Predicting the Potential Current and Future Distribution of the Endangered Endemic Vascular Plant Primula boveana Decne. ex Duby in Egypt

 ,
,  ,
,  ,
,  and
and
Abstract
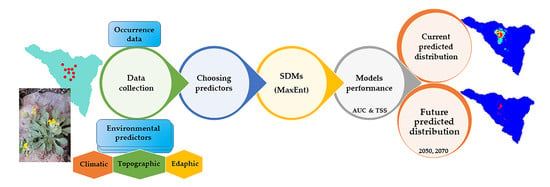
1. Introduction
2. Results
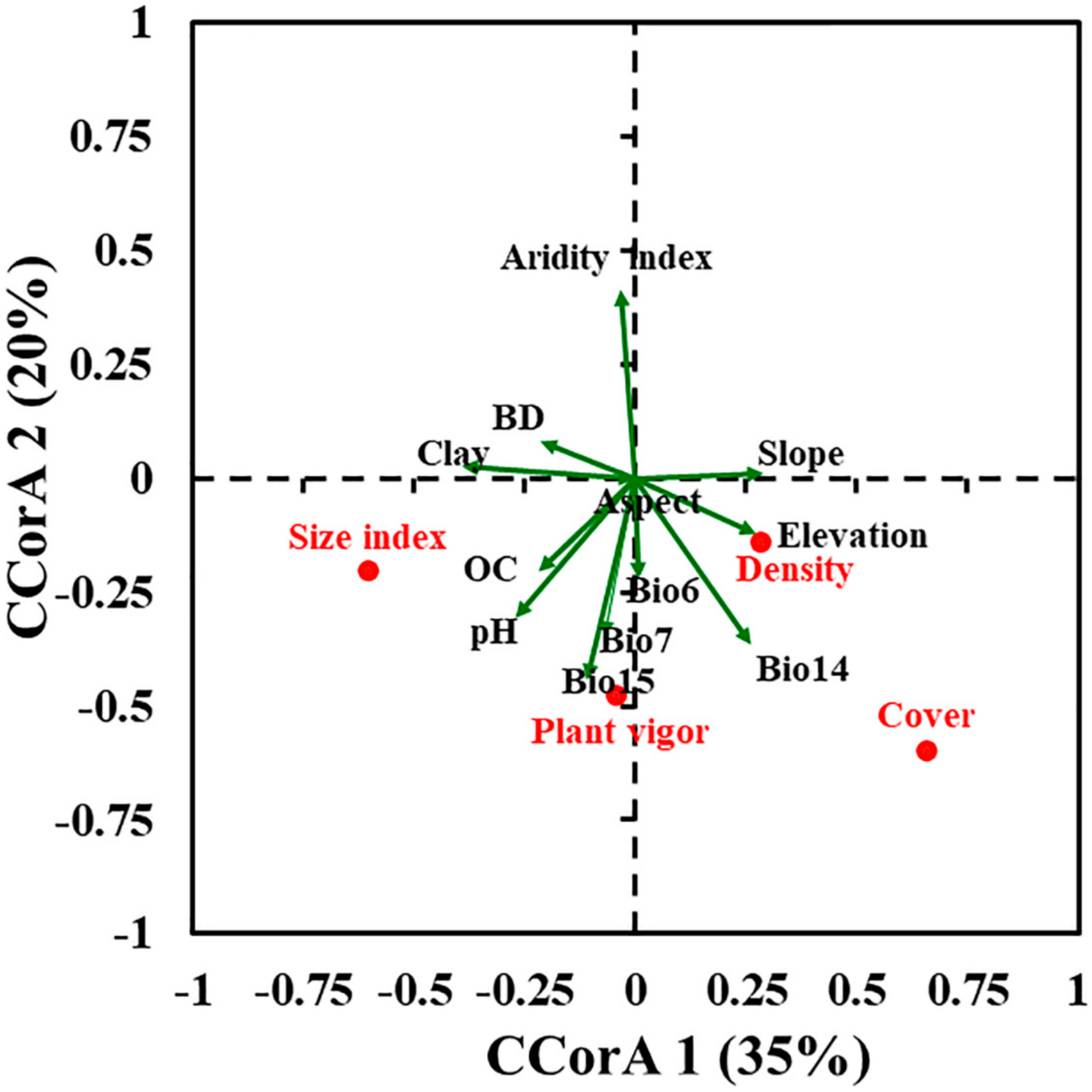
2.1. Population Status, Original Habitat Features, and Influencing Variables
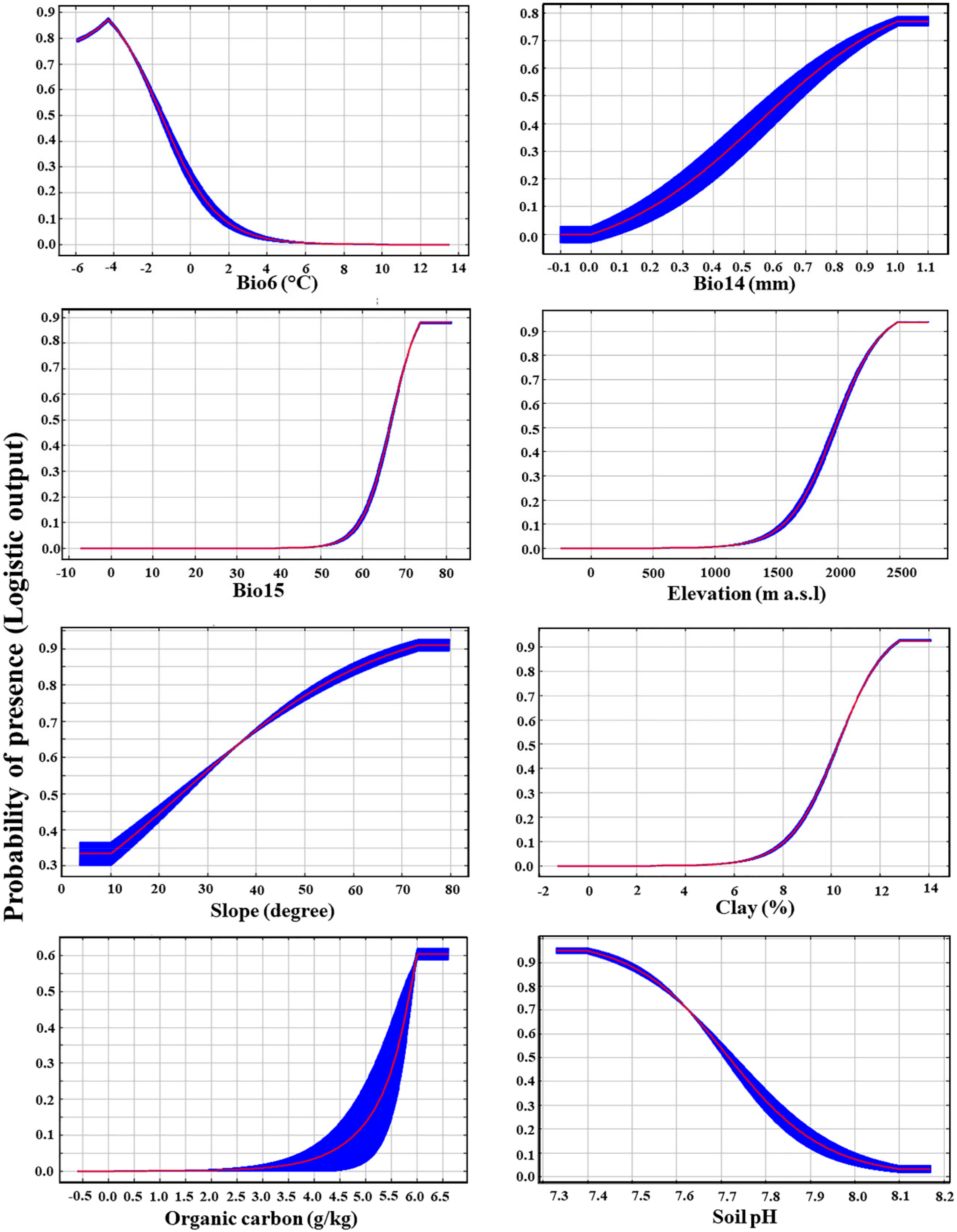
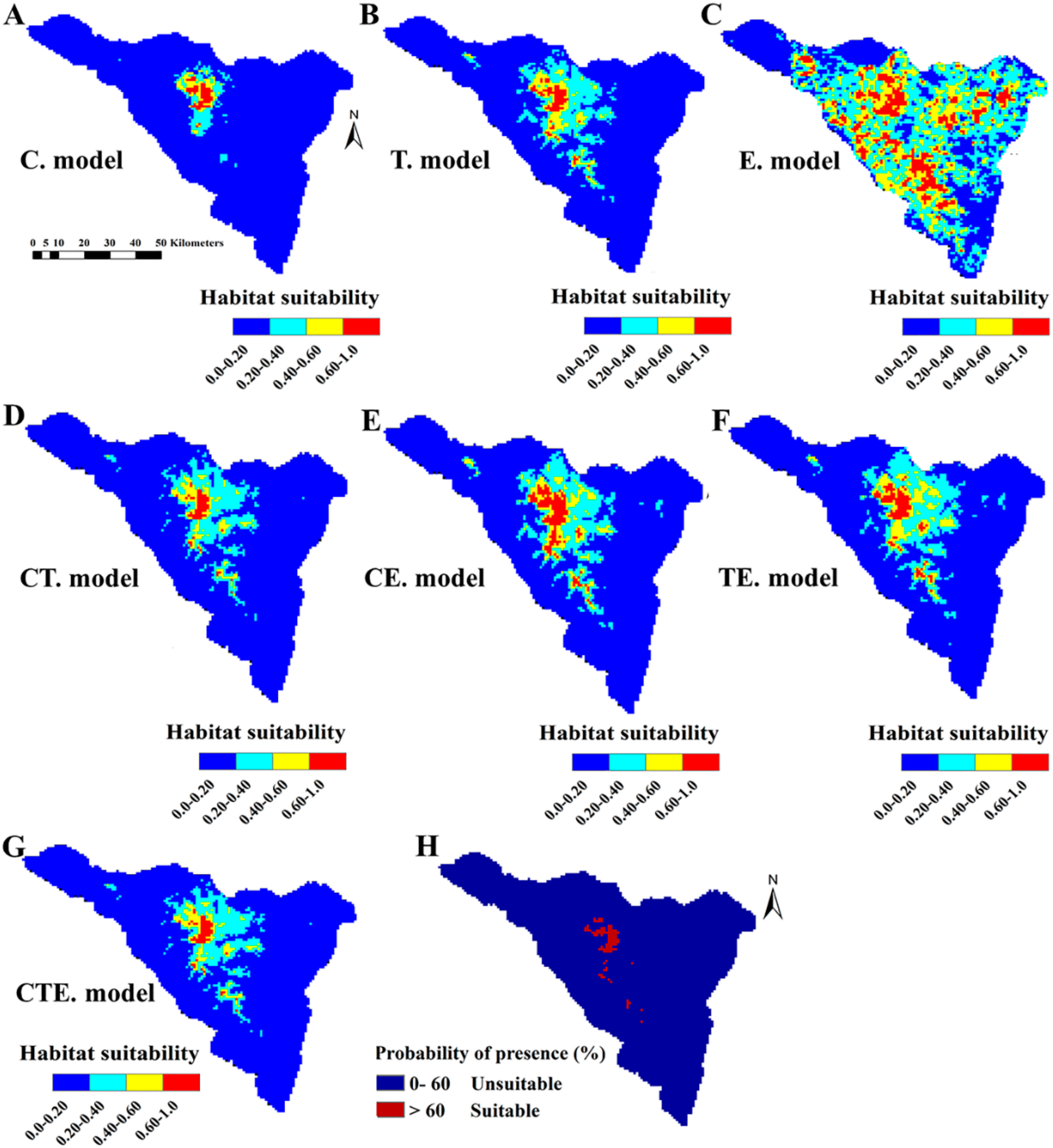
2.2. Models Evaluation and Contributions of Variables
2.3. Predictive Potential Current Habitat Suitability of P. boveana
2.4. Potential Areas for New Population Survey or Reintroduction
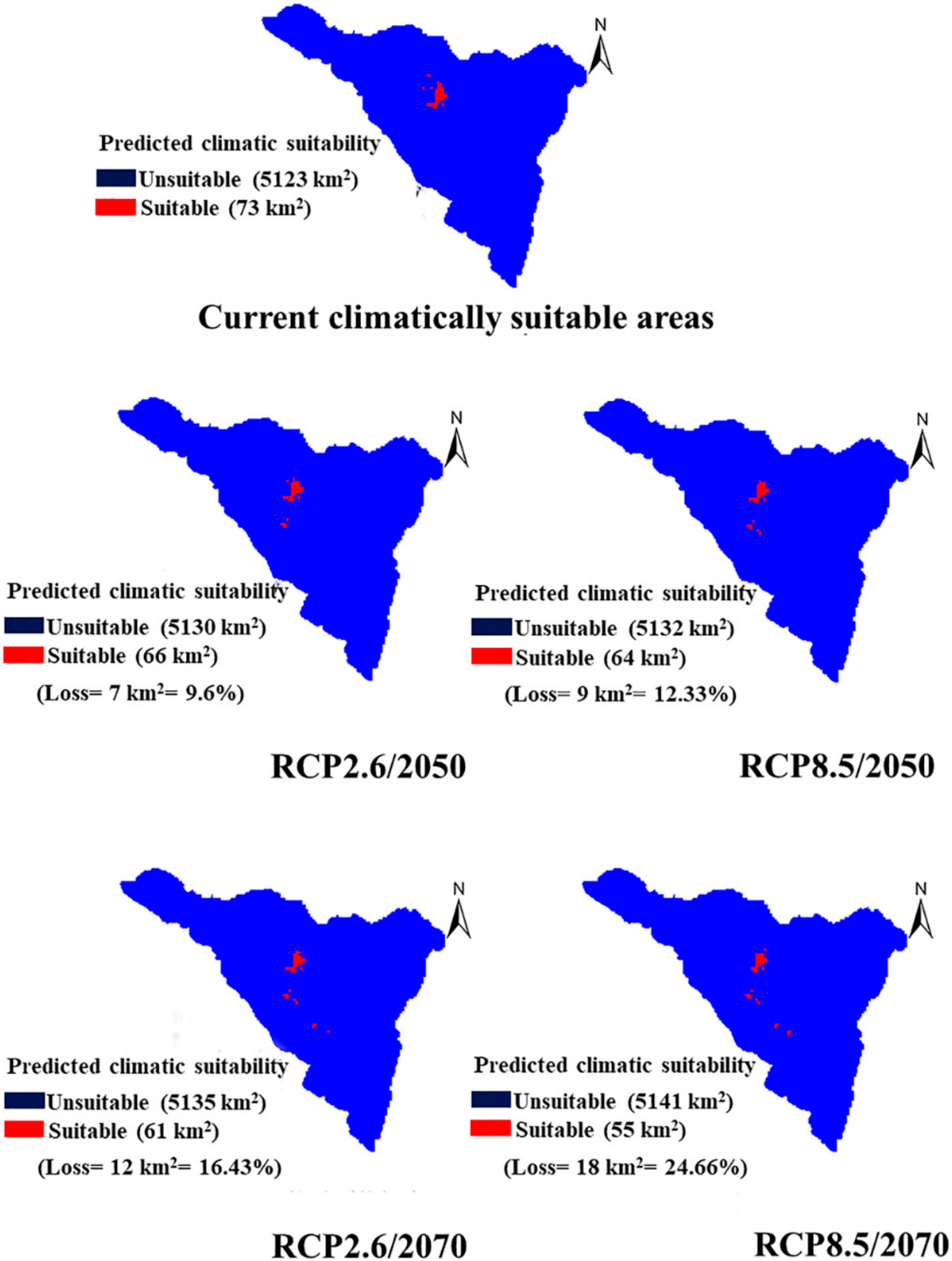
2.5. Impact of Climate Change Scenarios on the Future Distribution of P. boveana
3. Discussion
3.1. Limitations of the Study and the Best Set of Predictor Variables
3.2. The Predicted Current Suitable Sites for Survey or Translocation of P. boveana
3.3. Main Environmental Predictors for the Distribution of P. boveana
3.4. Future Predictive Distribution Area of P. boveana under Two Global Warming Scenarios
4. Materials and Methods
4.1. Study Area and Species
4.2. Environmental Predictors
4.3. MaxEnt Modeling Procedures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kier, G.; Kreft, H.; Tien, M.L.; Jetz, W.; Ibisch, P.L.; Nowicki, C.; Mutke, J.; Barthlott, W. A Global Assessment of Endemism and Species Richness across Island and Mainland Regions. Proc. Natl. Acad. Sci. USA 2009, 106, 9322–9327. [Google Scholar] [CrossRef] [PubMed]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global Biodiversity Conservation: The Critical Role of Hotspots. In Biodiversity Hotspots; Springel: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar] [CrossRef]
- Allen, J.L.; McMullin, R.T. Modeling Algorithm Influence on the Success of Predicting New Populations of Rare Species: Ground-Truthing Models for the Pale-Belly Frost Lichen (Physconia subpallida) in Ontario. Biodivers. Conserv. 2019, 28, 1853–1862. [Google Scholar] [CrossRef]
