
Assessing Role of Drought Indices in Anticipating Pine Decline in the Sierra Nevada, CA



Abstract
1. Introduction
2. Materials and Methods
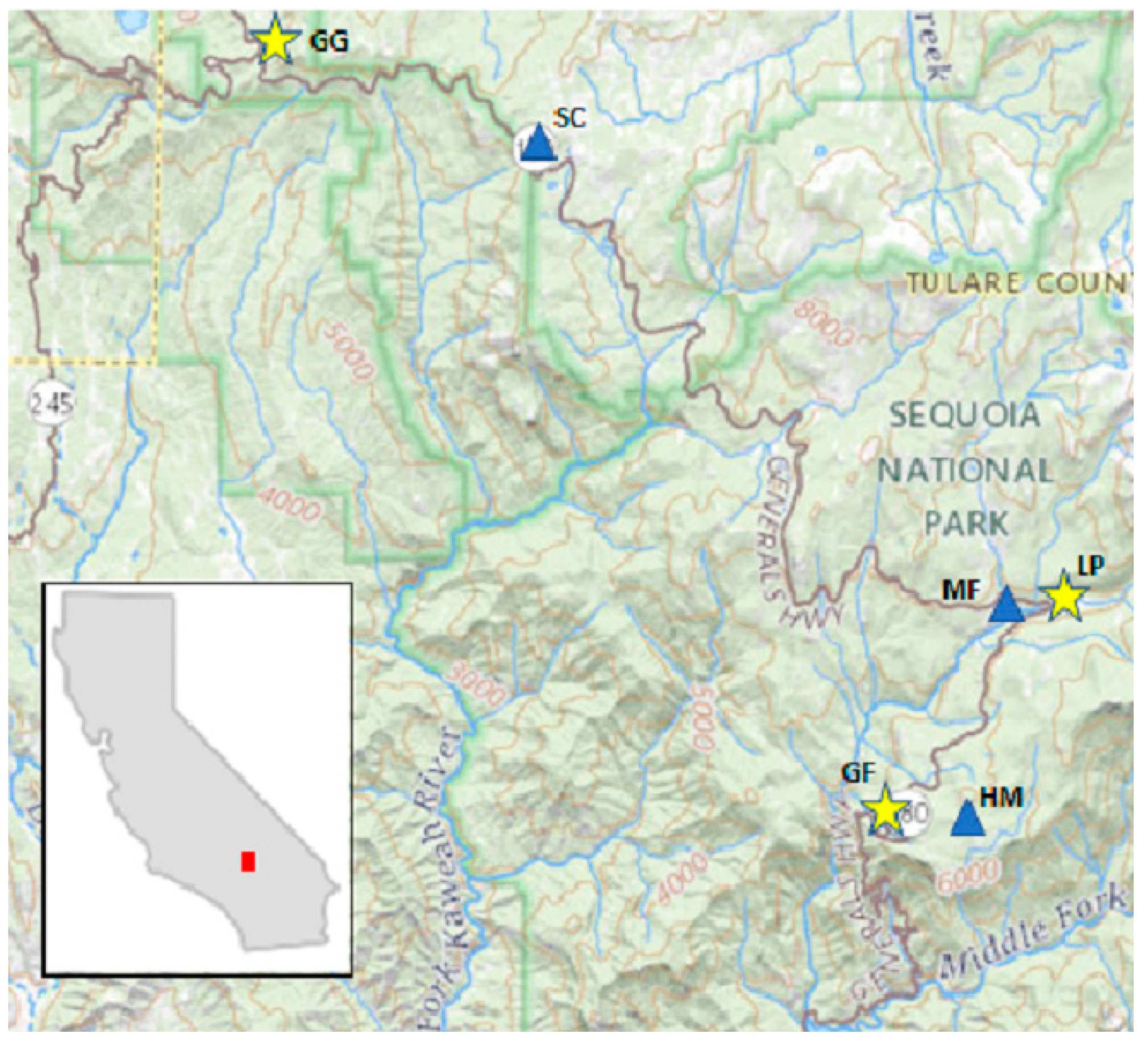
2.1. Study Site Description
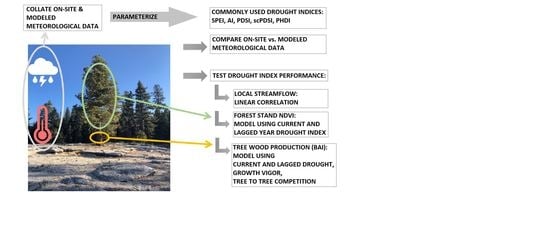
2.2. Work Flow
2.3. Collating Meteological Data
2.4. Parameterization of Drought Indices
2.5. Independent Tests of DI Effectiveness
2.5.1. Streamflow
2.5.2. NDVI
2.5.3. BAI
3. Results
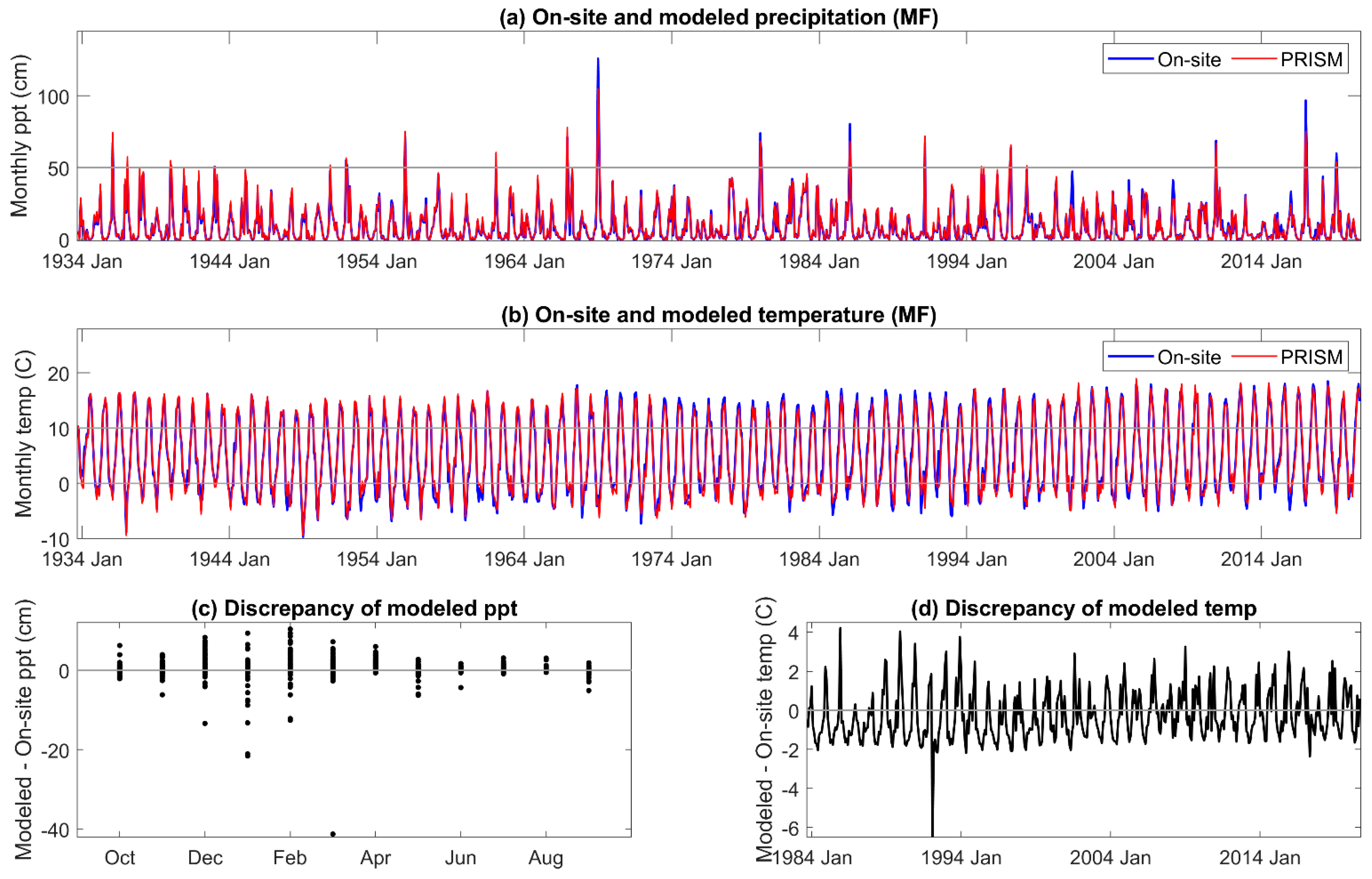
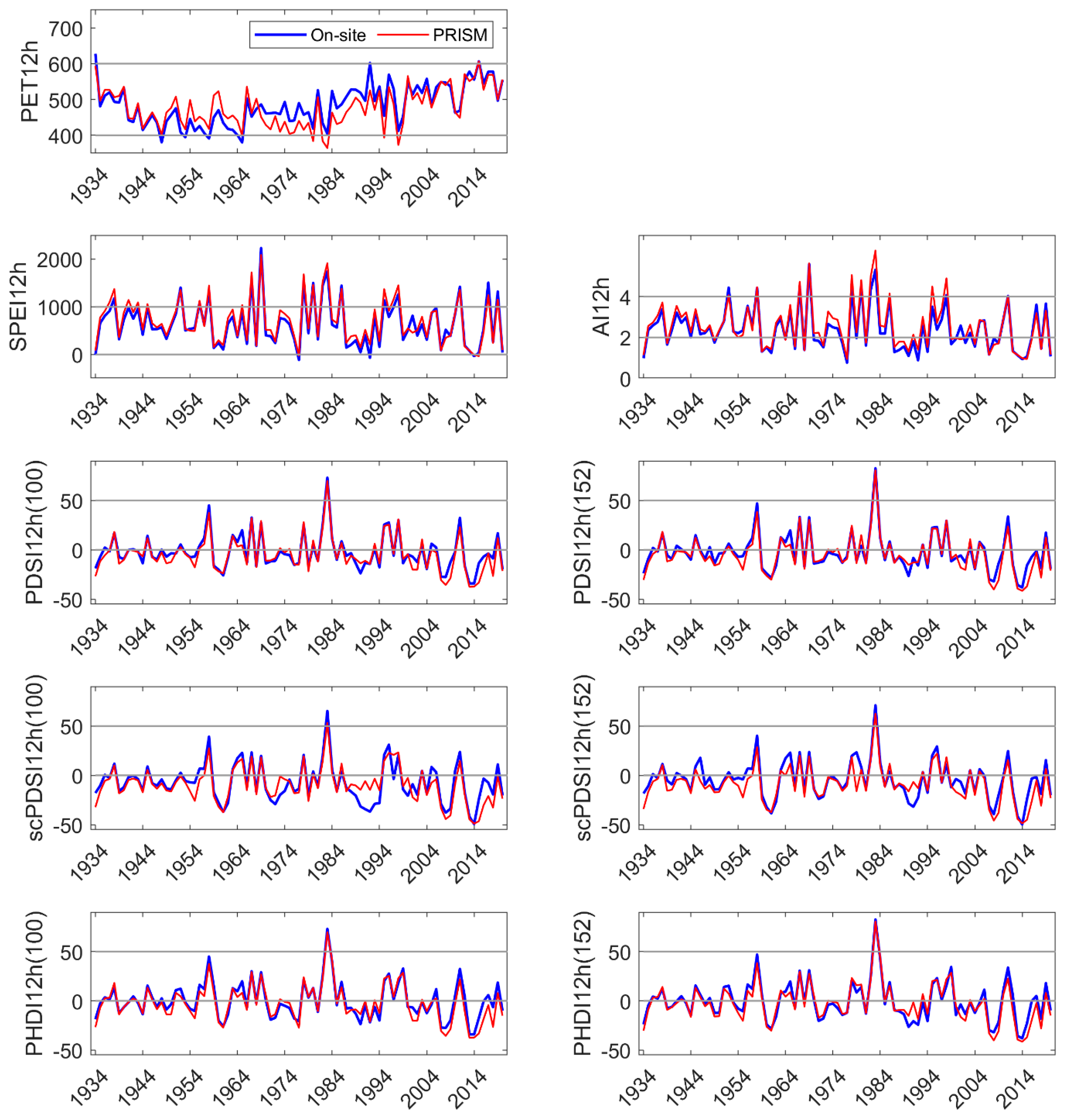
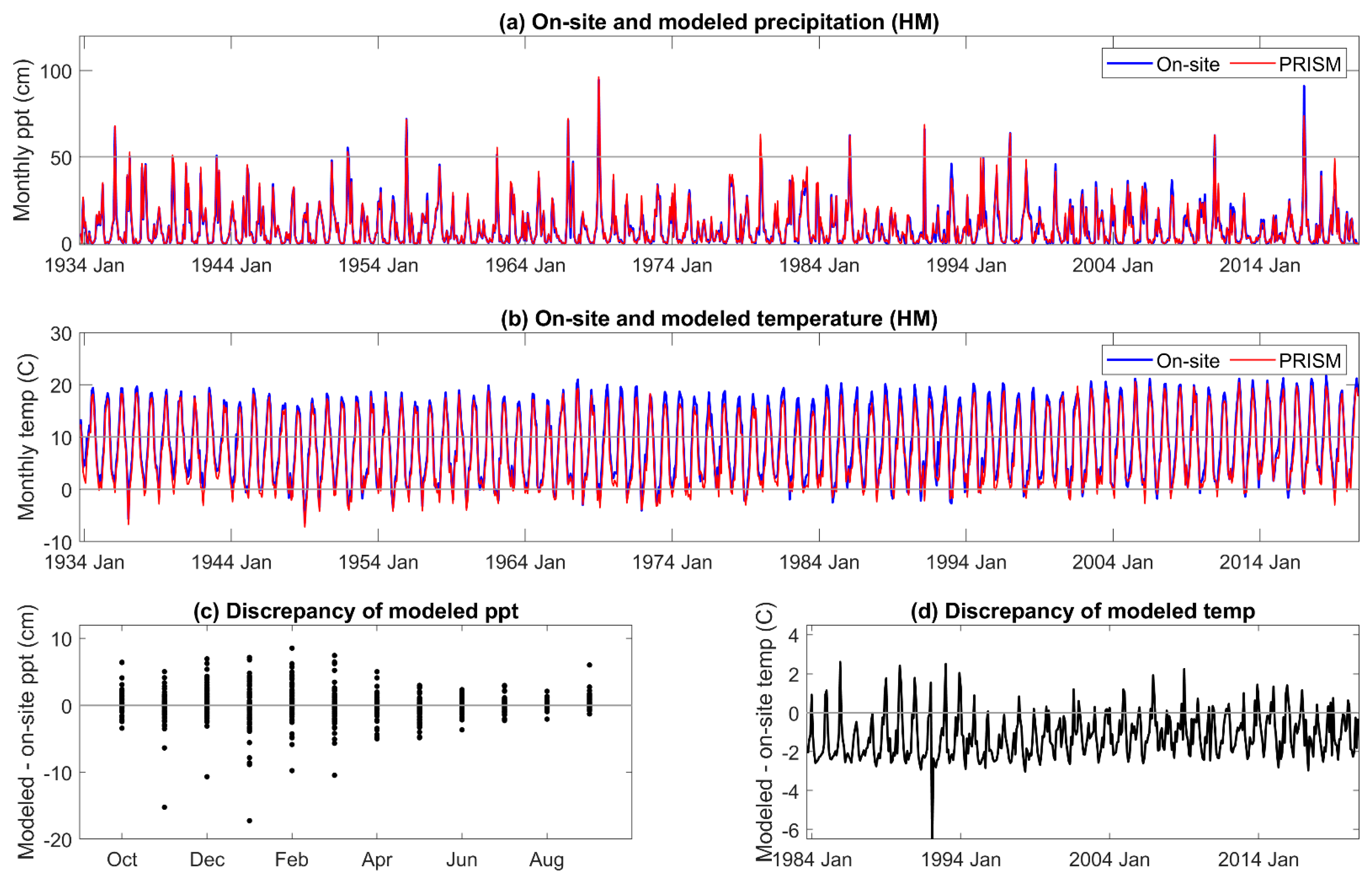
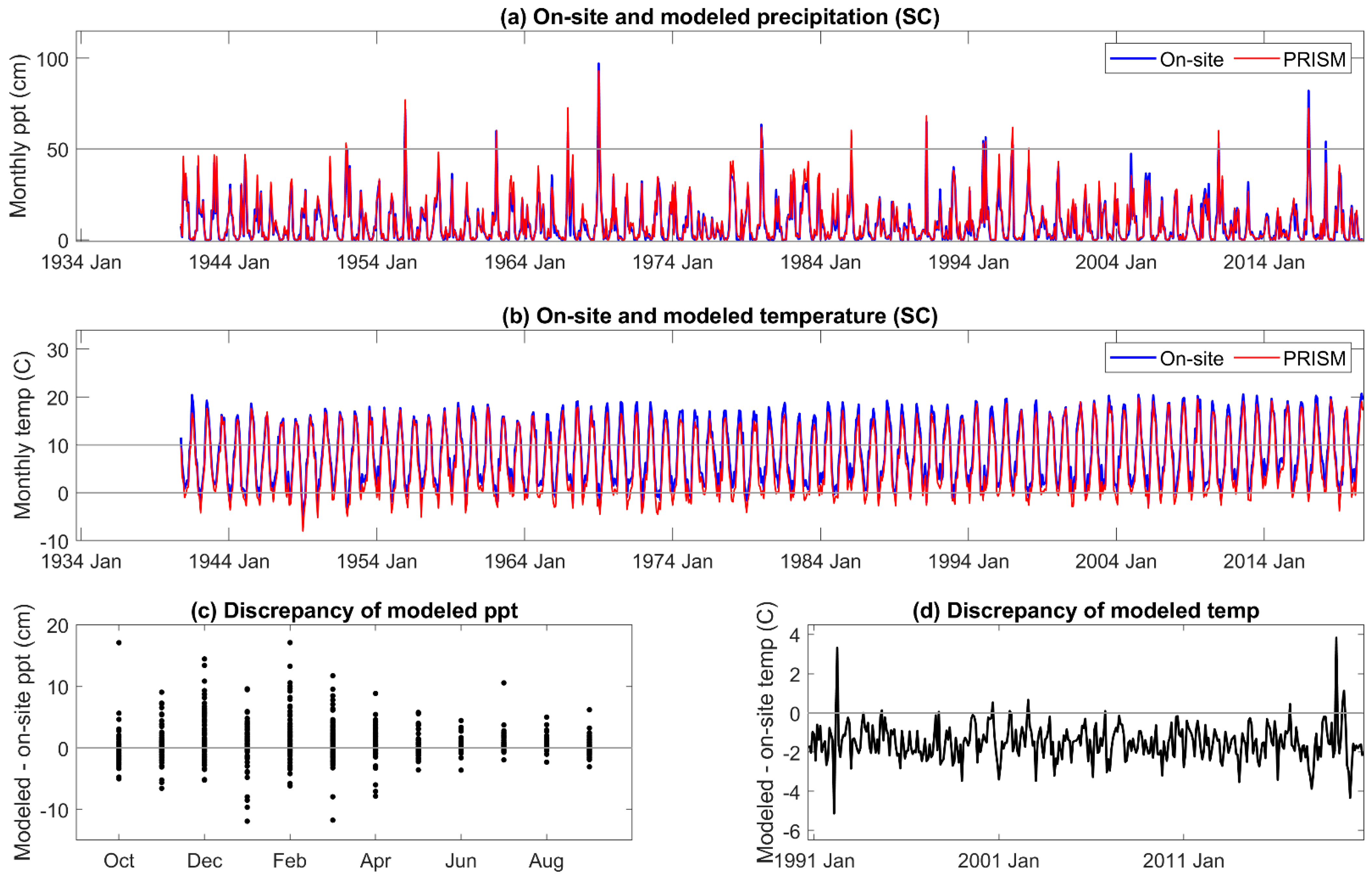
3.1. On-Site vs. PRISM Meteorology
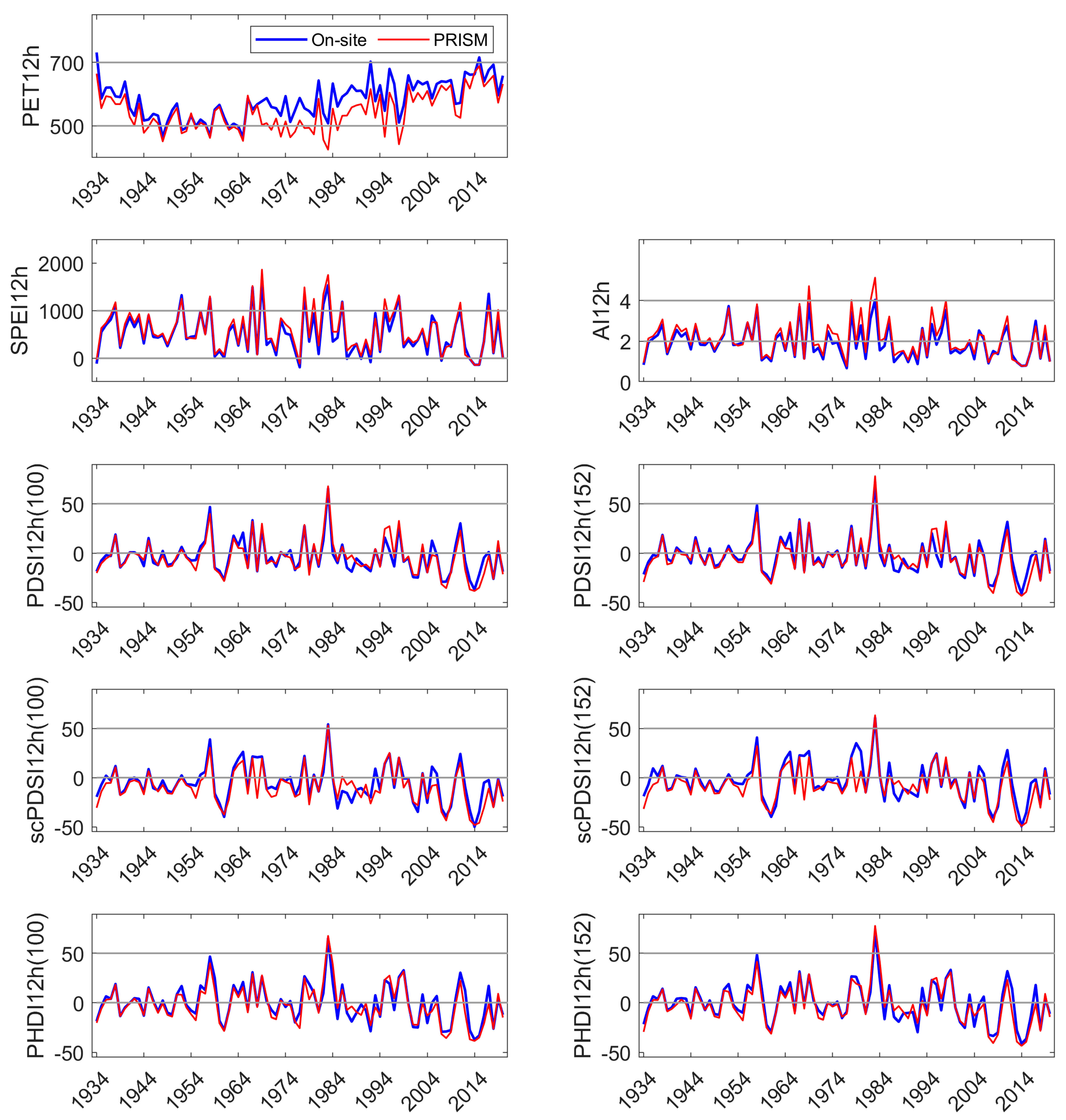
