Abstract
The increased mortality rates associated with antibiotic resistance has become a significant public health problem worldwide. Living beings produce a variety of endogenous compounds to defend themselves against exogenous pathogens. The knowledge of these endogenous compounds may contribute to the development of improved bioactive ingredients with antimicrobial properties, useful against conventional antibiotic resistance. Cowpea is an herbaceous legume of great interest due to its high protein content and high productivity rates. The study of genetic homology of vicillin (7S) from cowpea (Vigna unguiculata L.) with vicilins from soybean and other beans, such as adzuki, in addition to the need for further studies about potential biological activities of this vegetable, led us to seek the isolation of the vicilin fraction from cowpea and to evaluate the potential in vitro inhibitory action of pathogenic microorganisms. The cowpea beta viginin protein was isolated, characterized, and hydrolyzed in silico and in vitro by two enzymes, namely, pepsin and chymotrypsin. The antimicrobial activity of the protein hydrolysate fractions of cowpea flour was evaluated against Staphylococcus aureus and Pseudomonas aeruginosa, confirming the potential use of the peptides as innovative antimicrobial agents.
1. Introduction
The average mortality caused by bacterial resistance to antibiotics is expected to reach about 10 million people by 2050 globally, but with a higher rate in sub-Saharan African countries due to limited access to viable drugs [1]. In countries of higher income, antibiotic resistance results from the intensive and/or inappropriate use of antimicrobial drugs, triggering prominent multiresistant microorganisms. In addition, the report of new cases of resistance has been higher than the number of new drug substances with antibiotic activity that are being launched on the market [2].
Living beings produce a variety of substances against invasive pathogens. Improving the bioactive compounds from these organisms is therefore of great interest in searching for promising alternatives over conventional antibiotic drugs. In recent years, legume proteins have gained prominence, mainly due to the rapid expansion of knowledge about their bioactive peptides [3].
Peptides derived from legume seed proteins have been described to show various biological activities in vitro and in vivo, namely, with effects on the control of hunger [4], on the cardiovascular system [5,6], on inflammatory processes [7], cancer [8], and also with antimicrobial activity [9,10]. Antimicrobial peptides (AMPs) are a new class of biopharmaceuticals widely studied as important therapeutic alternatives [11].
AMPs are small molecules, playing an essential part of the defense system from various plant species [12]. These peptides are considered multifunctional since they show antibacterial, antifungal, antiparasitic, and antiviral properties, in addition to some antitumoral activity, capacity for insulin release, and immunomodulatory response mediated by cytokines [13]. Recently, several studies describe the action of peptides obtained from the digestion of legume proteins with antimicrobial properties [9,10]. There is growing interest in the production of food protein hydrolysates for potential therapeutic applications. Therefore, the ability of proteins to generate peptides during their gastrointestinal digestion, with favorable characteristics to reach the bloodstream, has been considered a fundamental condition [14]. Furthermore, the knowledge about the sequence of peptides is instrumental to understand the correlation between the composition of hydrolysates and their biological activity [14].
Peptides from the leguminous species Vigna unguiculata (L.) Walp, popularly known as cowpea, can be a natural and abundant source for the commercial production of antimicrobial peptides [15], since their dried seeds are a valuable source of proteins. The protein content of the seeds varies from 20 to 35%, with 7S globulins being the primary reserve proteins of cowpea [16,17]. Cowpea also has a genetic similarity with soybeans (64%) and adzuki beans (81%), and studies have shown that these legumes have significant antimicrobial, anticancer, antidiabetic, antioxidant, and hypocholesterolemic activities. In addition, V. unguiculata stands out for its low production cost, high nutritional value, and is abundant in the northeast region of Brazil [18]. The present study aimed to identify and characterize the beta vignin protein and its protein hydrolysates from cowpea with antimicrobial potential for use in pharmaceutics.
