Leaves of White Beetroot As a New Source of Antioxidant and Anti-Inflammatory Compounds

 ,
,  , and
, and 
Abstract
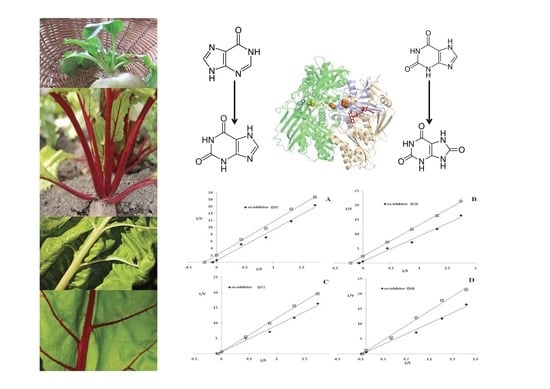
1. Introduction
2. Results
2.1. Analysis of Phenolic Compounds
2.2. Total Betalain Content
2.3. Antioxidant Capacity
2.4. Lipoxygenase Inhibition
2.5. Xanthine Oxidase Inhibition
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Extract Preparations
4.3. Total Phenolics (TPC) Estimation
4.4. Total Flavonoids (TFC) Estimation
4.5. Phenolic Acids (PA) Profile
4.6. Quantitation of Betalain Content
4.7. Antioxidant Assay
4.7.1. Ability to Scavenge ABTS Radicals
4.7.2. Ability to Scavenge the Hydroxyl (OH•) Radicals
4.7.3. Metal Chelating Activity (CHP)
4.8. Inhibition of Lipoxygenase Activity (LOXI)
4.9. Inhibition of Xanthine Oxidase Activity (XOI)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J.H.; Son, C.W.; Kim, M.Y.; Kim, M.H.; Kim, H.R.; Kwak, E.S.; Kim, S.; Kim, M.R. Red beet (Beta vulgaris L.) leaf supplementation improves antioxidant status in C57BL/6J mice fed high fat high cholesterol diet. Nutr. Res. Pract. 2009, 3, 114. [Google Scholar] [CrossRef] [PubMed]
- Pyo, Y.H.; Lee, T.C.; Logendra, L.; Rosen, R.T. Antioxidant activity and phenolic compounds of Swiss chard (Beta vulgaris subspecies cycla) extracts. Food Chem. 2004, 85, 19–26. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J.D. Bioactivities of phenolics by focusing on suppression of chronic diseases: A review. Int. J. Mol. Sci. 2018, 19, 1573. [Google Scholar] [CrossRef] [PubMed]
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E.J. The potential benefits of red beetroot supplementation in health and disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef] [PubMed]
- Treml, J.; Šmejkal, K. Flavonoids as Potent Scavengers of Hydroxyl Radicals. Compr. Rev. Food Sci. Food Saf. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef]
- Kopp, W. How western diet and lifestyle drive the pandemic of obesity and civilization diseases. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 2221–2236. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Schneider, C.; Pozzi, A. Cyclooxygenases and lipoxygenases in cancer. Cancer Metastasis Rev. 2011, 30, 277–294. [Google Scholar] [CrossRef]
- Battelli, M.G.; Bortolotti, M.; Polito, L.; Bolognesi, A. Metabolic syndrome and cancer risk: The role of xanthine oxidoreductase. Redox Biol. 2019, 21. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Koubaier, H.B.H.; Snoussi, A.; Essaidi, I.; Chaabouni, M.M.; Thonart, P.; Bouzouita, N. Betalain and phenolic compositions, antioxidant activity of Tunisian red beet (Beta vulgaris L. conditiva) roots and stems extracts. Int. J. Food Prop. 2014, 17, 1934–1945. [Google Scholar] [CrossRef]
- Zein, H.; Hashish, A.E.; Ismaiel, G. The antioxidant and Anticancer Activities of Swiss Chard and Red Beetroot Leaves. Curr. Sci. Int. 2015, 4, 491–498. [Google Scholar]
- Sacan, O.; Yanardag, R. Antioxidant and antiacetylcholinesterase activities of chard (Beta vulgaris L. var. cicla). Food Chem. Toxicol. 2010, 48, 1275–1280. [Google Scholar] [CrossRef]