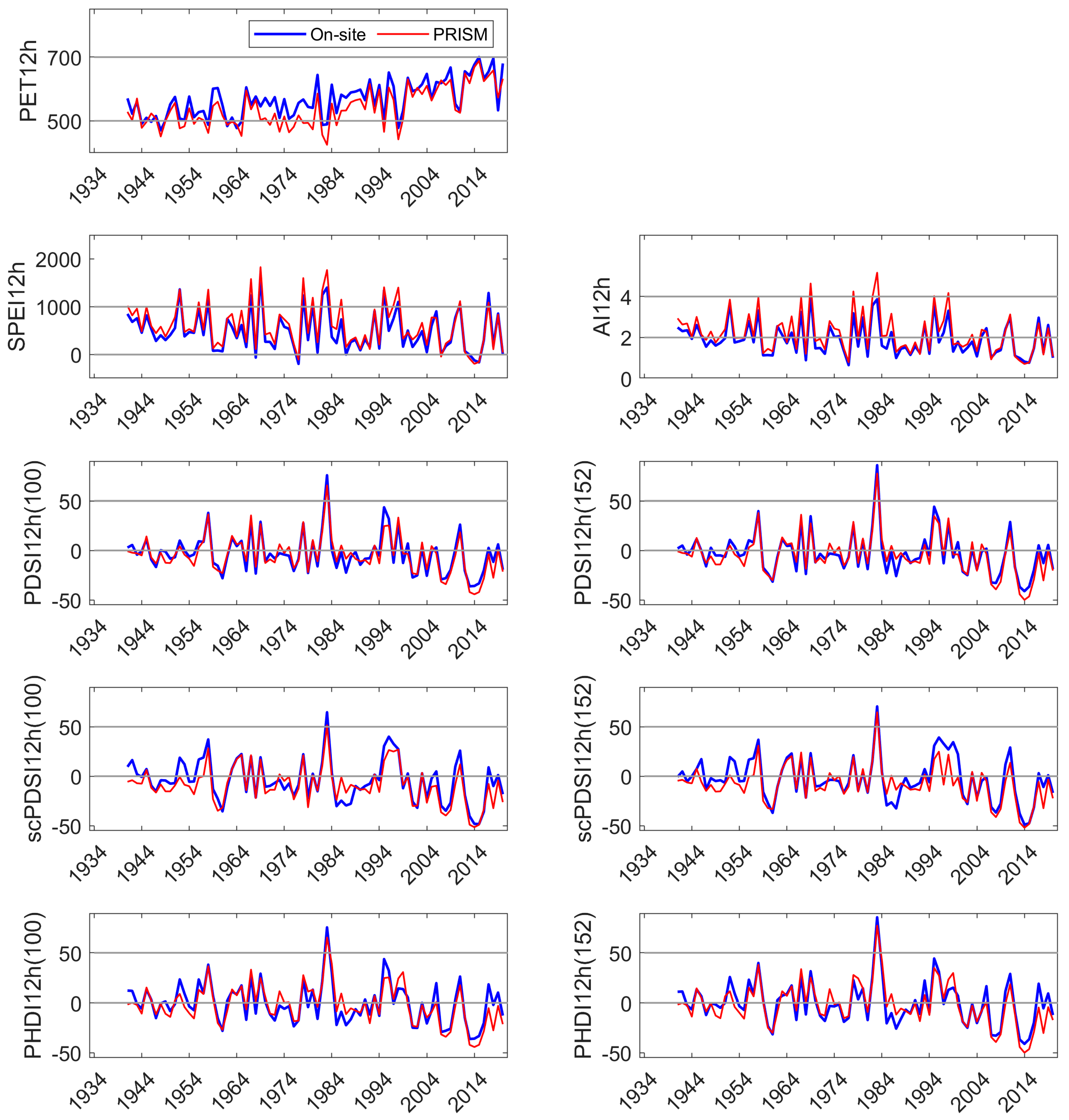
3.2. On-Site vs. PRISM Parameterization of Drought Indices
3.3. Correlation among Drought Indices
3.4. Drought Index Performance Relative to Short-Term Streamflow
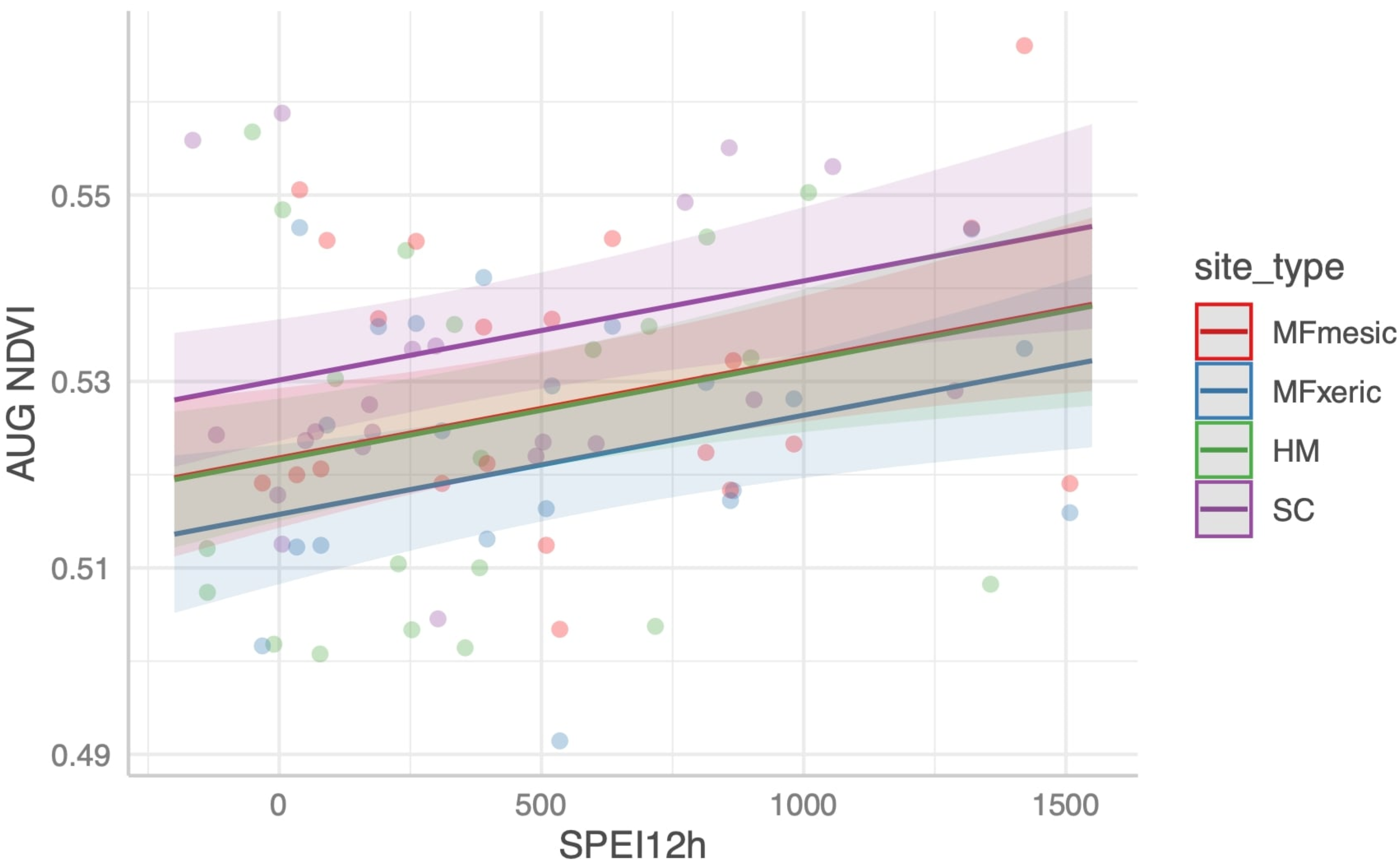
3.5. Drought Index Performance Predicting Site NDVI
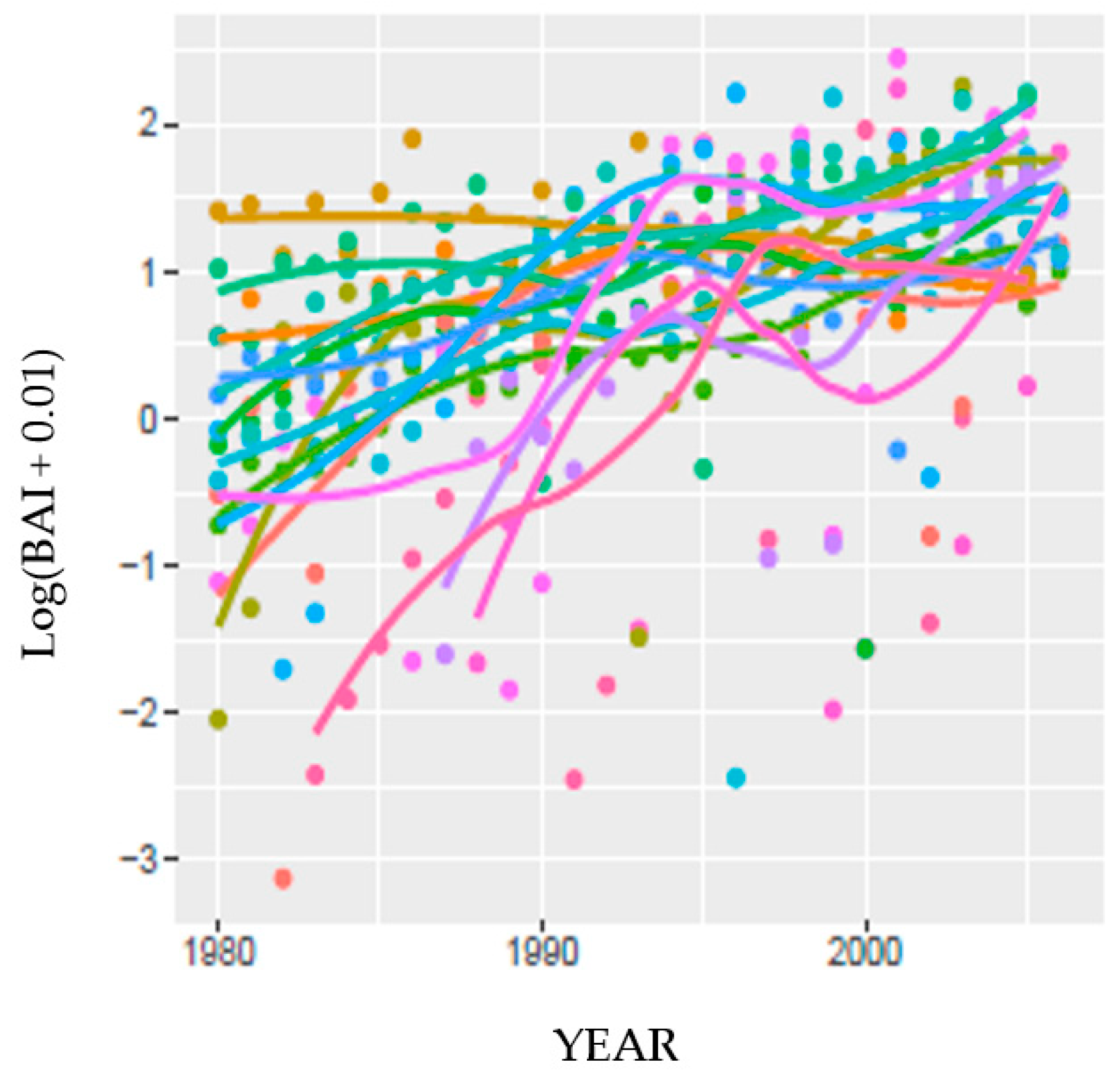
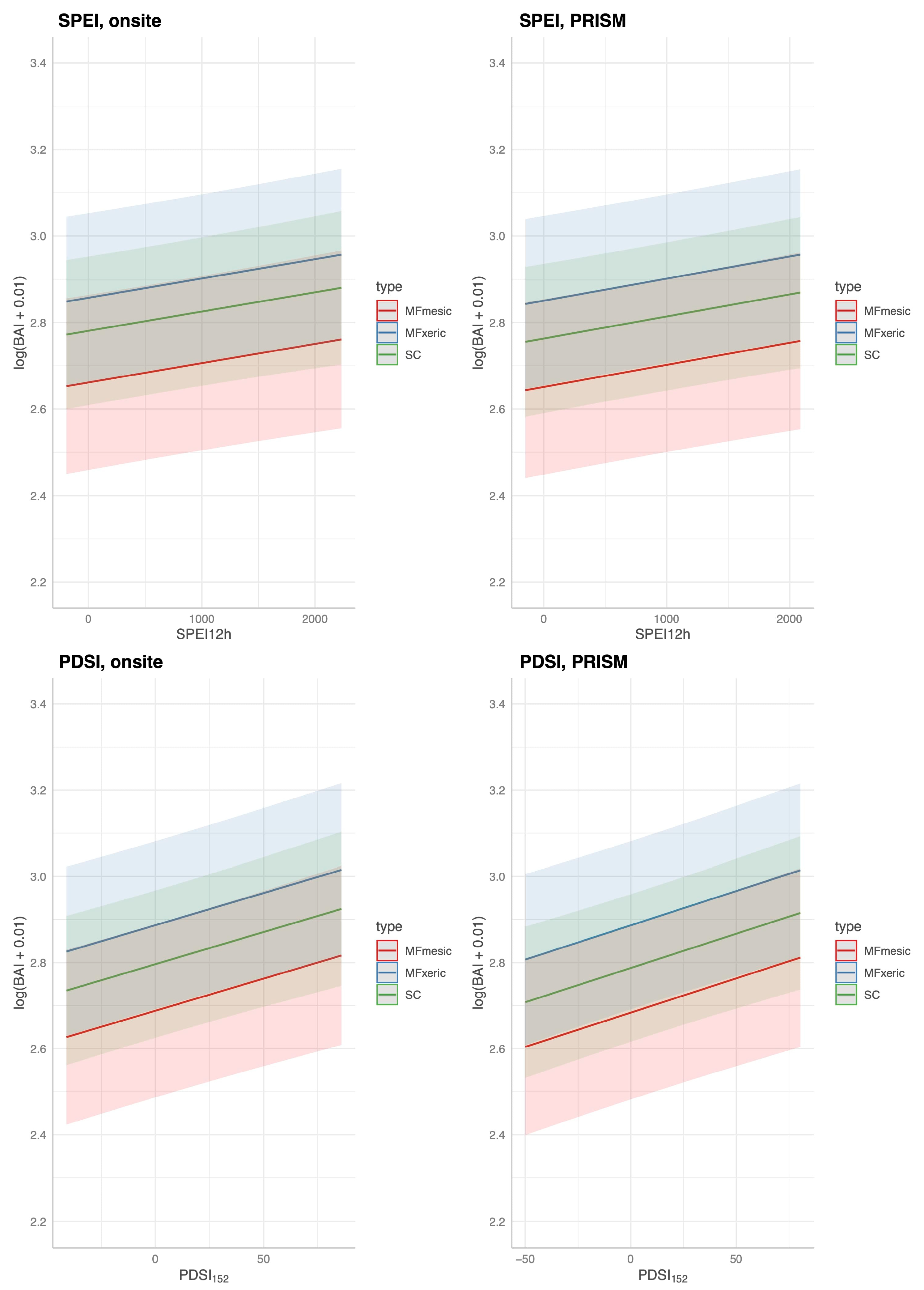
3.6. Drought Index Performance Predicting Site BAI
3.6.1. Predicting BAI at HM
3.6.2. Predicting BAI at MF and SC:
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. On-Site and PRISM Precipitation and Temperature for the HM Site

Appendix B. On-Site and PRISM Precipitation and Temperature for the SC Site