- Fois, M.; Fenu, G.; Cuena Lombraña, A.; Cogoni, D.; Bacchetta, G. A Practical Method to Speed up the Discovery of Unknown Populations Using Species Distribution Models. J. Nat. Conserv. 2015, 24, 42–48. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The Effect of Sample Size and Species Characteristics on Performance of Different Species Distribution Modeling Methods. Ecography (Cop.) 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Benito Alonso, J.L.; Coldea, G.; Dick, J.; Erschbamer, B.; Fernández Calzado, M.R.; et al. Continent-Wide Response of Mountain Vegetation to Climate Change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Wehn, S.; Johansen, L. The Distribution of the Endemic Plant Primula scandinavica, at Local and National Scales, in Changing Mountainous Environments. Biodiversity 2015, 16, 278–288. [Google Scholar] [CrossRef]
- Loarie, S.R.; Carter, B.E.; Hayhoe, K.; McMahon, S.; Moe, R.; Knight, C.A.; Ackerly, D.D. Climate Change and the Future of California’s Endemic Flora. PLoS ONE 2008, 3. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt Modeling to Predict the Potential Distribution of the Endemic Plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting Species Distribution: Offering More than Simple Habitat Models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Smeraldo, S.; Di Febbraro, M.; Bosso, L.; Flaquer, C.; Guixé, D.; Lisón, F.; Meschede, A.; Juste, J.; Prüger, J.; Puig-Montserrat, X.; et al. Ignoring Seasonal Changes in the Ecological Niche of Non-Migratory Species May Lead to Biases in Potential Distribution Models: Lessons from Bats. Biodivers. Conserv. 2018, 27, 2425–2441. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using Species Distribution Models at Local Scale to Guide the Search of Poorly Known Species: Review, Methodological Issues and Future Directions. Ecol. Modell. 2018, 385, 124–132. [Google Scholar] [CrossRef]
- Mccain, C.M.; Colwell, R.K. Assessing the Threat to Montane Biodiversity from Discordant Shifts in Temperature and Precipitation in a Changing Climate. Ecol. Lett. 2011, 14, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.