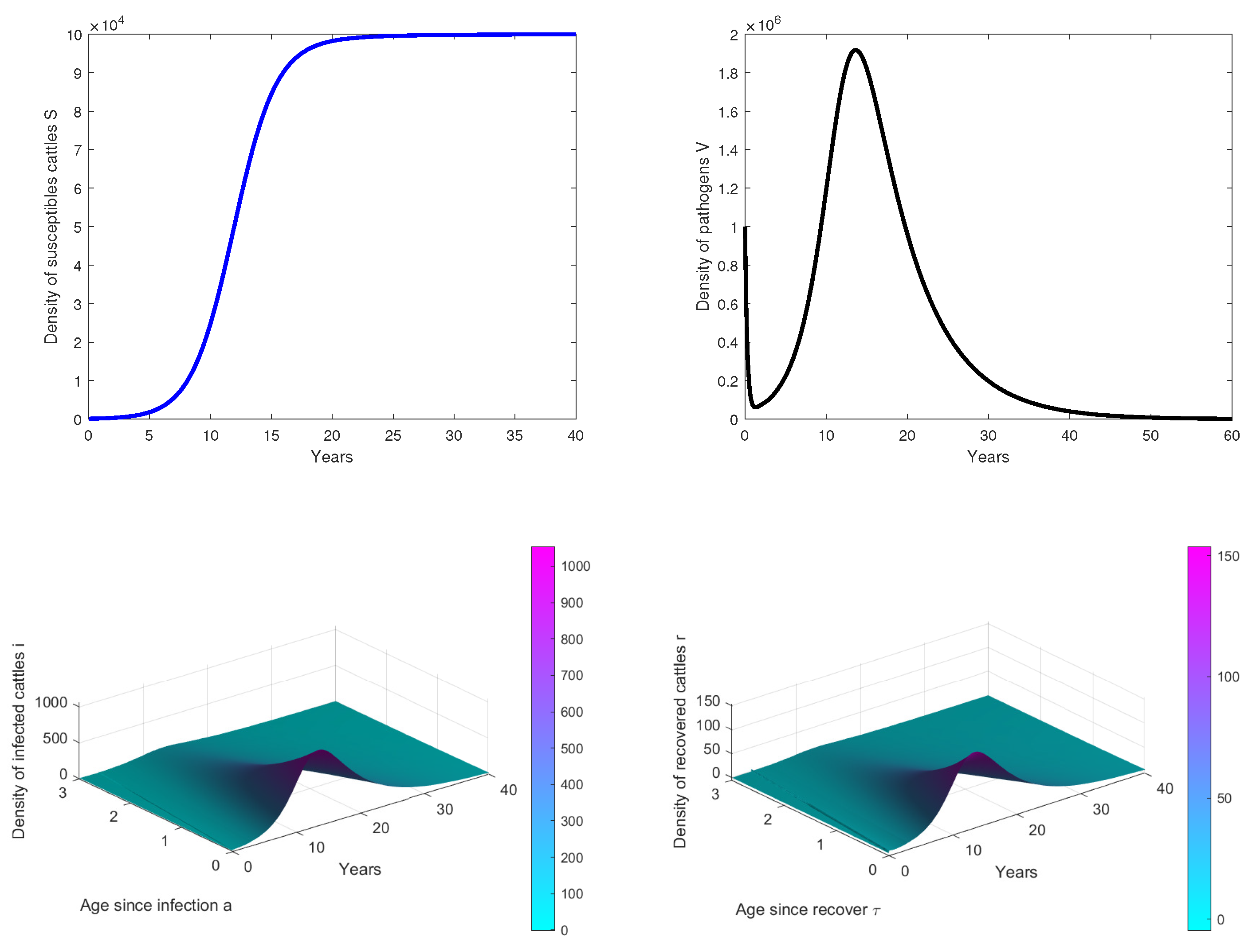
- In Figure 2, we observe that the disease dies out in the population when, . This confirms our theoretical results.