- Ninfali, P.; Angelino, D. Nutritional and functional potential of Beta vulgaris cicla and rubra. Fitoterapia 2013, 89, 188–199. [Google Scholar] [CrossRef]
- Chandra, A.K.; Mondal, C.; Sinha, S.; Chakraborty, A.; Pearce, E.N. Synergic actions of polyphenols and cyanogens of peanut seed coat (Arachis hypogaea) on cytological, biochemical and functional changes in thyroid. Indian J. Exp. Biol. 2015, 53, 143–151. [Google Scholar]
- Gliszczyńska-Świgło, A.; Szymusiak, H.; Malinowska, P. Betanin, the main pigment of red beet: Molecular origin of its exceptionally high free radical-scavenging activity. Food Addit. Contam. 2006, 23, 1079–1087. [Google Scholar] [CrossRef]
- Kugler, F.; Stintzing, F.C.; Carle, R. Identification of Betalains from Petioles of Differently Colored Swiss Chard (Beta vulgaris L. ssp. cicla [L.] Alef. Cv. Bright Lights) by High-Performance Liquid Chromatography-Electrospray Ionization Mass Spectrometry. J. Agric. Food Chem. 2004, 52, 2975–2981. [Google Scholar] [CrossRef]
- Ali, M.B.; Khandaker, L.; Oba, S. Comparative study on functional components, antioxidant activity and color parameters of selected colored leafy vegetables as affected by photoperiods. J. Food Agric. Environ. 2009, 7, 392–398. [Google Scholar]
- Huyut, Z.; Beydemir, Ş.; Gülçin, I. Antioxidant and antiradical properties of selected flavonoids and phenolic compounds. Biochem. Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Burri, S.C.M.; Ekholm, A.; Håkansson, Å.; Tornberg, E.; Rumpunen, K. Antioxidant capacity and major phenol compounds of horticultural plant materials not usually used. J. Funct. Foods 2017, 38, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Britton, R.S.; Leicester, K.L.; Bacon, B.R. Iron toxicity and chelation therapy. Int. J. Hematol. 2002, 76, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Cherrak, S.A.; Mokhtari-Soulimane, N.; Berroukeche, F.; Bensenane, B.; Cherbonnel, A.; Merzouk, H.; Elhabiri, M. In Vitro antioxidant versus metal ion chelating properties of flavonoids: A structure-activity investigation. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]
- Canabady-Rochelle, L.L.S.; Harscoat-Schiavo, C.; Kessler, V.; Aymes, A.; Fournier, F.; Girardet, J.M. Determination of reducing power and metal chelating ability of antioxidant peptides: Revisited methods. Food Chem. 2015, 183, 129–135. [Google Scholar] [CrossRef]
- Mashima, R.; Okuyama, T. The role of lipoxygenases in pathophysiology; new insights and future perspectives. Redox Biol. 2015, 6, 297–310. [Google Scholar] [CrossRef]
- Peng, X.; Ma, J.; Cheng, K.-W.; Jiang, Y.; Chen, F.; Wang, M. The effects of grape seed extract fortification on the antioxidant activity and quality attributes of bread. Food Chem. 2010, 119, 49–53. [Google Scholar] [CrossRef]
- Schneider, I.; Bucar, F. Lipoxygenase inhibitors from natural plant sources. Part 1: Medicinal plants with inhibitory activity on arachidonate 5-lipoxygenase and 5-lipoxygenase[sol ]cyclooxygenase. Phytother. Res. 2005, 19, 81–102. [Google Scholar] [CrossRef]
- Miguel, M.G. Betalains in some species of the amaranthaceae family: A review. Antioxidants 2018, 7, 53. [Google Scholar] [CrossRef]