-J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent Modeling for Predicting the Potential Distribution of Endangered Medicinal Plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Rus, J.D.; Ramírez-Rodríguez, R.; Amich, F.; Melendo-Luque, M. Habitat Distribution Modelling, under the Present Climatic Scenario, of the Threatened Endemic Iberian Delphinium Fissum subsp. sordidum (Ranunculaceae) and Implications for Its Conservation. Plant. Biosyst. 2018, 152, 891–900. [Google Scholar] [CrossRef]
- Wang, H.H.; Wonkka, C.L.; Treglia, M.L.; Grant, W.E.; Smeins, F.E.; Rogers, W.E. Incorporating Local-Scale Variables into Distribution Models Enhances Predictability for Rare Plant Species with Biological Dependencies. Biodivers. Conserv. 2019, 28, 171–182. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The Art of Modelling Range-Shifting Species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Alfaro-Saiz, E.; García-González, M.E.; del Río, S.; Penas, A.; Rodríguez, A.; Alonso-Redondo, R. Incorporating Bioclimatic and Biogeographic Data in the Construction of Species Distribution Models in Order to Prioritize Searches for New Populations of Threatened Flora. Plant. Biosyst. 2015, 149, 827–837. [Google Scholar] [CrossRef]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A Standard Protocol for Reporting Species Distribution Models. Ecography (Cop.) 2020, 43, 1–17. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting Species Distributions from Small Numbers of Occurrence Records: A Test Case Using Cryptic Geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Velazco, S.J.E.; Galvão, F.; Villalobos, F.; De Marco, P. Using Worldwide Edaphic Data to Model Plant Species Niches: An Assessment at a Continental Extent. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Preston, K.L.; Rotenberry, J.T.; Redak, R.A.; Allen, M.F. Habitat Shifts of Endangered Species under Altered Climate Conditions: Importance of Biotic Interactions. Glob. Chang. Biol. 2008, 14, 2501–2515. [Google Scholar] [CrossRef]
- Austin, M.P.; Van Niel, K.P. Improving Species Distribution Models for Climate Change Studies: Variable Selection and Scale. J. Biogeogr. 2011, 38, 1–8. [Google Scholar] [CrossRef]