Appendix C. Comparison of On-Site and PRISM Parameterization of PET and the Five Drought Indices at HM

Appendix D. Comparison of On-Site and PRISM Parameterization of PET and the Five Drought Indices at SC

References
- Szejner, P.; Belmecheri, S.; Babst, F.; Wright, W.E.; Frank, D.C.; Hu, J.; Monson, R.K. Stable isotopes of tree rings reveal seasonal-to-decadal patterns during the emergence of a megadrought in the Southwestern US. Oecologia 2021, 197, 1079–1094. [Google Scholar] [CrossRef] [PubMed]
- Ficklin, D.L.; Maxwell, J.T.; Letsinger, S.L.; Gholizadeh, H. A climatic deconstruction of recent drought trends in the United States. Environ. Res. Lett. 2015, 10, 44009. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Swain, D.L.; Touma, D. Anthropogenic warming has increased drought risk in California. Proc. Natl. Acad. Sci. USA 2015, 112, 3931–3936. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.; Anchukaitis, K.J. How unusual is the 2012–2014 California drought? Geophys. Res. Lett. 2014, 41, 9017–9023. [Google Scholar] [CrossRef]
- Allen, C.R.; Fontaine, J.J.; Pope, K.L.; Garmestani, A.S. Adaptive management for a turbulent future. J. Environ. Manag. 2011, 92, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Aerial Detection Survey. 2019. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fseprd700811.pdf (accessed on 31 March 2022).