- Vidal, P.J.; López-Nicolás, J.M.; Gandía-Herrero, F.; García-Carmona, F. Inactivation of lipoxygenase and cyclooxygenase by natural betalains and semi-synthetic analogues. Food Chem. 2014, 154, 246–254. [Google Scholar] [CrossRef]
- Mehmood, A.; Zhao, L.; Wang, C.; Nadeem, M.; Raza, A.; Ali, N.; Shah, A.A. Management of hyperuricemia through dietary polyphenols as a natural medicament: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1433–1455. [Google Scholar] [CrossRef] [PubMed]
- Vulić, J.J.; Ćebović, T.N.; Čanadanović-Brunet, J.M.; Ćetković, G.S.; Čanadanović, V.M.; Djilas, S.M.; Tumbas Šaponjac, V.T. In Vivo and In Vitro antioxidant effects of beetroot pomace extracts. J. Funct. Foods 2014, 6, 168–175. [Google Scholar] [CrossRef]
- Kong, L.D.; Cai, Y.; Huang, W.W.; Cheng, C.H.K.; Tan, R.X. Inhibition of xanthine oxidase by some Chinese medicinal plants used to treat gout. J. Ethnopharmacol. 2000, 73, 199–207. [Google Scholar] [CrossRef]
- Chen, C.-H.; Chan, H.-C.; Chu, Y.-T.; Ho, H.-Y.; Chen, P.-Y.; Lee, T.-H.; Lee, C.-K. Antioxidant activity of some plant extracts towards xanthine oxidase, lipoxygenase and tyrosinase. Molecules 2009, 14, 2947–2958. [Google Scholar] [CrossRef]
- Dew, T.P.; Day, A.J.; Morgan, M.R.A. Xanthine oxidase activity In Vitro: Effects of food extracts and components. J. Agric. Food Chem. 2005, 53, 6510–6515. [Google Scholar] [CrossRef]
- Moyo, M.; Amoo, S.O.; Ncube, B.; Ndhlala, A.R.; Finnie, J.F.; Van Staden, J. Phytochemical and antioxidant properties of unconventional leafy vegetables consumed in southern Africa. S. Afr. J. Bot. 2013, 84, 65–71. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Bahorun, T.; Luximon-Ramma, A.; Crozier, A.; Aruoma, O.I. Total phenol, flavonoid, proanthocyanidin and vitamin C levels and antioxidant activities of Mauritian vegetables. J. Sci. Food Agric. 2004, 84, 1553–1561. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Świeca, M.; Dziki, D. Comparison of phenolic acids profile and antioxidant potential of six varieties of spelt (Triticum spelta L.). J. Agric. Food Chem. 2012, 60, 4603–4612. [Google Scholar] [CrossRef]
- Nilsson, T. Studies into the pigments in beetroot (Beta vulgaris L. ssp. vulgaris var. rubra L.). Lantbr. Ann. 1970, 36, 179–219. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Su, X.Y.; Wang, Z.Y.; Liu, J.R. In Vitro and in Vivo antioxidant activity of Pinus koraiensis seed extract containing phenolic compounds. Food Chem. 2009, 117, 681–686. [Google Scholar] [CrossRef]
- Guo, J.T.; Lee, H.L.; Chiang, S.H.; Lin, F.I.; Chang, C.Y. Antioxidant Properties of the Extracts from Different Parts of Broccoli in Taiwan. J. Food Drug Anal. 2001, 9, 96–101. [Google Scholar]
- Maiga, A.; Malterud, K.E.; Diallo, D.; Paulsen, B.S. Antioxidant and 15-lipoxygenase inhibitory activities of the Malian medicinal plants Diospyros abyssinica (Hiern) F. White (Ebenaceae), Lannea velutina A. Rich (Anacardiaceae) and Crossopteryx febrifuga (Afzel) Benth. (Rubiaceae). J. Ethnopharmacol. 2006, 104, 132–137. [Google Scholar] [CrossRef]