- Coudun, C.; Gégout, J.C.; Piedallu, C.; Rameau, J.C. Soil Nutritional Factors Improve Models of Plant Species Distribution: An Illustration with Acer campestre (L.) in France. J. Biogeogr. 2006, 33, 1750–1763. [Google Scholar] [CrossRef]
- Nunes, A.; Köbel, M.; Pinho, P.; Matos, P.; Costantini, E.A.C.; Soares, C.; de Bello, F.; Correia, O.; Branquinho, C. Local Topographic and Edaphic Factors Largely Predict Shrub Encroachment in Mediterranean Drylands. Sci. Total Environ. 2019, 657, 310–318. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Liu, C. Modelling Species Distributions in Britain: A Hierarchical Integration of Climate and Land-Cover Data. Ecography (Cop.) 2004, 27, 285–298. [Google Scholar] [CrossRef]
- Ashcroft, M.B.; French, K.O.; Chisholm, L.A. An Evaluation of Environmental Factors Affecting Species Distributions. Ecol. Modell. 2011, 222, 524–531. [Google Scholar] [CrossRef]
- Bucklin, D.N.; Basille, M.; Benscoter, A.M.; Brandt, L.A.; Mazzotti, F.J.; Romañach, S.S.; Speroterra, C.; Watling, J.I. Comparing Species Distribution Models Constructed with Different Subsets of Environmental Predictors. Divers. Distrib. 2015, 21, 23–35. [Google Scholar] [CrossRef]
- Woodward, F.I. Climate and Plant Distribution. In Climate and Plant Distribution; Cambridge University Press: Cambridge, UK, 1987. [Google Scholar] [CrossRef]
- Nezer, O.; Bar-David, S.; Gueta, T.; Carmel, Y. High-Resolution Species-Distribution Model Based on Systematic Sampling and Indirect Observations. Biodivers. Conserv. 2017, 26, 421–437. [Google Scholar] [CrossRef]
- Körner, C. The Use of “Altitude” in Ecological Research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Dubuis, A.; Giovanettina, S.; Pellissier, L.; Pottier, J.; Vittoz, P.; Guisan, A. Improving the Prediction of Plant Species Distribution and Community Composition by Adding Edaphic to Topo-Climatic Variables. J. Veg. Sci. 2013, 24, 593–606. [Google Scholar] [CrossRef]
- Omar, K. Primula boveana. The IUCN Red List of Threatened Speciese 2014: e. T163968A1015883. 2014. Available online: https://dx.doi.org/10.2305/IUCN.UK.2014-3.RLTS.T163968A1015883.en (accessed on 10 February 2018).