2. Materials and Methods
2.1. Materials
Cowpea (Vigna unguiculata, L. Walp) seeds were kindly providedby Dr. RogérioFaria Vieira (Agricultural Research Company from Minas Gerais at the Federal University of Viçosa, Minas Gerais, Brazil). All others reagents were bought fromSigma-Aldrich (San Luis, MO, USA).
2.2. Preparation of Cowpea Flour
The seeds were selected and soaked in distilled water at 4 °C for 12 h. Afterward, the seeds were dried in an oven at 50 °C for 12 h and powdered to 60 mesh size. The flour was stored at 4 °C and used for protein extraction.
2.3. Isolation of Cowpea β-Vignin
The β-vignin was isolated according to the methods described by Ferreira et al. (2015) [19]. Aliquots of the isolated protein (80 mg of β-vignin) were solubilized in potassium phosphate solution (0.05 M) at pH 7.5, NaCl (0.5 M), and sodium azide (0.01%), for a Sepharose CL-6B column (1.0 cm × 100 cm) with filtration. The flow rate was 0.45 mL/min, and the protein elution was monitored by measuring the absorbance at 280 nm. The major fraction (peak tube) was dialyzed, precipitated, and lyophilized. The protein concentration was determined as described by Lowry et al. (1951) [20].
2.4. Gel Electrophoresis
Samples of total protein extract and β-vignin isolated by chromatography were analyzed by one-dimensional sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE), as described by Laemlli (1970) [21], using an electrophoresis system HoeferMiniVE (Amersham Biosciences®, Hercules, CA, EUA). Aliquots of 10 μg of the sample were applied to the gel, and low molecular weight proteins (between 97 and 14.4 kDa) were used as molecular markers (GE Healthcare®, Little Chalfont, UK). The gel images were analyzed by the software AlphaEase® (Alpha Innotech, San Leandro, CA, USA).
2.5. Enzymatic Hydrolysis and Fractionation
β-vignin was hydrolyzed in vitro according to the procedure described by Akeson and Stahman (1964) [22]. Enzymatic hydrolysis was performed with pepsin and chymotrypsin. To perform pepsin (EC 34231) hydrolysis, a 1:66 enzyme/substrate ratio was used at 37 °C for 3 h (pH = 2.0); for chymotrypsin (EC3421), a 1:25 enzyme/substrate ratio was used at 37 °C for 3 h (pH 7.0). The total hydrolyzed extract was ultrafiltered from >30 kDa to >3 kDa (MWCO) using ultrafiltration membrane filters (Merck® Millipore, Darmstadt, Germany).
2.6. High-Performance Liquid Chromatography
The chromatographic profiles of the hydrolyzate of the fractions containing 30–10 kDa and 10–3 kDa peptides were determined by high-performance liquid chromatography (HPLC) using a PerkinElmer system with a reversed-phase column (C18 × 0.45 cm × 25 cm) and a UV/VIS detector (HPLC, PerkinElmer system, Waltham, MA, USA). The gradient was used for 10 min at 95% A and 50 min to reach 25% B. The solvent system comprised 0.045% trifluoroacetic acid in ultrapure water (A) and 0.036% trifluoroacetic acid in acetonitrile (B), with a flow rate of 1.0 mL/min at temperature of 30 °C. Readings were recorded at 220 nm.
2.7. In Silico Screening of Peptides with Antimicrobial Properties
The primary sequences of β-vignin (NCBI/GenBank Blast: AM905848 and UniProtKB: A8YQH5_VIGUN), adzuki bean 7S globulin (NCBI/GenBank Blouse: AB292246.1; UniProtKB: A4PI98_PHAAN), and α subunit of soybean β-conglycinin (NCBI)/GenBank Blast: AY221105.1; UniProtKB: UniProtKB: GLCAP_SOYBN) were compared with protein modeling software [23]. Then, the β-vignin primary sequence was virtually hydrolyzed by the sequential action of the enzymes pepsin (EC 3.4.23.1) and chymotrypsin (EC 3.4.21.1), as available on the BIOPEP server [24]. Subsequently, the probability of bioactivity of β-vignin-derived peptides was analyzed according to the physicochemical characteristics that were presented.