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Adams, H.D.; Guardiola-Claramonte, M.; Barron-Gafford, G.A.; Villegas, J.C.; Breshears, D.D.; Zou, C.B.; Troch, P.A.; Huxman, T.E. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. USA 2009, 106, 7063–7066. [Google Scholar] [CrossRef]
- McDowell, N.G.; Williams, A.P.; Xu, C.; Pockman, W.T.; Dickman, L.T.; Sevanto, S.; Pangle, R.E.; Limousin, J.M.; Plaut, J.J.; Mackay, D.S.; et al. Multi-scale predictions of massive conifer mortality due to chronic temperature rise. Nat. Clim. Change 2016, 6, 295–300. [Google Scholar] [CrossRef]
- Timmer, K.; Suarez-Brand, M.; Cohen, J.; Clayburgh, J. State of Sierra Waters: A Sierra Nevada Watersheds Index; Sierra Nevada Alliance: Lake Tahoe: Tahoe City, CA, USA, 2006; Available online: https://sierranevadaalliance.org/wpcontent/uploads/sos.pdf (accessed on 15 May 2022).
- Crausbay, S.; Ramirez, A.R.; Carter, S.L.; Cross, M.S.; Hall, K.R.; Bathke, D.J.; Betancourt, J.L.; Colt, S.; Cravens, A.E.; Dalton, M.S.; et al. Defining ecological drought for the twenty-first century. Bull. Am. Meteor. Soc. 2017, 98, 2543–2550. [Google Scholar] [CrossRef]
- Daly, C.; Neilson, R.P.; Phillips, D.L. A statistical-topographic model for mapping climatological precipitation over mountainous terrain. J. Appl. Meteor. Clim. 1994, 33, 140–158. [Google Scholar] [CrossRef]
- PRISM Climate Data. Parameter-elevation Regressions on Independent Slopes Model (PRISM). 2021. Available online: https://prism.oregonstate.edu (accessed on 28 January 2022).
- Daly, C.; Slater, M.E.; Roberti, J.A.; Laseter, S.H.; Swift, L.W., Jr. High-resolution precipitation mapping in a mountainous watershed: Ground truth for evaluating uncertainty in a national precipitation dataset. Int. J. Clim. 2017, 37 (Suppl. S1), 124–137. [Google Scholar] [CrossRef]
- Strachan, S.; Daly, C. Testing the daily PRISM air temperature model on semiarid mountain slopes. J. Geophys. Res. Atmos. 2017, 122, 5697–5715. [Google Scholar] [CrossRef]
- Daly, C.; Conklin, D.R.; Unsworth, M.H. Local atmospheric decoupling in complex topography alters climate change impacts. Int. J. Clim. 2010, 30, 1857–1864. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multi-scalar drought index sensitive to global warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Baltas, E. Spatial distribution of climatic indices in northern Greece. Meteor. Appl. 2007, 14, 69–78. [Google Scholar] [CrossRef]
- Budyko, M.E. Climate and Life; Academic Press: New York, NY, USA, 1974; 508p, ISBN 9780080954530. [Google Scholar]
- Palmer, W.C. Meteorological Drought; Research Paper No. 45; U.S. Department of Commerce, Weather Bureau: Washington, DC, USA, 1965; 58p. Available online: http://www.ncdc.noaa.gov/temp-and-precip/drought/docs/palmer.pdf (accessed on 15 May 2022).
- Wells, N.; Goddard, S.; Hayes, M.J. A Self-Calibrating Palmer Drought Severity Index. J. Clim. 2004, 17, 2335–2351. [Google Scholar] [CrossRef]
- Hubbert, K.R.; Beyers, J.L.; Graham, R.C. Roles of weathered bedrock and soil in seasonal water relations of Pinus Jeffreyi and Arctostaphylos patula. Can. J. For. Res. 2001, 31, 1947–1957. [Google Scholar] [CrossRef]
- Rose, K.; Graham, R.C.; Parker, D. Water source utilization by Pinus jeffreyi and Arctostaphylos patula on thin soils over bedrock. Oecologia 2003, 134, 46–54. [Google Scholar] [CrossRef]
- Grulke, N.E. Physiological responses of ponderosa pine to gradients of environmental stressors. In Oxidant Air Pollution Impacts in the Montane Forests of Southern California: A Case Study of the San Bernardino Mountains; Ecological Studies; Miller, P.R., McBride, J.R., Eds.; Springer: New York, NY, USA, 1999; Volume 134, pp. 126–163. [Google Scholar] [CrossRef]
- Beyaztas, U.; Yaseen, Z.M. Drought interval simulation using functional data analysis. J. Hydrol. 2019, 579, 124141. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Barbero, R.; Wolf, J.W.; Holden, Z.A. Tracking interannual streamflow variability with drought indices in the U.S. Pacific Northwest. J. Hydrometeorol. 2014, 15, 1900–1912. [Google Scholar] [CrossRef]
- Quemada, C.; Pérez-Escudero, J.M.; Gonzalo, R.; Ederra, I.; Santesteban, L.G.; Torres, N.; Iriarte, J.C. Remote sensing for plant water content monitoring: A review. Remote Sens. 2021, 13, 2088. [Google Scholar] [CrossRef]
- Berner, L.T.; Law, B.E.; Hudiburg, T.W. Water availability limits tree productivity, carbon stocks, and carbon residence time in mature forests across the western US. Biogeosciences 2017, 14, 365–378. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, T.; Zhao, X.; Huang, K.; Gao, S.; Wu, H.; Luo, H. Assessments of drought impacts on vegetation in China with the optimal time scales of the Climatic Drought Index. Int. J. Environ. Res. Public Health 2015, 12, 7615–7634. [Google Scholar] [CrossRef]
- Vincente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Begueria, S.; Sanchez-Lorenzo, A. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2021, 110, 52–57. [Google Scholar] [CrossRef]
- Drew, D.M.; Allen, K.; Downes, G.M.; Evans, R.; Battaglia, M.; Baker, P. Wood properties in a long-lived conifer reveal strong climate signals where ring-width series do not. Tree Physiol. 2012, 33, 37–47. [Google Scholar] [CrossRef]
- Pompa-García, M.; Camarero, J.J.; Colangelo, M.; González-Cásares, M. Inter- and intra-annual links between climate, tree growth and NDVI: Improving the resolution of drought proxies in conifer forests. Int. J. Biometeorol. 2021, 65, 2111–2121. [Google Scholar] [CrossRef]
- Pasho, E.; Alla, A.Q. Climate impacts on radial growth and vegetation activity of two co-existing Mediterranean pine species. Can. J. For. Res. 2015, 45, 1748–1756. [Google Scholar] [CrossRef]
- Manion, P.D. Tree Disease Concept, 2nd ed.; Prentice Hall: Hoboken, NJ, USA, 1981; 399p, ISBN 978013930701037. [Google Scholar]
- Su, L.; Cao, Q.; Xiao, M.; Mocko, D.M.; Barlage, M.; Li, D.; Peters-Lidard, C.D.; Lettenmaier, D.P. Drought variability over the conterminous United States for the past century. J. Hydrometeorol. 2021, 22, 1153–1168. [Google Scholar] [CrossRef]
- Preisler, H.K.; Grulke, N.E.; Heath, Z.; Smith, S.L. Analysis and out-year forecast of beetle, borer, and drought-induced tree mortality in California. For. Ecol. Manag. 2017, 399, 166–178. [Google Scholar] [CrossRef]
- Grulke, N.E.; Bienz, C.; Hrinkevich, K.; Maxfield, J.; Uyeda, K. Quantitative and qualitative approaches to assess effectiveness of forest treatments in improving stand health in dry pine forests. For. Ecol. Manag. 2020, 465, 118085. [Google Scholar] [CrossRef]
- Looney, C.E.; D’Amato, A.W.; Jovan, S. Investigating linkages between the size-growth relationship and drought, nitrogen deposition, and structural complexity in western U.S. Forests. For. Ecol. Manag. 2021, 497, 119494. [Google Scholar] [CrossRef]
- Shaw, R. Tree Vigor Response and Competitive Zone Density in Mature Ponderosa Pine. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2017; 85p. Available online: https://ir.library.oregonstate.edu/concern/graduate_thesis_or_dissertations/mc87ps84d (accessed on 25 February 2021).
- Barbour, M.G.; Minnich, R.A. Californian upland forests and woodlands. In North American Terrestrial Vegetation; Barbour, M.G., Billings, W.D., Eds.; Cambridge University Press: Cambridge, UK, 1999; pp. 161–202. ISBN 052155027. Available online: http://catdir.loc.gov/catdir/samples/cam031/97029061.pdf (accessed on 15 May 2022).
- Jones, D.P.; Graham, R.C. Water-holding characteristics of weathered granitic rock in chaparral and forest ecosystems. Soil Sci. Soc. Am. J. 1993, 57, 256–261. [Google Scholar] [CrossRef]
- McCormick, E.L.; Dralle, D.N.; Hahm, W.J.; Tune, A.K.; Schmidt, L.M.; Chadwick, K.D.; Rempe, D.M. Widespread woody plant use of water stored in bedrock. Nature 2021, 597, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Grulke, N.E.; Johnson, R.; Esperanza, A.; Jones, D.; Nguyen, T.; Posch, S.; Tausz, M. Canopy transpiration of Jeffrey pine in mesic and xeric microsites: O3 uptake and injury response. Trees 2003, 17, 292–298. [Google Scholar] [CrossRef]
- Grulke, N.E.; Maxfield, J.; Riggan, P.J.; Schrader-Patton, C.S. Pre-emptive detection of mature pine drought stress using multispectral aerial imagery. Remote Sens. 2020, 12, 2338. [Google Scholar] [CrossRef]
- National Oceanic and Atmosphere Administration (NOAA); National Centers for Environmental Information (NCEI). Climate Data Records. Available online: https://www.ncei.noaa.gov/products/climate-data-records (accessed on 31 March 2022).