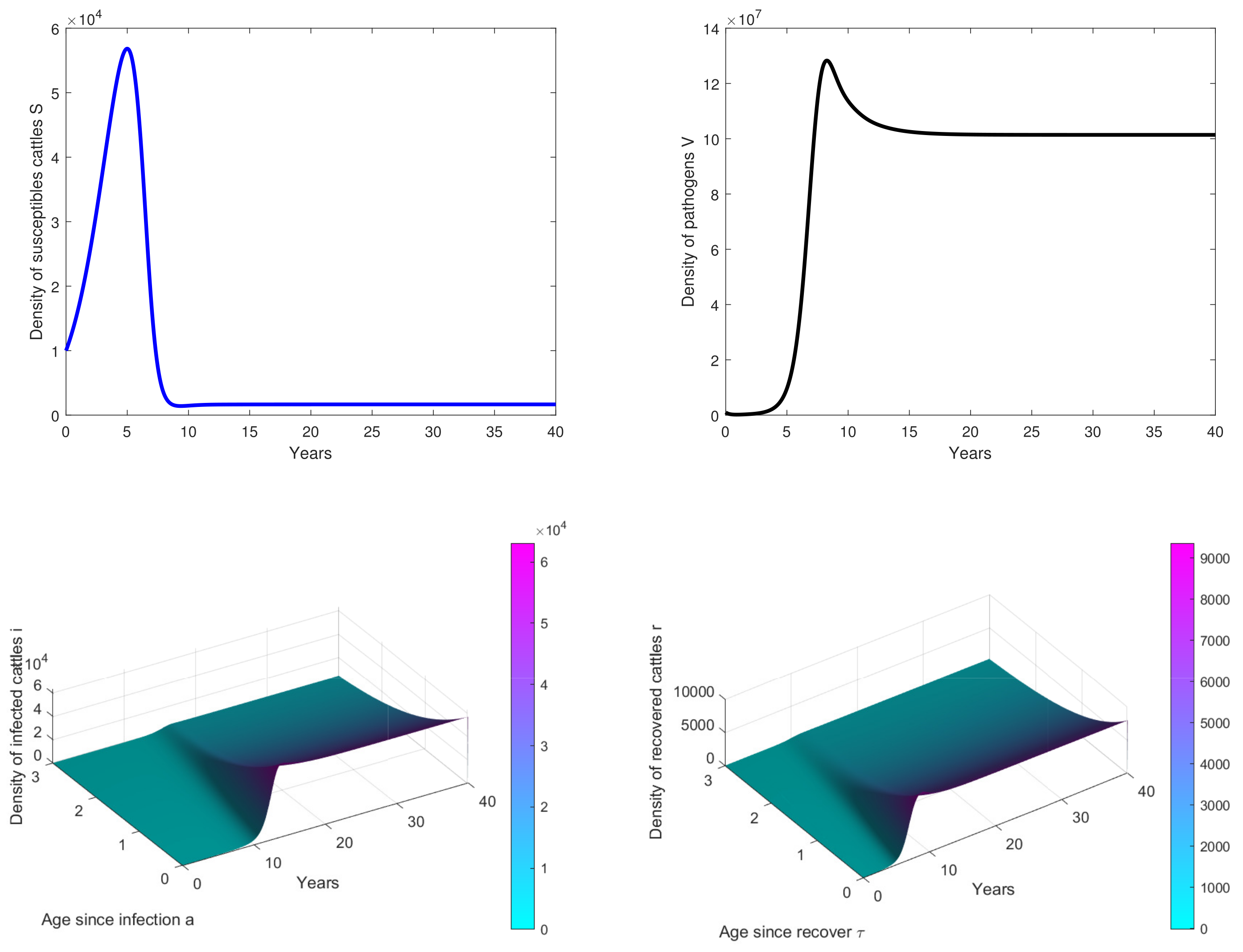
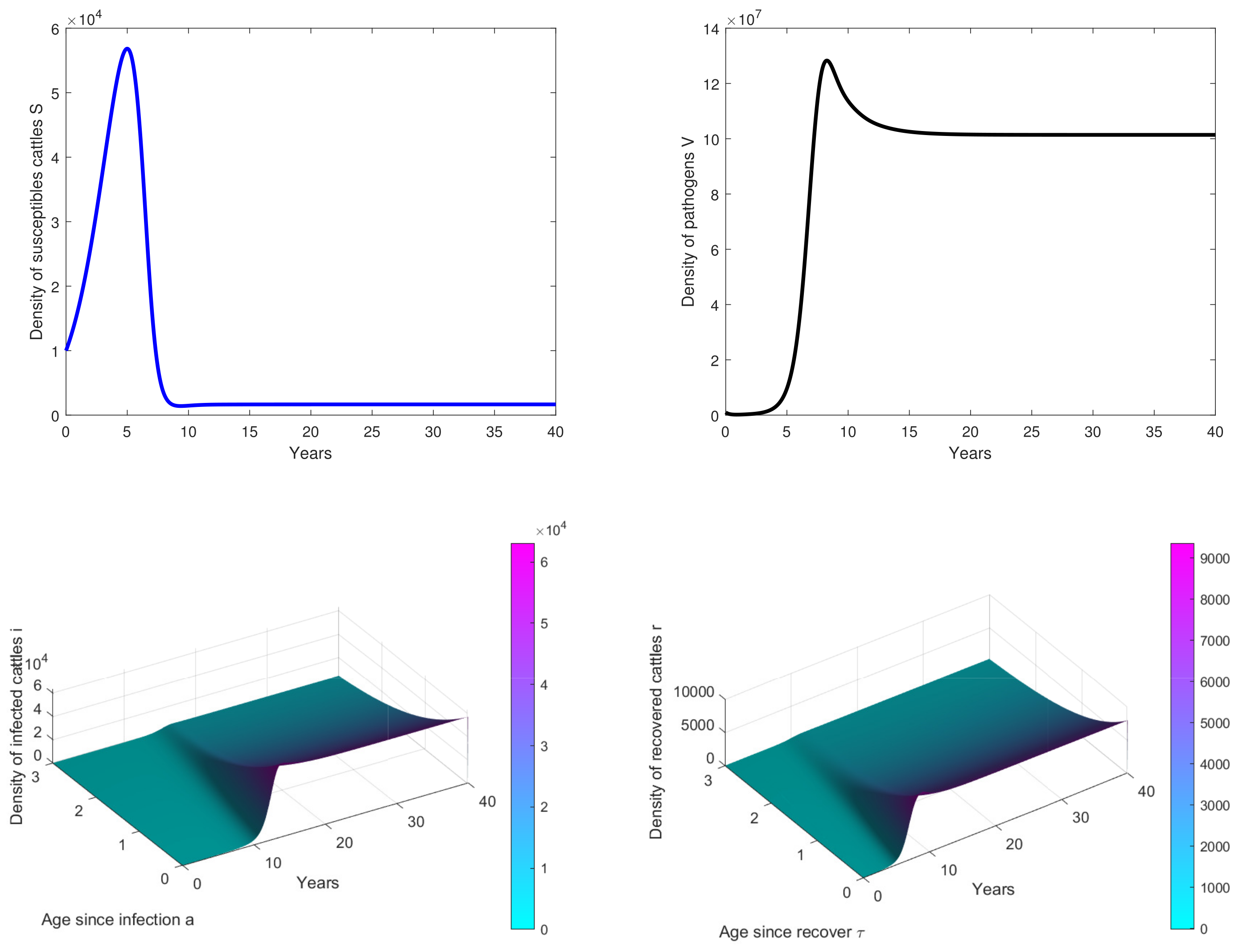
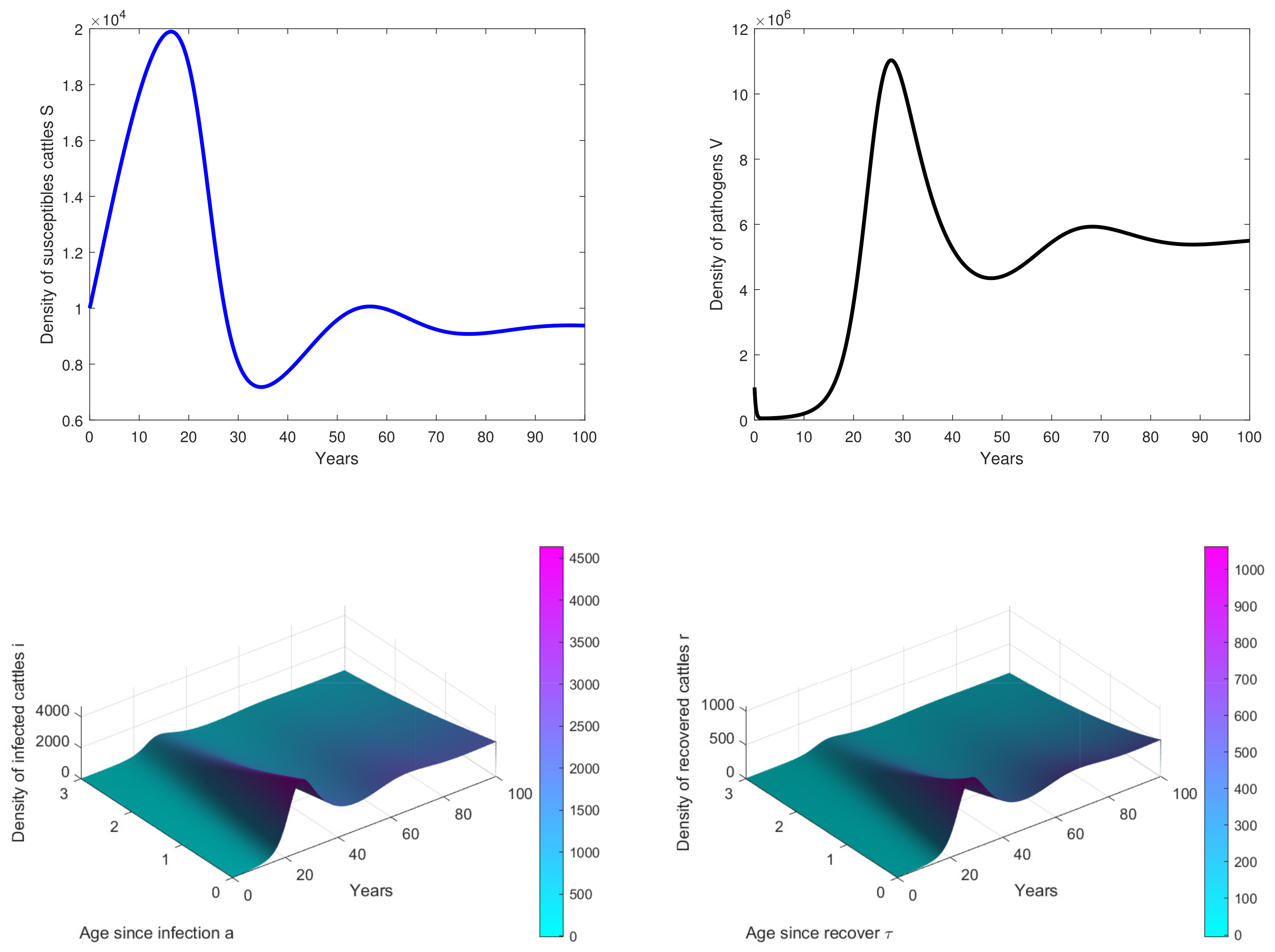
- In Figure 3, the initial dose is and we observe that the peak of infection is reached about 14 years post-infection. The figure shows that the disease could persist in the population when .
- In Figure 4, , we observe that the peak of infection is reached about 10 years post-infection and there are some oscillations on the curves. This means that the birth rate of cattle significantly influences the dynamics of the disease.
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godfroid, J. Brucellosis in livestock and wildlife: Zoonotic diseases without pandemic potential in need of innovative one health approaches. Arch. Public Health 2017, 75, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Omer, M.; Skjerve, E.; Holstad, G.; Woldehiwet, Z.; Macmillan, A. Prevalence of antibodies to Brucella spp. in cattle, sheep, goats, horses and camels in the State of Eritrea; influence of husbandry systems. Epidemiol. Infect. 2000, 125, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Megersa, B.; Biffa, D.; Abunna, F.; Regassa, A.; Godfroid, J.; Skjerve, E. Seroepidemiological study of livestock brucellosis in a pastoral region. Epidemiol. Infect. 2012, 140, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, G.Q.; Sun, X.D.; Hou, Q.; Li, M.; Huang, B.; Wang, H.; Jin, Z. Prediction and control of brucellosis transmission of dairy cattle in Zhejiang Province, China. PLoS ONE 2014, 9, e108592. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, C.A.; Arenas-Gamboa, A.M.; Maurizio, E. Caprine brucellosis: A historically neglected disease with significant impact on public health. PLoS Negl. Trop. Dis. 2017, 11, e0005692. [Google Scholar] [CrossRef] [Green Version]
- Diacovich, L.; Gorvel, J.P. Bacterial manipulation of innate immunity to promote infection. Nat. Rev. Microbiol. 2010, 8, 117–128. [Google Scholar] [CrossRef]