2.8. Minimum Inhibitory Concentration
Minimum inhibitory concentration (MIC) assays for the peptides were performed using the microdilution technique following the National Committee for Clinical Laboratory Standards guidelines [25]. Strains of the Staphylococcus aureus (ATCC 25923) and Pseudomonas aeruginosa (ATCC27853) were used. Colonies were harvested and resuspended to 1.5 × 108 CFU/mL (turbidity equivalent to 0.5 McFarland standard scale). Samples of hydrolyzed proteins and total proteins were diluted in dimethyl sulfoxide at concentrations ranging from 1.0 through 0.0019531 mg/mL, and added to Mueller–Hinton broth (Merck, Darmstadt, Germany). The negative control was 0.1 mL of Mueller–Hinton broth, and the positive control was ciprofloxacin (Merck, Darmstadt, Germany). Plates were incubated at 37 °C for 24 h. At the end of the incubation time, MIC was visually identified as the lowest concentration of the test compound that inhibits visible growth.
2.9. Agar Disk Diffusion Method
The agar disk diffusion method was carried out according to the National Committee for Clinical Laboratory Standards guidelines [26]. Bacterial suspensions were cultured in Mueller–Hinton broth for 24 h at 35 °C, standardized in sterile saline solution (0.9%) at a concentration of 108 CFU/mL, a 0.5 McFarland standard. The strains were sown with the sterile swab. After 10 min, three holes were made on the surface of the inoculated medium using light pressure, and the peptides were inserted according to their minimum inhibitory concentration values. The plates were incubated in a bacteriological oven at 37 °C for 24 h. The antimicrobial activity results of the tested sample were expressed through the diameter size of the inhibition halo.
2.10. Cell Viability in L929 Cell-Line
The human fibroblast line-L929 was used for the colorimetric method using methyl-thiazolyl-tetrazolium (MTT assay), following the ISO 10993-5 (2009) guidelines [27]. L929 cells were seeded in 96-well culture plates (2 × 10⁴ cells/well). A solution of MTT was placed in contact with the cells, and then incubated at 37 °C for 3 h. After removal of the MTT, dimethyl sulfoxide was placed for solubilization of the tetrazole salt crystals. Then, the optical density reading was performed on an automated plate reader at 570 nm wavelength. The tests were conducted in quadruplicate and then normalized [28]. The results are expressed as a relative percentage of cell viability compared to the control, calculated by applying the following equation:
where CV (%) is the percentage of cell viability, is the absorbance recorded for the sample, and is the absorbance recorded for the control.
3. Results
The spectrochromatographic profile of the cowpea 7S protein isolate on the Sepharose CL 6B column showed a single absorbance peak in tube 78 (Figure 1). Electrophoresis indicated that 7S globulins represent the major proteins of cowpea, being composed of bands corresponding to two significant polypeptides (55 to 60 kDa). Other smaller bands were registered by polyacrylamide gel electrophoresis of the protein fractions (Figure 2). The recorded data ensured the identification and quantification the β-vignin protein, resulting in a 50% yield of the 7S fraction of cowpea extract.
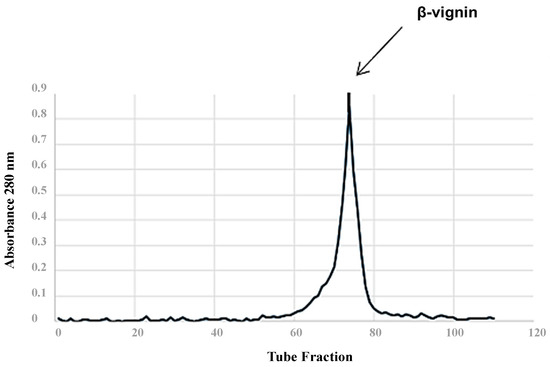
Figure 1.
Spectrochromatographic profile of cowpea 7S protein isolate on a Sepharose CL 6B column.

Figure 2.