- Western Regional Climate Center (WRCC). Southwest Climate and Environmental Information Collaborative (SCENIC), Reno, NV, USA, 1933–2020. Available online: https://wrcc.dri.edu/csc/scenic/ (accessed on 22 February 2022).
- National Atmospheric Deposition Program. (NRSP-3) (NADP), NTN SITE: CA75. Available online: https://nadp.slh.wisc.edu/networks/national-trends-network/ (accessed on 25 February 2022).
- Kim, Y.; Grulke, N.E. 80-Yr Meteorological Records and Drought Indices for Sequoia National Park and Sequoia National Monument, CA; Forest Service Research Data Archive, RDS-2022-0029; U.S. Department of Agriculture: Fort Collins, CO, USA, 2022. [Google Scholar] [CrossRef]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Stannard, D.I. Comparison of Penman-Monteith, Shuttleworth-Wallace, and Modified Priestley-Taylor Evapotranspiration Models for wildland vegetation in semiarid rangeland. Water Resour. Res. 1993, 29, 1379–1392. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper 56; Food and Agriculture Organization, United Nations: Rome, Italy, 1998; 15p, Available online: https://appgeodb.nancy.inra.fr/biljou/pdf/Allen_FAO1998.pdf (accessed on 15 May 2022).
- Karl, T.R. The sensitivity of the Palmer Drought Severity Index and Palmer’s Z-Index to their calibration coefficients including potential evapotranspiration. J. Appl. Meteor. Clim. 1986, 25, 77–86. [Google Scholar] [CrossRef]
- Zhong, R.; Chen, X.; Wang, Z.; Lai, C. scPDSI: Calculation of the Conventional and Self-Calibrating Palmer Drought Severity Index. 2018. Available online: https://rdrr.io/cran/scPDSI/ (accessed on 18 May 2022).
- United States Geological Survey (USGS); National Water Information System (NWIS). USGS Water Data for the Nation Help. Available online: https://help.waterdata.usgs.gov (accessed on 25 February 2022).
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Dougherty, P.M.; Whitehead, D.; Vose, J.M. Environmental Influences on the phenology of pine. Ecol. Bull. 1994, 43, 64–75. Available online: https://www.jstor.org/stable/20113132?seq=1#metadata_info_tab_contents (accessed on 15 May 2022).
- Sloan, J.P. Ponderosa and Lodgepole Pine Seedling Bud Burst Varies with Lift Date and Cultural Practices in Idaho Nursery; RN-INT-397; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1991; 9p, Available online: https://www.worldcat.org/title/ponderosa-and-lodgepole-pine-seedling-bud-burst-varies-with-lift-date-and-cultural-practices-in-idaho-nursery/oclc/679979260&referer=brief_results (accessed on 15 May 2022).
- Andersen, C.P.; Sucoff, E.I.; Dixon, R.K. Effects of root zone temperature on root initiation and elongation in red pine seedlings. Can. J. For. Res. 1986, 16, 696–700. [Google Scholar] [CrossRef]
- Levin, D.A.; Grulke, N.E.; Bienz, C.; Hrinkevich, K.; Merschel, A.; Uyeda, K.A. Forest treatment effects on wood production in ponderosa pine. For. Ecol. Manag. 2022, in press. Available online: https://pages.uoregon.edu/dlevin/PAPERS/sycan.pdf (accessed on 15 May 2022).
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree Ring Dating; University of Arizona Press: Flagstaff, AZ, USA, 1996; 73p, ISBN 9780816516803. [Google Scholar]
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1991, 21, 414–416. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree-Ring Res. 2000, 57, 205–221. Available online: https://www.ltrr.arizona.edu/~ellisqm/outgoing/dendroecology2014/readings/Grissino_mayer_COFECHA_2001.pdf (accessed on 15 May 2022).
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. Available online: https://www.ltrr.arizona.edu/~ellisqm/outgoing/dendroecology2014/readings/Holmes_1983.pdf (accessed on 15 May 2022).
- Applequist, M.B. A simple pith locator for use with off-center increment cores. J. For. 1958, 56, 141. Available online: https://academic.oup.com/jof/article/56/2/138/4685764?login=true92 (accessed on 15 May 2022).
- Grulke, N.E.; Johnson, R.; Monschein, S.; Nikolova, P.; Tausz, M. Variation in morphological and biochemical O3 injury attributes of mature Jeffrey pine within canopies and between microsites. Tree Physiol. 2003, 23, 923–929. [Google Scholar] [CrossRef][Green Version]
- University of Minnesota. United States Monthly Climate Summary. December 1992–January 1993. Available online: https://www.google.com/books/edition/Weekly_Climate_Bulletin/YRYs5pcurcoC?hl=en&gbpv=1&dq=%22united+states+monthly+climate+summary+December+1992%22&pg=PA11&printsec=frontcover (accessed on 22 February 2022).
- Athanasiadis, P.J.; Wallace, J.M.; Wettstein, J.J. Patterns of wintertime jet Stream variability and their relation to the storm tracks. J. Atmos. Sci. 2010, 67, 1361–1381. [Google Scholar] [CrossRef]
- Deo, R.C.; Şahin, M. Application of the extreme learning machine algorithm for the prediction of monthly Effective Drought Index in eastern Australia. Atmos. Res. 2015, 153, 512–525. [Google Scholar] [CrossRef]
- Bales, R.C.; Goulden, M.L.; Hunsaker, C.T.; Conklin, M.H.; Hartsough, P.C.; O’Geen, A.T.; Hopmans, J.; Safeeq, M. Mechanisms controlling the impact of multi-year drought on mountain hydrology. Sci. Rep. 2018, 8, 690. [Google Scholar] [CrossRef] [PubMed]
- Pechlivanidis, I.G.; Crochemore, L.; Rosberg, J.; Bosshard, T. What are the key drivers controlling the quality of seasonal streamflow forecasts? Water Resour. Res. 2020, 56, e2019WR026987. [Google Scholar] [CrossRef]
- Yuhas, A.N.; Scuderi, L.A. MODIS-derived NDVI characterisation of drought-induced evergreen die-off in western North America. Geogr. Res. 2009, 47, 34–45. [Google Scholar] [CrossRef]
- Easdale, M.H.; Bruzzone, O.; Mapfumo, P.; Tittonell, P. Phases or regimes? Revisiting NDVI trends as proxies for land degradation. Land Degrad. Dev. 2018, 29, 433–445. [Google Scholar] [CrossRef]
- Hargrove, W.W.; Spruce, J.P.; Gasser, G.E.; Hoffman, F.M. Toward a national early warning system for forest disturbances using remotely sensed canopy phenology. Photogramm. Eng. Remote Sens. 2009, 75, 1150–1156. Available online: https://www.fs.usda.gov/treesearch/pubs/33669 (accessed on 15 May 2022).
- Wong, C.Y.S.; Young, D.J.N.; Latimer, A.M.; Buckley, T.N.; Magney, T.S. Importance of the legacy effect for assessing spatiotemporal correspondence between interannual tree-ring width and remote sensing products in the Sierra Nevada. Remote Sens. Environ. 2021, 265, 112635. [Google Scholar] [CrossRef]
- Schrader-Patton, C.; Grulke, N.E.; Bienz, C. Landscape assessment of ponderosa pine vigor using 4-Band aerial imagery in south central Oregon. Forests 2021, 12, 612. [Google Scholar] [CrossRef]
- Lu, J.; Carbone, G.J.; Gao, P. Mapping the agricultural drought based on the long-term AVHRR NDVI and North American Regional Re-analysis (NARR) in the United States, 1981–2013. Appl. Geogr. 2019, 104, 10–20. [Google Scholar] [CrossRef]
- Uyeda, K.A.; Stow, D.A.; Roberts, D.A.; Riggan, P.J. Combining ground-based measurements and MODIS-based spectral vegetation indices to track biomass accumulation in post-fire chaparral. Int. J. Remote Sens. 2017, 38, 718–741. [Google Scholar] [CrossRef]
- Liu, Y.; Kumar, M.; Katul, G.G.; Porporato, A. Reduced resilience as an early warning signal of forest mortality. Nature Clim. Change 2019, 9, 880–885. Available online: https://www.nature.com/articles/s41558-019-0583-9 (accessed on 15 May 2022). [CrossRef]
- Peng, J.; Dadson, S.; Hirpa, F.; Dyer, E.; Lees, T.; Miralles, D.G.; Vicente-Serrano, S.M.; Funk, C. A pan-African high-resolution drought index dataset. Earth Syst. Sci. Data 2020, 12, 753–769. [Google Scholar] [CrossRef]
- Paz-Kagan, T.; Vaughn, N.R.; Martin, R.E.; Brodrick, P.G.; Stephenson, N.L.; Das, A.J.; Nydick, K.R.; Asner, G.P. Landscape-scale variation in canopy water content of giant sequoias during drought. For. Ecol. Manag. 2018, 419–420, 291–304. [Google Scholar] [CrossRef]
- Baeza, A.; Martin, R.E.; Stephenson, N.L.; Das, A.J.; Hardwick, P.; Nydick, K.; Mallory, J.; Slaton, M.; Evans, K.; Asner, G.P. Mapping the vulnerability of giant sequoias after extreme drought in California using remote sensing. Ecol. Appl. 2021, 31, e02395. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.E.; Ehleringer, J.R. Streamside trees that do not use Stream water. Nature 1991, 350, 335–337. [Google Scholar] [CrossRef]
- Miles, S.R.; Goudey, C.B. (Comps.) Ecological Sub-Regions of California: Section and Subsection Descriptions; R5-EM-TP-005; USDA Forest Service, Technical Publication, Pacific Southwest Region: Vallejo, CA, USA, 1997; 200p. [Google Scholar]
- Martin-Benito, D.; Beeckman, H.; Cañellas, I. Influence of drought on tree rings and tracheid features of Pinus nigra and Pinus sylvestris in a mesic Mediterranean forest. Eur. J. For. Res. 2012, 132, 33–45. [Google Scholar] [CrossRef]
- Bhuyan, U.; Zang, C.; Menzel, A. Different responses of multispecies tree ring growth to various DIs across Europe. Dendrochronologia 2017, 44, 1–8. [Google Scholar] [CrossRef]
- Staszak, J.; Grulke, N.E.; Marrett, M.J.; Prus-Glowacki, W. Isozyme markers associated with O3 tolerance indicate shift in genetic structure of ponderosa and Jeffrey pine in Sequoia National Park, California. Environ. Pollut. 2007, 149, 366–375. [Google Scholar] [CrossRef]
- Meyer, M.D.; North, M.P.; Gray, A.N.; Zald, H.S.J. Influence of soil thickness on stand characteristics in a Sierra Nevada mixed-conifer forest. Plant Soil 2007, 294, 113–123. [Google Scholar] [CrossRef]
- Potter, K.A.; Woods, H.A.; Pincebourde, S. Microclimate challenges in global change biology. Glob. Change Biol. 2013, 10, 2932–2939. [Google Scholar] [CrossRef]
- Robeson, S.M. Revisiting the recent California drought as an extreme value. Geophys. Res. Lett. 2015, 42, 6771–6779. [Google Scholar] [CrossRef]
- Su, Y.; Bales, R.C.; Ma, Q.; Nydick, K.; Ray, R.L.; Li, W.; Guo, Q. Emerging stress and relative resiliency of giant sequoia groves experiencing multiyear dry periods in a warming climate. J. Geophys. Res. Biogeosci. 2017, 122, 3063–3075. [Google Scholar] [CrossRef]
- Buras, A.; Schunk, C.; Zeiträg, C.; Herrmann, C.; Kaiser, L.; Lemme, H.; Straub, C.; Taeger, S.; Gößwein, S.; Klemmt, H.-J.; et al. Are Scots pine forest edges particularly prone to drought-induced mortality? Environ. Res. Lett. 2018, 13, 25001. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.A. Forest Stand Dynamics. Updated Edition. In Yale School of the Environment 1; John Wiley & Sons: Hoboken, NJ, USA, 1996; Available online: https://elischolar.library.yale.edu/fes_pubs/1/ (accessed on 15 May 2022).