- Dorneles, E.M.; Teixeira-Carvalho, A.; Araújo, M.S.; Sriranganathan, N.; Lage, A.P. Immune response triggered by Brucella abortus following infection or vaccination. Vaccine 2015, 33, 3659–3666. [Google Scholar] [CrossRef]
- Bang, B. Infectious Abortion in Cattle. J. Comp. Pathol. Ther. 1906, 19, 191–202. [Google Scholar] [CrossRef]
- Abatih, E.; Ron, L.; Speybroeck, N.; Williams, B.; Berkvens, D. Mathematical analysis of the transmission dynamics of brucellosis among bison. Math. Methods Appl. Sci. 2015, 38, 3818–3832. [Google Scholar] [CrossRef]
- Hou, Q.; Sun, X.; Zhang, J.; Liu, Y.; Wang, Y.; Jin, Z. Modeling the transmission dynamics of sheep brucellosis in Inner Mongolia Autonomous Region, China. Math. Biosci. 2013, 242, 51–58. [Google Scholar] [CrossRef]
- Li, M.T.; Sun, G.Q.; Zhang, W.Y.; Jin, Z. Model-based evaluation of strategies to control brucellosis in China. Int. J. Environ. Res. Public Health 2017, 14, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lolika, P.O.; Modnak, C.; Mushayabasa, S. On the dynamics of brucellosis infection in bison population with vertical transmission and culling. Math. Biosci. 2018, 305, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Lolika, P.O.; Mushayabasa, S. On the role of short-term animal movements on the persistence of brucellosis. Mathematics 2018, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Nyerere, N.; Luboobi, L.S.; Mpeshe, S.C.; Shirima, G.M. Mathematical model for the infectiology of brucellosis with some control strategies. New Trends Math. Sci. 2019, 4, 387–405. [Google Scholar] [CrossRef]
- Paride, P.O.L. Modeling the Transmission Dynamics of Brucellosis. Ph.D. Thesis, University of Zimbabwe, Harare, Zimbabwe, 2019. [Google Scholar]
- Zhang, W.; Zhang, J.; Wu, Y.P.; Li, L. Dynamical analysis of the SEIB model for Brucellosis transmission to the dairy cows with immunological threshold. Complexity 2019, 2019, 6526589. [Google Scholar] [CrossRef] [Green Version]