Polyacrylamide gel electrophoresis of protein fractions from cowpea. The columns represent the protein marker (A), the total protein extract (B), the defatted and purified beta vignin (C), and finally, the enzymatic hydrolysis (D,E).
The chromatographic profiles of the protein hydrolyzate of the 30–10 kDa and 10–3 kDa fractions of chymotrypsin (A and B) and in vitro pepsin hydrolyzate (C and D) of beta vignin are shown in Figure 3. The registered peaks are the indication of peptides present in samples under study. Although many peptides were produced by this protocol (Table 1), few significant peaks were observed in the 30–10 kDa hydrolysates for both enzymes. There was higher peak intensity in this case, but the retention times were the same. For hydrolyzates < 30 kDa, the test was not performed since the column would not support this molecular weight, causing the clogging of the column.

Figure 3.
Chromatogram of RP-HPLC chymotrypsin hydrolyzate of the fraction containing 30–10 kDa peptides (A); hydrolyzate of the fraction containing 10–3 kDa smaller peptides (B); pepsin hydrolyzate fraction containing 30–10 kDa smaller peptides (C), and hydrolysate fraction containing smaller peptides 10–3 kDa (D).

Table 1.
Quantification of the fraction consisting of peptides larger than 30, 30–10, and 10–3 kDa.
The computer simulation of enzymatic hydrolysis performed on cowpea beta vignin produced hundreds of peptide fragments. Most predicted bioactive peptides (Table 2 and Table 3) have between 10 and 30 amino acid residues, molecular mass from 1 to 5 kDa, with hydrophobicity values ranging from 0.9 to −0.1 kcal, positive charge ranging from +2 to 4.1, and predominance of the hydrophobic residues, namely, phenylalanine, tyrosine, or leucine. According to the simulation, the peptides that presented the most favorable characteristics and prediction for antimicrobial activity were those numbered 7 and 21 (Table 2) for protein hydrolyzates with chymotrypsin, and peptides numbered 19 and 21 (Table 3) for protein hydrolyzates with pepsin.
Table 2.
Screening prediction of β-vignin-derived peptides from cowpea hydrolyzed with chymotrypsin in silico and their characteristics of isoelectric point, molecular mass, and high-performance chromatography time.
Table 3.
Screening prediction of β-vignin-derived peptides from cowpea hydrolyzed with pepsin in silico and their characteristics of isoelectric point, molecular mass, and high-performance chromatography time.
These MIC values were recorded against two main strains, selected on the basis of commonly reported clinical diagnostic infections, as examples of Gram-positive (Staphylococcus aureus) and Gram-negative (Pseudomonas aeruginosa) bacteria. Staphylococcus aureus is one of the most common human pathogens and Pseudomonas aeruginosa is a common cause of nosocomial pneumonia, urinary tract infection, and surgical site infection. It has become a less frequent cause of bacteremia in patients with neutropenia in most parts of the world, but remains the most important pathogen in patients with cystic fibrosis. The MIC values for all peptide hydrolyzates were 512 µg/mL against Pseudomonas aeruginosa and Staphylococcus aureus (Table 4). These results shown in Table 2 do not depict the fraction of peptides with the highest antimicrobial activity since the hydrolyzates presented the same MIC. However, when evaluated by the disk diffusion test, the protein hydrolyzates of both enzymes showed bacterial inhibition and dose-dependent effect against the tested strains (Table 5). In general, larger inhibition halos were observed for pepsin hydrolyzates for Staphylococcus aureus (with a diameter of 11.11 mm at a concentration of 250 µg/mL and 12.1 mm at a concentration of 500 µg/mL) and Pseudomonas aeruginosa at a concentration of 500 µg/mL (10.9 mm in diameter).
Table 4.
MIC of fractionated protein hydrolysates of cowpea 7S globulin against Pseudomonas aeruginosa and Staphylococcus aureus. Results are expressed as (µg/mL) the mean ± standard deviation (n = 4).
Table 5.