- Crisp, M.D.; Laffan, S.; Linder, H.P.; Monro, A. Endemism in the Australian Flora. J. Biogeogr. 2001, 28, 183–198. [Google Scholar] [CrossRef]
- Gogol-Prokurat, M. Predicting Habitat Suitability for Rare Plants at Local Spatial Scales Using a Species Distribution Model. Ecol. Appl. 2011, 21, 33–47. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.R.; Leathwick, J.; Lehmann, A.; et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography (Cop.) 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E.; Elith, J.; Graham, C.H.; Phillips, S.; Peterson, A.T. What Matters for Predicting the Occurrences of Trees: Techniques, Data, or Species’ Characteristics? Ecol. Monogr. 2007, 77, 615–630. [Google Scholar] [CrossRef]
- Engler, R.; Guisan, A.; Rechsteiner, L. An Improved Approach for Predicting the Distribution of Rare and Endangered Species from Occurrence and Pseudo-Absence Data. J. Appl. Ecol. 2004, 41, 263–274. [Google Scholar] [CrossRef]
- Gu, W.; Swihart, R.K. Absent or Undetected? Effects of Non-Detection of Species Occurrence on Wildlife-Habitat Models. Biol. Conserv. 2004, 116, 195–203. [Google Scholar] [CrossRef]
- Bertrand, R.; Perez, V.; Gégout, J.C. Disregarding the Edaphic Dimension in Species Distribution Models Leads to the Omission of Crucial Spatial Information under Climate Change: The Case of Quercus pubescens in France. Glob. Chang. Biol. 2012, 18, 2648–2660. [Google Scholar] [CrossRef]
- Beauregard, F.; De Blois, S. Beyond a Climate-Centric View of Plant Distribution: Edaphic Variables Add Value to Distribution Models. PLoS ONE 2014, 9, e92642. [Google Scholar] [CrossRef]
- Diekmann, M.; Michaelis, J.; Pannek, A. Know Your Limits-The Need for Better Data on Species Responses to Soil Variables. Basic Appl. Ecol. 2015, 16, 563–572. [Google Scholar] [CrossRef]
- Condit, R.; Engelbrecht, B.M.J.; Pino, D.; Pérez, R.; Turnera, B.L. Species Distributions in Response to Individual Soil Nutrients and Seasonal Drought across a Community of Tropical Trees. Proc. Natl. Acad. Sci. USA 2013, 110, 5064–5068. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, M.C.; Gove, A.D.; Sanders, N.J.; Dunn, R.R. Climate Change, Plant Migration, and Range Collapse in a Global Biodiversity Hotspot: The Banksia (Proteaceae) of Western Australia. Glob. Chang. Biol. 2008, 14, 1337–1352. [Google Scholar] [CrossRef]
- Arundel, S.T. Using Spatial Models to Establish Climatic Limiters of Plant Species’ Distributions. Ecol. Modell. 2005, 182, 159–181. [Google Scholar] [CrossRef]
- Hageer, Y.; Esperón-Rodríguez, M.; Baumgartner, J.B.; Beaumont, L.J. Climate, Soil or Both? Which Variables Are Better Predictors of the Distributions of Australian Shrub Species? PeerJ 2017, 5, e3446. [Google Scholar] [CrossRef]
- Buri, A.; Cianfrani, C.; Pinto-Figueroa, E.; Yashiro, E.; Spangenberg, J.E.; Adatte, T.; Verrecchia, E.; Guisan, A.; Pradervand, J.N. Soil Factors Improve Predictions of Plant Species Distribution in a Mountain Environment. Prog. Phys. Geogr. 2017, 41, 703–722. [Google Scholar] [CrossRef]
- Baudraz, M.E.A.; Pradervand, J.N.; Beauverd, M.; Buri, A.; Guisan, A.; Vittoz, P. Learning from Model Errors: Can Land Use, Edaphic and Very High-Resolution Topo-Climatic Factors Improve Macroecological Models of Mountain Grasslands? J. Biogeogr. 2018, 45, 429–437. [Google Scholar] [CrossRef]
- Hosseini, S.Z.; Kappas, M.; Zare Chahouki, M.A.; Gerold, G.; Erasmi, S.; Rafiei Emam, A. Modelling Potential Habitats for Artemisia sieberi and Artemisia sucheri in Poshtkouh Area, Central Iran Using the Maximum Entropy Model and Geostatistics. Ecol. Inform. 2013, 18, 61–68. [Google Scholar] [CrossRef]
- Danin, A. Desert Vegetation of Israel and Sinai; Cana Publishing House: Jerusalem, Israel, 1983. [Google Scholar]
- Al Wadi, H. Primula boveana and Jebel Katarina. Bull. Alp. Gar Soc. 1993, 61, 68–70. [Google Scholar]
- Zaghloul, M.S. Ecological Studies on Some Endemic Plant Species in South Sinai. Master’s Thesis, Faculty of Science, Suez Canal University, Ismailia, Egypt, 1997. [Google Scholar]
- Moustafa, A.; Ramadan, A.A.; Zaghloul, M.S.; Helmy, M.A. Characteristics of Two Endemic and Endangered Species, Primula boveana and Kickxia macilenta, Growing in South Sinai Mountains, Egypt. Egypt. J. Bot. 2001, 41, 17–39. [Google Scholar]