- Hou, Q.; Sun, X.D. Modeling sheep brucellosis transmission with a multi-stage model in Changling County of Jilin Province, China. J. Appl. Math. Comput. 2016, 51, 227–244. [Google Scholar] [CrossRef]
- Lolika, P.O.; Mushayabasa, S. Dynamics and stability analysis of a brucellosis model with two discrete delays. Discret. Dyn. Nat. Soc. 2018, 2018, 6456107. [Google Scholar] [CrossRef]
- Sun, G.Q.; Li, M.T.; Zhang, J.; Zhang, W.; Pei, X.; Jin, Z. Transmission dynamics of brucellosis: Mathematical modelling and applications in China. Comput. Struct. Biotechnol. J. 2020, 18, 3843. [Google Scholar] [CrossRef]
- Tumwiine, J.; Robert, G. A mathematical model for treatment of bovine brucellosis in cattle population. J. Math. Model. 2017, 5, 137–152. [Google Scholar]
- Lolika, P.O.; Mushayabasa, S.; Bhunu, C.P.; Modnak, C.; Wang, J. Modeling and analyzing the effects of seasonality on brucellosis infection. Chaos Solitons Fractals 2017, 104, 338–349. [Google Scholar] [CrossRef]
- Yang, J.; Xu, R.; Li, J. Threshold dynamics of an age–space structured brucellosis disease model with Neumann boundary condition. Nonlinear Anal. Real World Appl. 2019, 50, 192–217. [Google Scholar] [CrossRef]
- Yang, J.; Xu, R.; Sun, H. Dynamics of a seasonal brucellosis disease model with nonlocal transmission and spatial diffusion. Commun. Nonlinear Sci. Numer. Simul. 2021, 94, 105551. [Google Scholar] [CrossRef]
- Ainseba, B.; Benosman, C.; Magal, P. A model for ovine brucellosis incorporating direct and indirect transmission. J. Biol. Dyn. 2010, 4, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Nannyonga, B.; Mwanga, G.; Luboobi, L. An optimal control problem for ovine brucellosis with culling. J. Biol. Dyn. 2015, 9, 198–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganiere, J.P. La brucellose animale. Ecoles Nationales Vétérinaires Françaises Merial, 2004; 1–47. [Google Scholar]
- Richard, Q.; Choisy, M.; Lefèvre, T.; Djidjou-Demasse, R. Human-vector malaria transmission model structured by age, time since infection and waning immunity. Nonlinear Anal. Real World Appl. 2022, 63, 103393. [Google Scholar] [CrossRef]