Antimicrobial activity of cowpea beta vignin total protein hydrolyzate against Gram-positive and Gram-negative bacteria indicated by halo inhibition (mm) at two concentrations.
The inhibitory effect of the derivatives of the hydrolysis of beta vignin with pepsin on the proliferation of mammalian cells at a concentration of 512 µg/mL was greater than 80%, indicating biocompatibility (Figure 4A). However, for the chymotrypsin hydrolysates (Figure 4B) at a concentration of 512 µg/mL, the sample showed cytotoxicity (48% cell viability). For the other concentrations of chymotrypsin hydrolysates, cytotoxicity can be considered insignificant for mammalian cells.

Figure 4.
Evaluation of the viability of L929 human fibroblasts determined by the MTT assay after 24 h of incubation in pepsin (A) and chymotrypsin (B) hydrolysates.
4. Discussion
Globins constitute about 80% of the total protein in cowpea [29]. Studies indicate that this protein fraction is formed mainly by α-, β-, and γ-vignin proteins. However, in our study we found 7S globulins (β-vignina), the main proteins in cowpea, with a yield similar to that reported in the literature [6,19].
Our study demonstrated that the total globulin fraction consists of eight polypeptide chains (Figure 2B). However, the β-vignin protein obtained by chromatography had three polypeptide chains (Figure 2C), comprising two main glycosylated polypeptide chains with 50 and 55 kDa molecular weights. According to the literature, β-vignin is composed of two main chains of glycosylated polypeptides with molecular weights of 60 and 55 kDa and other smaller chains [30]. However, under denaturing and reducing conditions, cowpea vicilins are a heterogeneous mixture of polypeptides of various sizes [19,29,31]. Molecular mass analysis by electrophoresis showed a difference between the values calculated from the amino acid sequences. These differences are attributed to the glycosylation that these polypeptides undergo in the post-translational processing of their precursors [17].
The computer simulation of enzymatic hydrolysis performed on beta vignin from cowpea produced hundreds of peptide fragments, most of them with a molecular mass of 1 to 5 kDa. Although expected, this same pattern was not observed when this enzymatic proteolysis was performed in vitro. There was a higher amount of hydrolyzates with molecular mass greater than 30 kDa when compared to the other fractions. However, some peaks in the chromatographic samples were seen, showing many peptides within the samples under study. Several review studies do highlight the therapeutic potential of food-derived bioactive peptides, which have an antimicrobial function [32]. In our work, we used bacterial strains of Pseudomonas aeruginosa and Staphylococcus aureus to determine the MIC. The MIC value for all hydrolyzates was the same (512 µg/mL) when evaluated in both bacterial strains. There is no consensus on the acceptable standard for hydrolyzed protein isolates of legume proteins compared to conventional antibiotics. Some authors consider results only similar to known antibiotics, provided they work with a fraction already determined. In the present study, we did not work with the predetermined peptide fraction of beta vignin of cowpea. We followed the criteria suggested by Holetz et al. (2002) [33] and Carvalho et al. (2014) [34], who consider that a MIC below 100 mg/mL has appropriate antimicrobial activity and while concentrations above 500 mg/mL have poor activity and are difficult to use in the treatment of bacterial infections.
The antimicrobial activity of peptides is primarily based on the interaction between the peptide structure and the microorganism’s cell membrane [35]. Because of their cationic or amphiphilic character, peptides bind to lipid membranes because of the attraction of the arginine and lysine residues of the peptide structure to the phospholipids present in the bilayer and through the interaction between the hydrophobic amino acid residues of the peptide and the membrane. These interactions allow the peptides to cross the lipid bilayer and reach the inner side of the cell, causing membrane dysfunction through the formation of pores with extravasation of ions and metabolites, depolarization, loss of membrane coupled respiration, and, ultimately, cell death [35,36,37].