- Mansour, H.; Jiménez, A.; Keller, B.; Nowak, M.D.; Conti, E. Development of 13 Microsatellite Markers in the Endangered Sinai Primrose (Primula boveana, Primulaceae). Appl. Plant. Sci. 2013, 1, 1200515. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, A.; Mansour, H.; Keller, B.; Conti, E. Low Genetic Diversity and High Levels of Inbreeding in the Sinai Primrose (Primula boveana), a Species on the Brink of Extinction. Plant. Syst. Evol. 2014, 300, 1199–1208. [Google Scholar] [CrossRef]
- Omar, K. Assessing the Conservation Status of the Sinai Primrose (Primula boveana). Middle East. J. Sci. Res. 2014, 21, 1027–1036. [Google Scholar]
- Omar, K.; Elgamal, I. Reproductive and Germination Ecology of Sinai Primrose, Primula boveana Decne. Ex Duby. J. Glob. Biosci. 2014, 3, 694–707. [Google Scholar]
- Zaghloul, M.S.; Moustafa, A.A.; Mauricio, R.; Mansour, H.M.H. Evolution and Conservation of Sinai’s Primrose (Primula boveana): An Endangered Pre-Glacial Relict Species Growing on Sinai Mountain, Egypt. Biog. J. 2016, 4, 56–72. [Google Scholar]
- Abolmaali, S.M.R.; Tarkesh, M.; Bashari, H. MaxEnt Modeling for Predicting Suitable Habitats and Identifying the Effects of Climate Change on a Threatened Species, Daphne Mucronata, in Central Iran. Ecol. Inform. 2018, 43, 116–123. [Google Scholar] [CrossRef]
- Khafaga, O.; Hatab, E.E.; Omar, K. Predicting the Potential Geographical Distribution of Nepeta septemcrenata in Saint Katherine Protectorate, South Sinai, Egypt Using Maxent. Acad. Arena 2011, 3, 45–50. [Google Scholar]
- Khafagi, O.; Hatab, E.E.; Omar, K. Ecological Niche Modelling as a Tool for Conservation Planning: Suitable Habitat for Hypericum sinaicum in South Sinai, Egypt. Univ. J. Environ. Res. Technol. 2012, 2, 515–524. [Google Scholar]
- Randin, C.F.; Engler, R.; Normand, S.; Zappa, M.; Zimmermann, N.E.; Pearman, P.B.; Vittoz, P.; Thuiller, W.; Guisan, A. Climate Change and Plant Distribution: Local Models Predict High-Elevation Persistence. Glob. Chang. Biol. 2009, 15, 1557–1569. [Google Scholar] [CrossRef]
- Oke, O.A.; Thompson, K.A. Distribution Models for Mountain Plant Species: The Value of Elevation. Ecol. Modell. 2015, 301, 72–77. [Google Scholar] [CrossRef]
- Moustafa, A.; Zaghloul, M.S.; EI-Wahab, R.H.A.; Shaker, M. Evaluation of Plant Diversity and Endemism in Saint Catherine Protectorate, South Sinai, Egypt. Egypt. J. Bot. 2001, 41, 121–139. [Google Scholar]
- Thuiller, W. On the Importance of Edaphic Variables to Predict Plant Species Distributions-Limits and Prospects. J. Veg. Sci. 2013, 24, 591–592. [Google Scholar] [CrossRef] [PubMed]
- Gobat, J.M.; Aragno, M.; Matthey, W. The Living Soil: Fundamentals of Soil Science and Soil Biology; Science Publishers: Enfield, NH, USA, 2004. [Google Scholar]
- Hoyle, M.; James, M. Global Warming, Human Population Pressure, and Viability of the World’s Smallest Butterfly. Conserv. Biol. 2005, 19, 1113–1124. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of Global Warming on Wild Animals and Plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Anderson, R.P. A Framework for Using Niche Models to Estimate Impacts of Climate Change on Species Distributions. Ann. N. Y. Acad. Sci. 2013, 1297, 8–28. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Biogeographical Characterisation of Egypt Based on Environmental Features and Endemic Vascular Plants Distribution. Appl. Geogr. 2020, 119, 1–10. [Google Scholar] [CrossRef]
- Ayyad, M.A.; Fakhry, A.M.; Moustafa, A.-R.A. Plant Biodiversity in the Saint Catherine Area of the Sinai Peninsula, Egypt. Biodivers. Conserv. 2000, 9, 265–281. [Google Scholar] [CrossRef]
- Moustafa, A.; Klopatek, J.M. Vegetation and Landforms of the Saint Catherine Area, Southern Sinai, Egypt. J. Arid Environ. 1995, 30, 385–395. [Google Scholar] [CrossRef]
- Moustafa, A.; Zaghloul, M.; Mansour, S.; Alsharkawy, D.; Alotaibi, M. Long Term Monitoring of Rosa arabica Populations as a Threatened Species in South Sinai, Egypt. J. Biodivers. Endanger. Species 2017, 5, 1–8. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Critical Checklist of the Endemic Vascular Plants of Egypt. Phytotaxa 2018, 360, 19–34. [Google Scholar] [CrossRef]
- Grainger, J.; Gilbert, F. Around the Sacred Mountain: The St. Katherine Protectorate in South Sinai, Egypt. In Protected Landscapes and Cultural and Spiritual Values. Values of Protected Landscapes and Seascapes; Mallarach, J.M., Ed.; Kasparek Verlag, (IUCN): Heidelberg, Germany, 2008. [Google Scholar]
- Boulos, L. Flora of Egypt. Geraniaceae-Boraginaceae; Al Hadara Publishing: Cairo, Egypt, 2000. [Google Scholar]
- Richards, A.J. Primula, 2nd ed.; Timber Press: Portland, ON, USA, 2003. [Google Scholar]