- Biging, G.S.; Dobbertin, M. Evaluation of competition indices in individual tree growth models. For. Sci. 1995, 41, 360–377. [Google Scholar]
- Grulke, N.E.; Paine, T.; Minnich, R.; Chavez, D.; Riggan, P.; Dunn, A. Air pollution increases forest susceptibility to wildfire. In Wildland Fires and Air Pollution; Developments in Environmental Science; Bytnerowicz, A., Arbaugh, M., Riebau, A., Andersen, C., Eds.; Elsevier Publishers: The Hague, The Netherlands, 2009; Volume 8, pp. 365–403. [Google Scholar] [CrossRef]
- Asner, G.P. Progressive forest canopy water loss during the 2012–2015 California drought. Proc. Natl. Acad. Sci. USA 2016, 113, E249–E255. [Google Scholar] [CrossRef]
- Wong, C.Y.S.; Gamon, J.A. Three causes of variation in the photochemical reflectance index (PRI) in evergreen conifers. New Phytol. 2015, 206, 187–195. Available online: https://www.jstor.org/stable/newphytologist.206.1.187 (accessed on 15 May 2022). [CrossRef]
- Bell, D.M.; Cohen, W.B.; Reilly, M.; Yang, Z. Visual interpretation and time series modeling of Landsat imagery highlight drought’s role in forest canopy declines. Ecosphere 2018, 9, e02195. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Berry, J.A.; Smith, D.D.; Sperry, J.S.; Anderegg, L.D.L.; Field, C.B. The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proc. Natl. Acad. Sci. USA 2012, 109, 233–237. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| INDEX | MF | HM | SC |
|---|---|---|---|
| PET12h | 0.99 | 0.99 | 1.00 |
| SPEI12h | 0.98 | 0.98 | 0.97 |
| AI12h | 0.97 | 0.97 | 0.97 |
| PDSI12h152 | 0.96 | 0.95 | 0.95 |
| PDSI12Hh100 | 0.95 | 0.94 | 0.94 |
| scPDSI12h152 | 0.91 | 0.88 | 0.88 |
| scPDSI12h100 | 0.87 | 0.90 | 0.92 |
| PHDI12h152 | 0.95 | 0.95 | 0.91 |
| PHDI12h100 | 0.95 | 0.94 | 0.90 |
| SPEI12h | AI12h | PDSI12h152 | PDSI12h100 | scPDSI12h152 | scPDSI12h100 | PHDI12h152 | |
|---|---|---|---|---|---|---|---|
| AI12h | 0.98 (0.98) | ||||||
| PDSI12h152 | 0.81 (0.85) | 0.81 (0.85) | |||||
| PDSI12h100 | 0.81 (0.85) | 0.81 (0.86) | 0.99 (0.99) | ||||
| scPDFI12h152 | 0.71 (0.82) | 0.70 (0.82) | 0.90 (0.98) | 0.88 (0.97) | |||
| scPDFI12h100 | 0.72 (0.81) | 0.72 (0.80) | 0.91 (0.95) | 0.91 (0.95) | 0.93 (0.97) | ||
| PHDI12h152 | 0.75 (0.77) | 0.75 (0.77) | 0.90 (0.91) | 0.88 (0.90) | 0.89 (0.91) | 0.88 (0.88) | |
| PHDI12h100 | 0.75 (0.78) | 0.75 (0.78) | 0.89 (0.91) | 0.89 (0.91) | 0.88 (0.90) | 0.88 (0.89) | 0.99 (0.99) |
| SPEI12h | AI12h | PDSI12h152 | PDSI12h100 | scPDSI12h152 | scPDSI12h100 | PHDI12h152 | PHDI12h100 | |
|---|---|---|---|---|---|---|---|---|
| On-site | 1.00 | 0.98 | 0.80 | 0.74 | 0.78 | 0.72 | 0.82 | 0.81 |
| PRISM | 0.99 | 0.98 | 0.88 | 0.88 | 0.87 | 0.74 | 0.85 | 0.84 |
| Estimate | S. E. | t Value | Pr(>|t|) | Estimate | S. E. | t Value | Pr(>|t|) | ||
|---|---|---|---|---|---|---|---|---|---|
| SPEI/ON-SITE | PDSI152/ON-SITE | ||||||||
| (Intercept) | 0.5268 | 0.0031 | 168.83 | 0.00 | (Intercept) | 0.5296 | 0.0033 | 160.66 | 0.00 |
| site_typeMFxeric | −0.0061 | 0.0043 | −1.41 | 0.16 | site_typeMFxeric | −0.0061 | 0.0046 | −1.30 | 0.20 |
| site_typeHM | −0.0002 | 0.0045 | −0.04 | 0.96 | site_typeHM | −0.0060 | 0.0047 | −1.28 | 0.20 |
| site_typeSC | 0.0084 | 0.0046 | 1.82 | 0.07 | site_typeSC | 0.0029 | 0.0047 | 0.62 | 0.53 |
| scale(SPEI12h_nolag) | 0.0047 | 0.0017 | 2.81 | 0.01 | scale(SPEI12h_nolag) | 0.0058 | 0.0019 | 3.00 | 0.00 |
| scale(SPEI12h_lag1) | 0.0064 | 0.0016 | 4.03 | 0.00 | scale(SPEI12h_lag1) | 0.0022 | 0.0018 | 1.24 | 0.22 |
| scale(SPEI12h_lag2) | 0.0028 | 0.0016 | 1.81 | 0.08 | scale(SPEI12h_lag2) | 0.0011 | 0.0018 | 0.06 | 0.55 |
| scale(SPEI12h_lag3) | 0.0042 | 0.0017 | 2.46 | 0.02 | scale(SPEI12h_lag3) | 0.0037 | 0.0020 | 1.84 | 0.07 |
| scale(YEAR) | 0.0034 | 0.0016 | 2.19 | 0.03 | scale(YEAR) | 0.0044 | 0.0018 | 2.43 | 0.02 |
| SPEI/PRISM | PDSI152/PRISM | ||||||||
| (Intercept) | 0.5282 | 0.0031 | 169.33 | 0.00 | (Intercept) | 0.5300 | 0.0032 | 163.34 | 0.00 |
| site_typeMFxeric | −0.0061 | 0.0044 | −1.39 | 0.17 | site_typeMFxeric | −0.0061 | 0.0046 | −1.32 | 0.19 |
| site_typeHM | −0.0021 | 0.0045 | −0.46 | 0.64 | site_typeHM | −0.0068 | 0.0046 | −1.48 | 0.14 |
| site_typeSC | 0.0045 | 0.0044 | 1.01 | 0.32 | site_typeSC | 0.0021 | 0.0046 | 0.45 | 0.65 |
| scale(SPEI12h_nolag) | 0.0047 | 0.0017 | 2.78 | 0.01 | scale(SPEI12h_nolag) | 0.0059 | 0.0019 | 3.06 | 0.00 |
| scale(SPEI12h_lag1) | 0.0061 | 0.0016 | 3.85 | 0.00 | scale(SPEI12h_lag1) | 0.0027 | 0.0018 | 1.55 | 0.13 |
| scale(SPEI12h_lag2) | 0.0014 | 0.0016 | 0.87 | 0.39 | scale(SPEI12h_lag2) | 0.0000 | 0.0019 | −0.02 | 0.98 |
| scale(SPEI12h_lag3) | 0.0035 | 0.0018 | 1.98 | 0.05 | scale(SPEI12h_lag3) | 0.0036 | 0.0021 | 1.70 | 0.09 |
| scale(YEAR) | 0.0047 | 0.0017 | 2.74 | 0.01 | scale(YEAR) | 0.0058 | 0.0021 | 2.80 | 0.01 |
| AI/ON-SITE | PDSI100/ON-SITE | ||||||||
| (Intercept) | 0.5257 | 0.0032 | 163.13 | 0.00 | (Intercept) | 0.5292 | 0.0033 | 162.55 | 0.00 |
| site_typeMFxeric | −0.0061 | 0.0043 | −1.40 | 0.17 | site_typeMFxeric | −0.0061 | 0.0046 | −1.32 | 0.19 |
| site_typeHM | 0.0024 | 0.0048 | 0.50 | 0.62 | site_typeHM | −0.0054 | 0.0046 | −1.17 | 0.25 |
| site_typeSC | 0.0101 | 0.0048 | 2.10 | 0.04 | site_typeSC | 0.0040 | 0.0047 | 0.85 | 0.40 |
| scale(SPEI12h_nolag) | 0.0050 | 0.0017 | 2.93 | 0.00 | scale(SPEI12h_nolag) | 0.0061 | 0.0019 | 3.24 | 0.00 |
| scale(SPEI12h_lag1) | 0.0067 | 0.0016 | 4.09 | 0.00 | scale(SPEI12h_lag1) | 0.0026 | 0.0017 | 1.55 | 0.13 |