Previous studies have highlighted the difficulty in identifying bioactive peptides derived from enzymatic proteolysis due to the significant variability of the primary sequences that were found [38,39]. Therefore, an in silico simulation was performed, during which it was possible to identify four hydrolyzates that could show more favorable characteristics and prediction for antimicrobial activity. These hydrolyzates are small sequences with a mass of less than 3 kDa. However, it is worth noting that a pool of peptides that were not yet isolated was used in our study, and they may be modulating the antimicrobial activity evaluated against S. aureus and P. aeruginosa. The dissociation of these peptides into even smaller fractions is necessary to identify and isolate the peptide with the best antimicrobial activity [40].
In addition, it is worth considering that Pseudomonas aeruginosa species becomes increasingly difficult to control due to a diversity of intrinsic and acquired drug-resistance mechanisms [41]. There are even records in the literature of resistance to some antimicrobial peptides against Gram-negative bacteria [42].
For the cytotoxicity evaluation, the MTT test was used against fibroblast cell line-L929 as recommended in the ISO 10993/2009 guideline [27]. Additionally, the L929 is highly proliferative and is widely used in cytotoxicity testing, mainly to check toxicity toward cellular viability and proliferation. Fibroblast cells are the most common cells of all types of connective tissues, being actively engaged in the synthesis and upkeep of the collagenous extracellular matrix, and also modulating adjacent cell behavior, including migration, proliferation, and differentiation. In this way, biological evaluation with fibroblast cell cultures might be regarded as a general bioassay, providing reliable information concerning basal cytotoxicity. The results showed that the hydrolyzed fractions of beta vignin from cowpea generally have low cytotoxicity (Figure 4). Thus, the peptide fractions used in the present work are safe and show antimicrobial potential. The unfolding is exactly in the sequencing and identification of the major peptides of this fraction, bearing in mind that the MIC of beta vignin protein hydrolyzates showed satisfactory concentrations.
5. Conclusions
Cowpea protein hydrolyzatesare shownto be safe and can be considered a potential alternative for developing innovative antimicrobials. Further studies of the peptide composition on amino acid sequences of beta vignin are needed to understand the structure–activity relationships of these peptides to elucidate their antimicrobial mechanisms of action and possible applicability in the market as new biotechnological drugs for human health or in the construction of transgenic plants with resistance to pathogens.
Author Contributions
T.S.A., C.A.d.C.S., M.B.d.C.e.S., F.P.R.B., E.S.F., A.L.S.S., L.N.S. and C.R.M. contributed in the conceptualization, methodology, validation, formal analysis, investigation, and writing—original draft preparation. C.B., M.L.B., J.C.C., R.L.C.d.A.-J., R.d.M.B., M.M.P., E.B.S., C.M.F.S. and P.S. contributed for the methodology, supervision, writing—review and editing, project administration, resources, and funding acquisition. All authors have contributed substantially to the work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CNPq, FAPERJ and CAPES (financial code-001), and by the Fundação Carolina (Movilidad de profesoradoBrasil-España, Movilidad. Estancias de Investigación, C.2020) granted to P.S.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Brown, D. Antibiotic resistance breakers: Can repurposed drugs fill the antibiotic discovery void? Nat. Rev. Drug Discov. 2015, 14, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Okoye, C.O.; Ezeorba, T.P.C.; Okeke, E.S.; Okagu, I.U. Recent Findings on the Isolation, Identification and Quantification of Bioactive Peptides. Appl. Food Res. 2022, 2, 100065. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Rupasinghe, S.G.; Schuler, M.A.; de Mejia, E.G. Peptides from purified soybean beta-conglycinin inhibit fatty acid synthase by interaction with the thioesterase catalytic domain. FEBS J. 2010, 277, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Two Peptides from Soy β-Conglycinin Induce a Hypocholesterolemic Effect in HepG2 Cells by a Statin-Like Mechanism: Comparative in Vitro and in Silico Modeling Studies. J. Agric. Food Chem. 2015, 63, 7945–7951. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.R.; Fontanari, G.G.; Pimenta, D.C.; Soares-Freitas, R.M.; Arêas, J.A.G. Proteolytic hydrolysis of cowpea proteins is able to release peptides with hypocholesterolemic activity. Food Res. Int. 2015, 77, 43–48. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Food Biochem. 2019, 43, e12531. [Google Scholar] [CrossRef]