- Magal, P.; Seydi, O.; Wang, F.B. Monotone abstract non-densely defined Cauchy problems applied to age structured population dynamic models. J. Math. Anal. Appl. 2019, 479, 450–481. [Google Scholar] [CrossRef] [Green Version]
- Martcheva, M.; Thieme, H.R. Progression age enhanced backward bifurcation in an epidemic model with super-infection. J. Math. Biol. 2003, 46, 385–424. [Google Scholar] [CrossRef]
- Roth, F.; Zinsstag, J.; Orkhon, D.; Chimed-Ochir, G.; Hutton, G.; Cosivi, O.; Carrin, G.; Otte, J. Human health benefits from livestock vaccination for brucellosis: Case study. Bull. World Health Organ. 2003, 81, 867–876. [Google Scholar]
- Magal, P.; McCluskey, C.; Webb, G. Lyapunov functional and global asymptotic stability for an infection-age model. Appl. Anal. 2010, 89, 1109–1140. [Google Scholar] [CrossRef]
- Kenne, C.; Dorville, R.; Mophou, G.; Zongo, P. An Age-Structured Model for Tilapia Lake Virus Transmission in Freshwater with Vertical and Horizontal Transmission. Bull. Math. Biol. 2021, 83, 1–35. [Google Scholar] [CrossRef]
- Jiao, J.; Yang, J.; Yang, X. The dairy cattle brucellosis and prevention and control (Nainiu bulujunbing jiqi fangkong). Anim. Husb. Feed. Sci. 2009, 30, 170–171. [Google Scholar]
- Dobson, A.; Meagher, M. The population dynamics of brucellosis in the Yellowstone National Park. Ecology 1996, 77, 1026–1036. [Google Scholar] [CrossRef]
- Sun, T.; Wu, Z.; Pang, X. Prevention measures and countermeasures on brucellosis in Inner Mongolia. Neimenggu Prev. Med. 2000, 1, 136–139. [Google Scholar]
- Blyuss, K.B.; Kyrychko, Y.N. Stability and bifurcations in an epidemic model with varying immunity period. Bull. Math. Biol. 2010, 72, 490–505. [Google Scholar] [CrossRef] [Green Version]
- Okuwa, K.; Inaba, H.; Kuniya, T. An age-structured epidemic model with boosting and waning of immune status. Math. Biosci. Eng. 2021, 18, 5707–5736. [Google Scholar] [CrossRef]