- Boulos, L. Flora of Egypt Checklist, Revised Annotated Edition; Al Hadara Publishing: Cairo, Egypt, 2009. [Google Scholar]
- Alfaro-Saiz, E.; Granda, V.; Rodríguez, A.; Alonso-Redondo, R.; García-González, M.E. Optimal Census Method to Estimate Population Sizes of Species Growing on Rock Walls: The Case of Mature Primula pedemontana. Glob. Ecol. Conserv. 2019, 17, e00563. [Google Scholar] [CrossRef]
- Shaltout, K.H.; Ahmed, D.A.; Shabana, H.A. Population Structure and Dynamics of the Endemic Species Phlomis aurea Decne in Different Habitats in Southern Sinai Peninsula, Egypt. Glob. Ecol. Conserv. 2015, 4, 505–515. [Google Scholar] [CrossRef][Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Title, P.O.; Bemmels, J.B. ENVIREM: An Expanded Set of Bioclimatic and Topographic Variables Increases Flexibility and Improves Performance of Ecological Niche Modeling. Ecography (Cop.) 2018, 41, 291–307. [Google Scholar] [CrossRef]
- Amatulli, G.; Domisch, S.; Tuanmu, M.N.; Parmentier, B.; Ranipeta, A.; Malczyk, J.; Jetz, W. Data Descriptor: A Suite of Global, Cross-Scale Topographic Variables for Environmental and Biodiversity Modeling. Sci. Data 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Hengl, T.; De Jesus, J.M.; MacMillan, R.A.; Batjes, N.H.; Heuvelink, G.B.M.; Ribeiro, E.; Samuel-Rosa, A.; Kempen, B.; Leenaars, J.G.B.; Walsh, M.G.; et al. SoilGrids1km-Global Soil Information Based on Automated Mapping. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Naimi, B.; Araújo, M.B. Sdm: A Reproducible and Extensible R Platform for Species Distribution Modelling. Ecography (Cop.) 2016, 39, 368–375. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2018. Available online: https://www.R-project.org (accessed on 10 April 2019).
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Sanjerehei, M.; Rundel, P.W. The Impact of Climate Change on Habitat Suitability for Artemisia sieberi and Artemisia aucheri (Asteraceae), a Modeling Approach. Polish J. Ecol. 2017, 65, 97–109. [Google Scholar] [CrossRef]
- Al-Qaddi, N.; Vessella, F.; Stephan, J.; Al-Eisawi, D.; Schirone, B. Current and Future Suitability Areas of Kermes Oak (Quercus coccifera L.) in the Levant under Climate Change. Reg. Environ. Chang. 2017, 17, 143–156. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Ahmadi, M. Climate Modelling Shows Increased Risk to Eucalyptus sideroxylon on the Eastern Coast of Australia Compared to Eucalyptus albens. Plants 2017, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Hoveka, L.N.; Bezeng, B.S.; Yessoufou, K.; Boatwright, J.S.; Van der Bank, M. Effects of Climate Change on the Future Distributions of the Top Five Freshwater Invasive Plants in South Africa. S. Afr. J. Bot. 2016, 102, 33–38. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography (Cop.) 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of Methods for the Assessment of Prediction Errors in Conservation Presence/Absence Models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the Accuracy of Species Distribution Models: Prevalence, Kappa and the True Skill Statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent Modeling for Predicting Impacts of Climate Change on the Potential Distribution of Thuja sutchuenensis Franch. an Extremely Endangered Conifer from Southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent Modeling for Predicting the Potential Distribution of Medicinal Plant, Justicia adhatoda L. in Lesser Himalayan Foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting Thresholds of Occurrence in the Prediction of Species Distributions. Ecography (Cop.) 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next Generation Python-Based GIS Toolkit for Landscape Genetic, Biogeographic and Species Distribution Model Analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Location | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Wadi Shaq Mousa (WSM) | Wadi Garagniah (WG) | Maen Shennarah (MS) | Kahf El-Ghoula (KG) | Gebal Alahmar (GA) | Sad Abu Hebeik (SH) | ||||||
| Population parameters | |||||||||||
| Total individuals | 408.3 ± 12.6 | 77.7 ± 1.5 | 61.3 ± 3.1 | 74.7 ± 0.6 | 91.6 ± 1.5 | 10.0 ± 1.0 | 54.0 ± 1.0 | 4.0 ± 0.0 | 7.0 ± 0.0 | 5.0 ± 0.0 | |
| Mature individuals | 55.0 ± 2.0 | 15.0 ± 2.0 | 8.0 ± 1.0 | 11.3 ± 1.5 | 22.7 ± 1.5 | 2.7 ± 0.6 | 14.0 ± 1.0 | 0.7 ± 0.6 | 5.0 ± 1.0 | 0.7 ± 0.6 | |