- Moreno, C.; Mojica, L.; González de Mejía, E.; Camacho Ruiz, R.M.; Luna-Vital, D.A. Combinations of Legume Protein Hydrolysates Synergistically Inhibit Biological Markers Associated with Adipogenesis. Food 2020, 9, 1678. [Google Scholar] [CrossRef]
- Farkas, A.; Maróti, G.; Kereszt, A.; Kondorosi, É. Comparative Analysis of the Bacterial Membrane Disruption Effect of Two Natural Plant Antimicrobial Peptides. Front. Microbiol. 2017, 8, 51. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Sánchez-Pérez, A.; Calo-Mata, P.; Villa, T.G. Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria. Biochem. Pharmacol. 2017, 133, 117–138. [Google Scholar] [CrossRef]
- Luong, H.X.; Thanh, T.T.; Tran, T.H. Antimicrobial peptides—Advances in development of therapeutic applications. Life Sci. 2020, 260, 118407. [Google Scholar] [CrossRef] [PubMed]
- Benko-Iseppon, A.M.; Galdino, S.L.; Calsa, T., Jr.; Kido, E.A.; Tossi, A.; Belarmino, L.C.; Crovella, S. Overview on plant antimicrobial peptides. Curr. Protein Pept. Sci. 2010, 11, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Mechkarska, M.; Lukic, M.L.; Flatt, P.R. Potential therapeutic applications of multifunctional host-defense peptides from frog skin as anti-cancer, anti-viral, immunomodulatory, and anti-diabetic agents. Peptides 2014, 57, 67–77. [Google Scholar] [CrossRef]
- Aiello, G.; Lammi, C.; Boschin, G.; Zanoni, C.; Arnoldi, A. Exploration of Potentially Bioactive Peptides Generated from the Enzymatic Hydrolysis of Hempseed Proteins. J. Agric. Food Chem. 2017, 65, 10174–10184. [Google Scholar] [CrossRef]
- Osman, A.; Enan, G.; Al-Mohammadi, A.-R.; Abdel-Shafi, S.; Abdel-Hameid, S.; Sitohy, M.Z.; El-Gazzar, N. Antibacterial Peptides Produced by Alcalase from Cowpea Seed Proteins. Antibiotics 2021, 10, 870. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Guan, N.; Li, P.; Li, J.; Luo, J. Monitoring and dietary exposure assessment of pesticide residues incowpea (Vigna unguiculata L. Walp) in Hainan, China. Food Control 2016, 59, 250–255. [Google Scholar] [CrossRef]
- Rocha, A.J.; Sousa, B.L.; Girão, M.S.; Barroso-Neto, I.L.; Monteiro-Júnior, J.E.; Oliveira, J.T.A.; Nagano, C.S.; Carneiro, R.F.; Monteiro-Moreira, A.C.O.; Rocha, B.A.M.; et al. Cloning of cDNA sequences encoding cowpea (Vigna unguiculata) vicilins: Computational simulations suggest a binding mode of cowpea vicilins to chitin oligomers. Int. J. Biol. Macromol. 2018, 117, 565–573. [Google Scholar] [CrossRef]
- Pina-Pérez, M.C.; Ferrús Pérez, M.A. Antimicrobial potential of legume extracts against foodborne pathogens: A review. Trends Food Sci. Technol. 2018, 72, 114–124. [Google Scholar] [CrossRef]
- Ferreira, E.S.; Amaral, A.L.S.; Demonte, A.; Zanelli, C.F.; Capraro, J.; Duranti, M.; Neves, V.A. Hypocholesterolaemic effect of rat-administered oral doses of the isolated 7S globulins from cowpeas and adzuki beans. J. Nutr. Sci. 2015, 4, e7. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Akeson, W.R.; Stahmann, M.A. A pepsin pancreatin digest index of protein quality evaluation. J. Nutr. 1964, 83, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2022, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci 2019, 20, 5978. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Antimicrobial Broth Dilution and Disk Diffusion Susceptibility Testing of Bacteria Isolated From Aquatic Animals, 2nd ed.; CLSI Guideline VET03; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; CLSI Standard M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- ISO 10993-1:2009; Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. British Standards: London, UK, 2009.