- Gandolfi, A.; Pugliese, A.; Sinisgalli, C. Epidemic dynamics and host immune response: A nested approach. J. Math. Biol. 2015, 70, 399–435. [Google Scholar] [CrossRef] [Green Version]
- Gulbudak, H.; Cannataro, V.L.; Tuncer, N.; Martcheva, M. Vector-borne pathogen and host evolution in a structured immuno-epidemiological system. Bull. Math. Biol. 2017, 79, 325–355. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Dimension | Values | Sources | |
|---|---|---|---|---|
| b | birth rate | year | [24] | |
| Normalization parameter for shedding rate | CFU.year .animal | fitted | ||
| Mean duration of the latency period | year | [4,33] | ||
| Mean duration of the infectious period | year | 2 | [34] | |
| Density non-dependent mortality rate | year | [17,18] | ||
| Density dependent mortality rate | year | fitted | ||
| Mortality rate of virus | year | [10,17] | ||
| Horizontal direct transmission rate | CFU | [17,18] | ||
| Horizontal indirect transmission rate | CFU.year | fitted | ||
| p | Probability of disease transmission due to direct contact | - | assumed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kenne, C.; Mophou, G.; Dorville, R.; Zongo, P. A Model for Brucellosis Disease Incorporating Age of Infection and Waning Immunity. Mathematics 2022, 10, 670. https://doi.org/10.3390/math10040670
Kenne C, Mophou G, Dorville R, Zongo P. A Model for Brucellosis Disease Incorporating Age of Infection and Waning Immunity. Mathematics. 2022; 10(4):670. https://doi.org/10.3390/math10040670
Chicago/Turabian StyleKenne, Cyrille, Gisèle Mophou, René Dorville, and Pascal Zongo. 2022. "A Model for Brucellosis Disease Incorporating Age of Infection and Waning Immunity" Mathematics 10, no. 4: 670. https://doi.org/10.3390/math10040670
APA StyleKenne, C., Mophou, G., Dorville, R., & Zongo, P. (2022). A Model for Brucellosis Disease Incorporating Age of Infection and Waning Immunity. Mathematics, 10(4), 670. https://doi.org/10.3390/math10040670