| Density (individuals/25 m2) | 16.8 ± 1.3 | 3.1 ± 0.3 | 2.5 ± 0.1 | 3.0 ± 0.5 | 3.7 ± 0.8 | 0.4 ± 0.0 | 2.2 ± 0.2 | 0.2 ± 0.0 | 0.3 ± 0.0 | 0.2 ± 0.0 | |
| Cover (%) | 45.0 ± 1.5 | 25.0 ± 0.5 | 26.7 ± 2.5 | 25.0 ± 3.0 | 35.0 ± 5.0 | 5.0 ± 0.9 | 25.0 ± 1.0 | 2.0 ± 0.0 | 5.0 ± 0.0 | 1.0 ± 0.0 | |
| Size index (cm) | 22.7 ± 2.1 | 12.0 ± 2.6 | 16.0 ± 3.0 | 12.3 ± 1.5 | 10.0 ± 2.6 | 14.3 ± 2.0 | 16.3 ± 1.5 | 12.0 ± 2.0 | 10.0 ± 2.0 | 12.0 ± 2.6 | |
| Plant vigor | 1.1 ± 0.2 | 1.2 ± 0.6 | 1.4 ± 0.0 | 1.6 ± 0.2 | 2.7 ± 0.5 | 1.1 ± 0.0 | 1.3 ± 0.0 | 1.0 ± 0.0 | 1.6 ± 0.2 | 1.0 ± 0.1 | |
| Topography | Elevation (m) | 2065 | 2050 | 1950 | 2165 | 2208 | 1890 | 2032 | 1803 | 1915 | 1687 |
| Slope (degree) | 90 | 90 | 85 | 70 | 90 | 60 | 90 | 90 | 80 | 90 | |
| Aspect (degree, direction) | 40.3 (NE) | 25.2 (NE) | 30.3 (NE) | 33.2 (NE) | 25.5 (NE) | 30.3 (NE) | 45.5 (NE) | 69.7 (E) | 70.2 (E) | 27.9 (NE) | |
| Microhabitat | gorge | gorge | gorge | gorge | slope | gorge | gorge | cave | slope | gorge | |
| Climate-Only Model | Topography-Only Model | Edaphic-Only Model | Climate-Topography Model | Climate-Edaphic Model | Topography-Edaphic Model | Climate-Topography-Edaphic Model | |
|---|---|---|---|---|---|---|---|
| Model performance | |||||||
| AUC training | 0.991 bcd ± 0.00 | 0.990 abc ±0.00 | 0.845 a ± 0.01 | 0.992 cd ± 0.00 | 0.990 cd ± 0.00 | 0.989 ab ± 0.00 | 0.993 d ± 0.00 |
| AUC test | 0.989 b ± 0.01 | 0.990 b ± 0.01 | 0.833 a ± 0.07 | 0.991 b ± 0.01 | 0.985 ab ± 0.01 | 0.987 ab ± 0.01 | 0.990 b ± 0.01 |
| TSS | 0.889 cd ± 0.07 | 0.884 abc ± 0.02 | 0.725 a ± 0.30 | 0.893 b ± 0.05 | 0.887 ab ± 0.09 | 0.882 bc ± 0.20 | 0.895 d ± 0.02 |
| Average percent contribution | |||||||
| Bio6 (°C) | 25.2 | 15.1 | 12.8 | 15.2 | |||
| Bio7 (°C) | 3.8 | 0.6 | 1.4 | 1 | |||
| Bio14 (mm) | 40.4 | 24.5 | 31.7 | 18.1 | |||
| Bio15 | 28.6 | 17.5 | 25.1 | 16.4 | |||
| Aridity index | 2 | 0 | 1.8 | 0.2 | |||
| Elev. (m) | 80.3 | 38.3 | 68.4 | 30.4 | |||
| Slope (degree) | 16.1 | 4 | 3 | 4 | |||
| Aspect (degree) | 3.6 | 0 | 0.1 | ||||
| BD (g/cm3) | 2.4 | 0.2 | 0 | 0.6 | |||
| Clay (%) | 22 | 1.3 | 6.2 | 3.5 | |||
| OC (g/kg) | 36.2 | 10.1 | 7.3 | 4.6 | |||
| pH | 39.4 | 15.5 | 13 | 6 | |||
| Predicted habitat suitability class | |||||||
| <0.20 | 4816 | 4158 | 1804 | 4210 | 4000 | 2697 | 4303 |
| 0.20–0.40 | 201 | 698 | 1980 | 679 | 722 | 813 | 632 |
| 0.40–0.60 | 106 | 262 | 1090 | 225 | 296 | 332 | 192 |
| >0.60 | 73 | 78 | 322 | 82 | 178 | 159 | 69 |
| Category | Code/Unit | Predictors | VIF | Source and Resolution |
|---|---|---|---|---|
| Bioclimatic | Bio6 (°C) | Min temperature of coldest month | 4.80 | WorldClim v.2 (~1 km2). |
| Bio7 (°C) | Temperature annual range (Bio5-Bio6) | 2.73 | ||
| Bio14 (mm) | Precipitation of driest month | 2.30 | ||
| Bio15 (unitless) | Precipitation seasonality (coefficient of variation) | 4.23 | ||
| Aridity index (unitless) | Degree of water deficit below water need | 3.65 | ENVIREM (~1 km2). | |
| Topographic | Elev (m. a.s.l) | Elevation | 3.62 | DIVA-GIS (90 m) |
| Slope (degree) | Slope | 1.58 | Derived from Elev. | |
| Aspect (degree) | Aspect | 4.17 | Derived from Elev. | |
| Edaphic | BD (g/cm3) | Bulk density | 3.18 | SoilGrids (1 km2) |
| Clay (%) | Clay content | 3.44 | ||
| OC (g/kg) | Organic carbon content | 4.50 | ||
| pH | pH in H2O | 1.38 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelaal, M.; Fois, M.; Dakhil, M.A.; Bacchetta, G.; El-Sherbeny, G.A. Predicting the Potential Current and Future Distribution of the Endangered Endemic Vascular Plant Primula boveana Decne. ex Duby in Egypt. Plants 2020, 9, 957. https://doi.org/10.3390/plants9080957
Abdelaal M, Fois M, Dakhil MA, Bacchetta G, El-Sherbeny GA. Predicting the Potential Current and Future Distribution of the Endangered Endemic Vascular Plant Primula boveana Decne. ex Duby in Egypt. Plants. 2020; 9(8):957. https://doi.org/10.3390/plants9080957
Chicago/Turabian StyleAbdelaal, Mohamed, Mauro Fois, Mohammed A. Dakhil, Gianluigi Bacchetta, and Ghada A. El-Sherbeny. 2020. "Predicting the Potential Current and Future Distribution of the Endangered Endemic Vascular Plant Primula boveana Decne. ex Duby in Egypt" Plants 9, no. 8: 957. https://doi.org/10.3390/plants9080957
APA StyleAbdelaal, M., Fois, M., Dakhil, M. A., Bacchetta, G., & El-Sherbeny, G. A. (2020). Predicting the Potential Current and Future Distribution of the Endangered Endemic Vascular Plant Primula boveana Decne. ex Duby in Egypt. Plants, 9(8), 957. https://doi.org/10.3390/plants9080957