- Severino, P.; Chaud, M.V.; Shimojo, A.; Antonini, D.; Lancelloti, M.; Santana, M.H.A.; Souto, E.B. Sodium alginate-cross-linked polymyxin B sulphate-loaded solid lipid nanoparticles: Antibiotic resistance tests and HaCat and NIH/3T3 cell viability studies. Colloids Surf. B Biointerfaces 2015, 129, 191–197. [Google Scholar] [CrossRef]
- Aluko, R.E.; Girgih, A.T.; He, R.; Malomo, S.; Li, H.; Offengenden, M.; Wu, J. Structural and functional characterization of yellow field pea seed (Pisum sativum L.) protein-derived antihypertensive peptides. Food Res. Int. 2015, 77, 10–16. [Google Scholar] [CrossRef]
- Freitas, R.L.; Teixeira, A.R.; Ferreira, R.B. Characterization of the proteins from Vigna unguiculata seeds. J. Agric. Food Chem. 2004, 52, 1682–1687. [Google Scholar] [CrossRef]
- Gonçalves, A.; Goufo, P.; Barros, A.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef]
- Awika, J.M.; Duodu, K.G. Bioactive polyphenols and peptides in cowpea (Vigna unguiculata) and their health promoting properties: A review. J. Funct. Foods 2017, 38, 686–697. [Google Scholar] [CrossRef]
- Holetz, F.B.; Pessini, G.L.; Sanches, N.R.; Cortez, D.A.G.; Nakamura, C.V.; Dias Filho, B.P. Screening of some plants used in the Brazilian folk medicine for the treatment of infectious diseases. Memórias Instituo Oswaldo Cruz 2002, 97, 1027–1031. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Silva, D.M.I.; Silva, T.R.C.; Scarcelli, E.; Manhani, M.R. Evaluation of the antibacterial activity of ethanolic and cyclohexane extracts of chamomile flowers (Matricaria chamomilla L.). Rev. Bras. Plantas Med. 2014, 16, 521–526. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and consequences of bacterial resistance to antimicrobial peptides. Drug Resist. Updates 2016, 26, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Bahar, A.A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [PubMed]
- Salas, C.E.; Badillo-Corona, J.A.; Ramírez-Sotelo, G.; Oliver-Salvador, C. Biologically active and antimicrobial peptides from plants. BioMed Res. Int. 2015, 2015, 102129. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Souza, C.; Philadelpho, B.O.; Cunha, M.; Batista, F.P.R.; Silva, J.R.D.; Druzian, J.I.; Castilho, M.S.; Cilli, E.M.; Ferreira, E.S. In vitro and in silico studies of 3-hydroxy-3-methyl-glutaryl coenzyme A reductase inhibitory activity of the cowpea Gln-Asp-Phe peptide. Food Chem. 2018, 259, 270–277. [Google Scholar] [CrossRef]
- Fassini, P.G.; Noda, R.W.; Ferreira, E.S.; Silva, M.A.; Neves, V.A.; Demonte, A. Soybean glycinin improves HDL-C and suppresses the effects of rosuvastatin on hypercholesterolemic rats. Lipids Health Dis. 2011, 10, 165. [Google Scholar] [CrossRef]
- Xiang, N.; Lyu, Y.; Zhu, X.; Bhunia, A.K.; Narsimhan, G. Methodology for identification of pore forming antimicrobial peptides from soy protein subunits β-conglycinin and glycinin. Peptides 2016, 85, 27–40. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Shai, Y. Short native antimicrobial peptides and engineered ultrashort lipopeptides: Similarities and differences in cell specificities and modes of action. Cell. Mol. Life Sci. 2011, 68, 2267–2280. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).