| scale(SPEI12h_lag2) | 0.0022 | 0.0016 | 1.37 | 0.18 | scale(SPEI12h_lag2) | 0.0012 | 0.0018 | 0.69 | 0.49 |
| scale(SPEI12h_lag3) | 0.0034 | 0.0017 | 1.98 | 0.15 | scale(SPEI12h_lag3) | 0.0039 | 0.0019 | 2.05 | 0.04 |
| scale(YEAR) | 0.0036 | 0.0016 | 2.20 | 0.03 | scale(YEAR) | 0.0043 | 0.0018 | 2.44 | 0.02 |
| AI/PRISM | PDSI100/PRISM | ||||||||
| (Intercept) | 0.5276 | 0.0032 | 165.45 | 0.00 | (Intercept) | 0.5301 | 0.0032 | 164.10 | 0.00 |
| site_typeMFxeric | −0.0061 | 0.0044 | −1.38 | 0.17 | site_typeMFxeric | −0.0061 | 0.0046 | −1.33 | 0.19 |
| site_typeHM | −0.0006 | 0.0047 | −0.14 | 0.89 | site_typeHM | −0.0070 | 0.0046 | −1.53 | 0.13 |
| site_typeSC | 0.0053 | 0.0046 | 1.17 | 0.25 | site_typeSC | 0.0017 | 0.0046 | 0.36 | 0.72 |
| scale(SPEI12h_nolag) | 0.0048 | 0.0017 | 2.83 | 0.01 | scale(SPEI12h_nolag) | 0.0058 | 0.0018 | 3.12 | 0.00 |
| scale(SPEI12h_lag1) | 0.0061 | 0.0016 | 3.77 | 0.00 | scale(SPEI12h_lag1) | 0.0030 | 0.0017 | 1.77 | 0.08 |
| scale(SPEI12h_lag2) | 0.0006 | 0.0017 | 0.37 | 0.71 | scale(SPEI12h_lag2) | −0.0005 | 0.0019 | −0.25 | 0.81 |
| scale(SPEI12h_lag3) | 0.0028 | 0.0018 | 1.55 | 0.13 | scale(SPEI12h_lag3) | 0.0032 | 0.0020 | 1.61 | 0.11 |
| scale(YEAR) | 0.0045 | 0.0018 | 2.53 | 0.01 | scale(YEAR) | 0.0053 | 0.0020 | 2.63 | 0.01 |
| SPEI/ON-SITE | AIC | BIC | logLik | Deviance | Chisq | Pr(>Chisq) |
|---|---|---|---|---|---|---|
| submodel | 1953.53 | 1995.49 | −967.77 | 1935.53 | ||
| full model | 1954.58 | 2015.19 | −964.29 | 1928.58 | 6.95 | 0.14 |
| SPEI/PRISM | ||||||
| submodel | 1953.53 | 1995.49 | −967.77 | 1935.53 | ||
| full model | 1954.08 | 2014.68 | −964.04 | 1928.08 | 7.45 | 0.11 |
| PDSI152/ON-SITE | ||||||
| submodel | 1953.53 | 1995.49 | −967.77 | 1935.53 | ||
| full model | 1957.87 | 2018.47 | −964.94 | 1931.87 | 3.66 | 0.45 |
| PDSI152/PRISM | ||||||
| submodel | 1953.53 | 1995.49 | −967.77 | 1935.53 | ||
| full model | 1956.15 | 2016.75 | −964.08 | 1930.15 | 5.38 | 0.25 |
| SPEI/ON-SITE | Value | S.E. | t | p | PDSI152/ON-SITE | Value | S.E. | t | p |
|---|---|---|---|---|---|---|---|---|---|
| (intercept) | 1.05 | 0.24 | 4.36 | 0.00 | (intercept) | 1.05 | 0.24 | 4.37 | 0.00 |
| scale(SPEI12h_nolag) | 0.02 | 0.01 | 3.15 | 0.00 | scale(SPEI12h_nolag) | 0.03 | 0.01 | 4.61 | 0.00 |
| scale(SPEI12h_lag1) | 0.02 | 0.01 | 3.48 | 0.00 | scale(SPEI12h_lag1) | 0.03 | 0.01 | 5.17 | 0.00 |
| scale(SPEI12h)_lag2) | −0.01 | 0.01 | −1.10 | 0.27 | scale(SPEI12h)_lag2) | −0.02 | 0.01 | −2.91 | 0.00 |
| scale(SPEI12h_lag3) | −0.01 | 0.01 | −1.03 | 0.30 | scale(SPEI12h_lag3) | 0.00 | 0.01 | −0.27 | 0.79 |
| typeMFxeric | 0.12 | 0.15 | 0.79 | 0.43 | typeMFxeric | 0.12 | 0.15 | 0.79 | 0.43 |
| typeSC | 0.05 | 0.13 | 0.35 | 0.73 | typeSC | 0.04 | 0.13 | 0.29 | 0.77 |
| DBH_AGE | 3.21 | 0.26 | 12.36 | 0.00 | DBH_AGE | 3.21 | 0.26 | 12.36 | 0.00 |
| RANK | 0.06 | 0.08 | 0.81 | 0.42 | RANK | 0.06 | 0.08 | 0.81 | 0.42 |
| CZD | −0.01 | 0.01 | −2.72 | 0.01 | CZD | −0.01 | 0.01 | −2.72 | 0.01 |
| s(YEAR):typeMFmesicFx1 | 0.18 | 0.10 | 1.80 | 0.07 | s(YEAR):typeMFmesicFx1 | 0.18 | 0.10 | 1.74 | 0.08 |
| s(YEAR):typeMFxericFx1 | 0.12 | 0.10 | 1.21 | 0.22 | s(YEAR):typeMFxericFx1 | 0.11 | 0.11 | 1.08 | 0.28 |
| s(YEAR):typeSCFx1 | 0.54 | 0.18 | 3.02 | 0.00 | s(YEAR):typeSCFx1 | 0.53 | 0.18 | 2.99 | 0.00 |
| SPEI/PRISM | PDSI152/PRISM | ||||||||
| (intercept) | 1.05 | 0.24 | 4.37 | 0.00 | (intercept) | 1.05 | 0.24 | 4.38 | 0.00 |
| scale(SPEI12h_nolag) | 0.03 | 0.01 | 3.64 | 0.00 | scale(SPEI12h_nolag) | 0.03 | 0.01 | 4.66 | 0.00 |
| scale(SPEI12h_lag1) | 0.03 | 0.01 | 3.96 | 0.00 | scale(SPEI12h_lag1) | 0.03 | 0.01 | 4.41 | 0.00 |
| scale(SPEI12h)_lag2) | −0.01 | 0.01 | −1.58 | 0.11 | scale(SPEI12h)_lag2) | −0.02 | 0.01 | −2.80 | 0.01 |
| scale(SPEI12h_lag3) | −0.01 | 0.01 | −1.20 | 0.23 | scale(SPEI12h_lag3) | 0.00 | 0.01 | −0.28 | 0.78 |
| typeMFxeric | 0.12 | 0.15 | 0.79 | 0.43 | typeMFxeric | 0.12 | 0.15 | 0.79 | 0.43 |
| typeSC | 0.04 | 0.13 | 0.31 | 0.76 | typeSC | 0.04 | 0.13 | 0.27 | 0.79 |
| DBH_AGE | 3.21 | 0.26 | 12.36 | 0.00 | DBH_AGE | 3.21 | 0.26 | 12.36 | 0.00 |
| RANK | 0.06 | 0.08 | 0.81 | 0.42 | RANK | 0.06 | 0.08 | 0.81 | 0.42 |
| CZD | −0.01 | 0.01 | −2.72 | 0.01 | CZD | −0.01 | 0.01 | −2.72 | 0.01 |
| s(YEAR):typeMFmesicFx1 | 0.19 | 0.10 | 1.85 | 0.06 | s(YEAR):typeMFmesicFx1 | 0.20 | 0.11 | 1.87 | 0.06 |
| s(YEAR):typeMFxericFx1 | 0.13 | 0.11 | 1.20 | 0.23 | s(YEAR):typeMFxericFx1 | 0.12 | 0.11 | 1.13 | 0.26 |
| s(YEAR):typeSCFx1 | 0.55 | 0.18 | 3.07 | 0.00 | s(YEAR):typeSCFx1 | 0.54 | 0.18 | 3.07 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Grulke, N.E.; Merschel, A.G.; Uyeda, K.A. Assessing Role of Drought Indices in Anticipating Pine Decline in the Sierra Nevada, CA. Climate 2022, 10, 72. https://doi.org/10.3390/cli10050072
Kim Y, Grulke NE, Merschel AG, Uyeda KA. Assessing Role of Drought Indices in Anticipating Pine Decline in the Sierra Nevada, CA. Climate. 2022; 10(5):72. https://doi.org/10.3390/cli10050072
Chicago/Turabian StyleKim, Yoonji, Nancy E. Grulke, Andrew G. Merschel, and Kellie A. Uyeda. 2022. "Assessing Role of Drought Indices in Anticipating Pine Decline in the Sierra Nevada, CA" Climate 10, no. 5: 72. https://doi.org/10.3390/cli10050072
APA StyleKim, Y., Grulke, N. E., Merschel, A. G., & Uyeda, K. A. (2022). Assessing Role of Drought Indices in Anticipating Pine Decline in the Sierra Nevada, CA. Climate, 10(5), 72. https://doi.org/10.3390/cli10050072