- Axelrod, B.; Cheesbrough, T.M.; Laakso, S. Lipoxygenase from Soybeans: EC 1.13.11.12 Linoleate: Oxygen oxidoreductase. Methods Enzymol. 1981, 71, 441–451. [Google Scholar]
- Sweeney, A.P.; Wyllie, S.G.; Shalliker, R.A.; Markham, J.L. Xanthine oxidase inhibitory activity of selected Australian native plants. J. Ethnopharmacol. 2001, 75, 273–277. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Acid | Red Chard (LC) | White Beetroot (SK) | Red Beetroot (CC) | White Chard (BL) |
|---|---|---|---|---|
| Protocatechuic | 2.83 ± 0.03 a * | 1.20 ± 0.00 c * | 1.72 ± 0.05 b * | nd |
| p-OH benzoic | 2.26 ± 0.06 a | 1.21 ± 0.01 b | 0.96 ± 0.00 c | 1.37 ± 0.01 b * |
| Caffeic | 2.11 ± 0.01 a | 1.79 ± 0.01 b | nd | nd |
| Syryngic | 1.45 ± 0.01 b | 1.36 ± 0.01 c | 1.74 ± 0.06 a | 1.34 ± 0.01 c |
| Vanilic | 1.43 ± 0.02 a | 1.36 ± 0.00 a | nd | nd |
| p-coumaric | 1.40 ± 0.00 a | 1.32 ± 0.00 a | nd | nd |
| Ferulic | 5.32 ± 0.17 a | 2.73 ± 0.05 c | 4.72 ± 0.21 b | 2.81 ± 0.04 c |
| Synapic | 7.92 ± 0.45 c | 23.23 ± 1.29 a | 14.90 ± 1.16 b | 4.59 ± 0.09 d |
| Salicylic | 159.01 ± 9.98 a | 108.64 ± 6.23 c | 94.56 ± 7.60 d | 142.24 ± 9.91 b |
| Sum | 180.90 a | 141.64 c | 116.89 d | 152.35 b |
| Plant | Mode of Inhibition | EC50 [mg FW/mL] | Ki [mg FW/mL] | Vmax [ΔAU/min] |
|---|---|---|---|---|
| White beetroot | uncompetitive | 72.29 ± 0.85 b | 72.01 ± 2.11 b | 416.66 ± 8.32 b |
| Red beetroot | uncompetitive | 56.97 ± 1.05 c | 55.99 ± 1.21 c | 412.58 ± 7.25 c |
| White chard | uncompetitive | 53.23 ± 2.23 d | 49.13 ± 1.02 d | 400.02 ± 5.55 d |
| Red chard | uncompetitive | 78.29 ± 1.13 a | 78.61 ± 3.22 a | 434.32 ± 6.25 a |
| Plant | Mode of Inhibition | EC50 [mg FW/mL] | Ki [mg FW/mL] | Vmax [ΔAU/min] |
|---|---|---|---|---|
| White beetroot | uncompetitive | 81.04 ± 2.22 d | 24.62 ± 0.14 d | 0.42 ± 0.01 c |
| Red beetroot | noncompetitive | 205.09 ± 4.58 a | 1020.48 ± 13.58 a | 1.5 ± 0.02 a |
| White chard | noncompetitive | 147.28 ± 5.32 b | 229.71 ± 8.23 b | 1.11 ± 0.4 b |
| Red chard | uncompetitive | 105.32 ± 7.41 c | 39.156 ± 1.54 c | 0.49 ± 0.01 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gawlik-Dziki, U.; Dziki, L.; Anisiewicz, J.; Habza-Kowalska, E.; Sikora, M.; Dziki, D. Leaves of White Beetroot As a New Source of Antioxidant and Anti-Inflammatory Compounds. Plants 2020, 9, 944. https://doi.org/10.3390/plants9080944
Gawlik-Dziki U, Dziki L, Anisiewicz J, Habza-Kowalska E, Sikora M, Dziki D. Leaves of White Beetroot As a New Source of Antioxidant and Anti-Inflammatory Compounds. Plants. 2020; 9(8):944. https://doi.org/10.3390/plants9080944
Chicago/Turabian StyleGawlik-Dziki, Urszula, Laura Dziki, Jakub Anisiewicz, Ewa Habza-Kowalska, Małgorzata Sikora, and Dariusz Dziki. 2020. "Leaves of White Beetroot As a New Source of Antioxidant and Anti-Inflammatory Compounds" Plants 9, no. 8: 944. https://doi.org/10.3390/plants9080944
APA StyleGawlik-Dziki, U., Dziki, L., Anisiewicz, J., Habza-Kowalska, E., Sikora, M., & Dziki, D. (2020). Leaves of White Beetroot As a New Source of Antioxidant and Anti-Inflammatory Compounds. Plants, 9(8), 944. https://doi.org/10.3390/plants9080944