Comparative Study of the Effects of Salinity on Growth, Gas Exchange, N Accumulation and Stable Isotope Signatures of Forage Oat (Avena sativa L.) Genotypes




Abstract
1. Introduction
2. Results
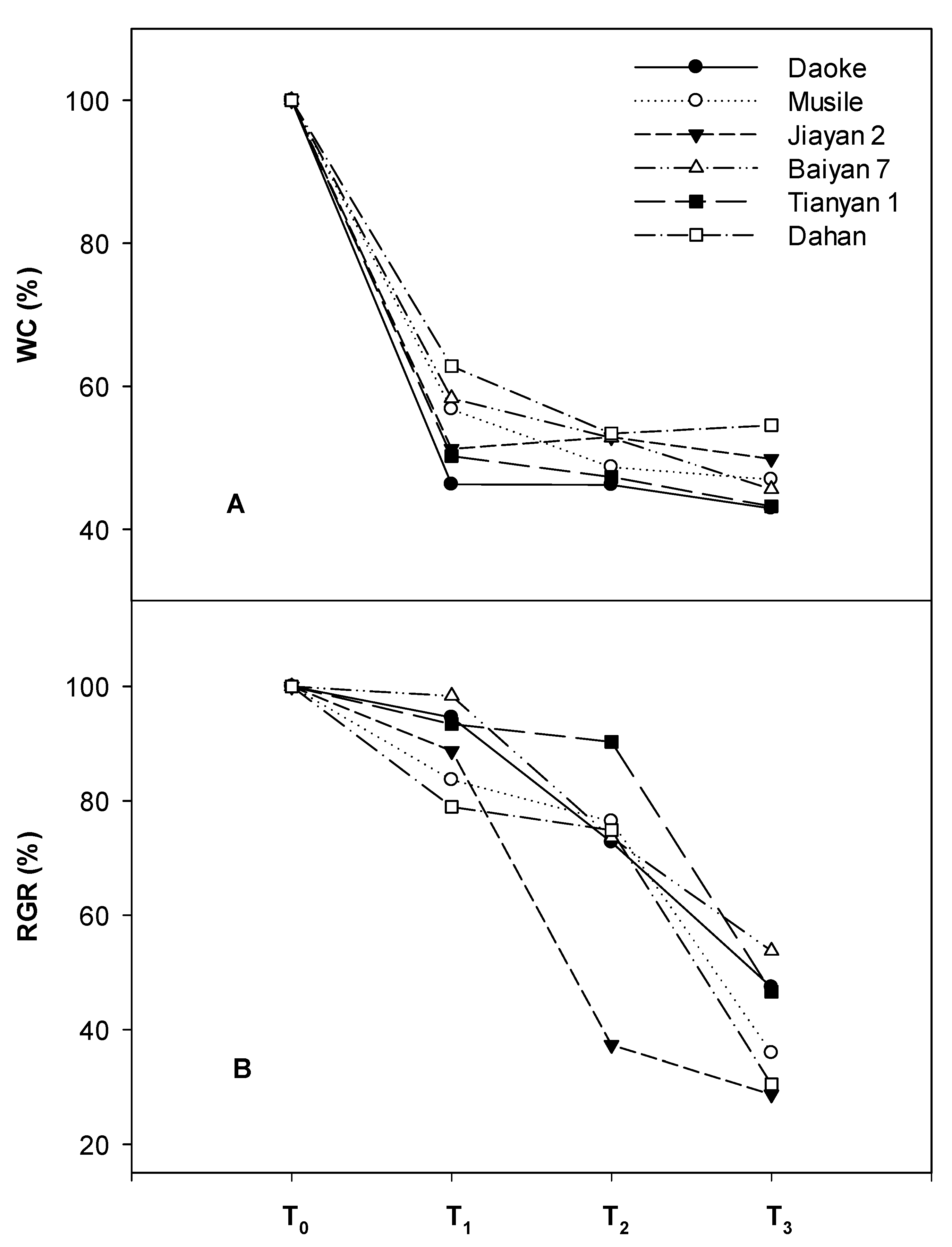
2.1. Biomass and Related Growth Traits
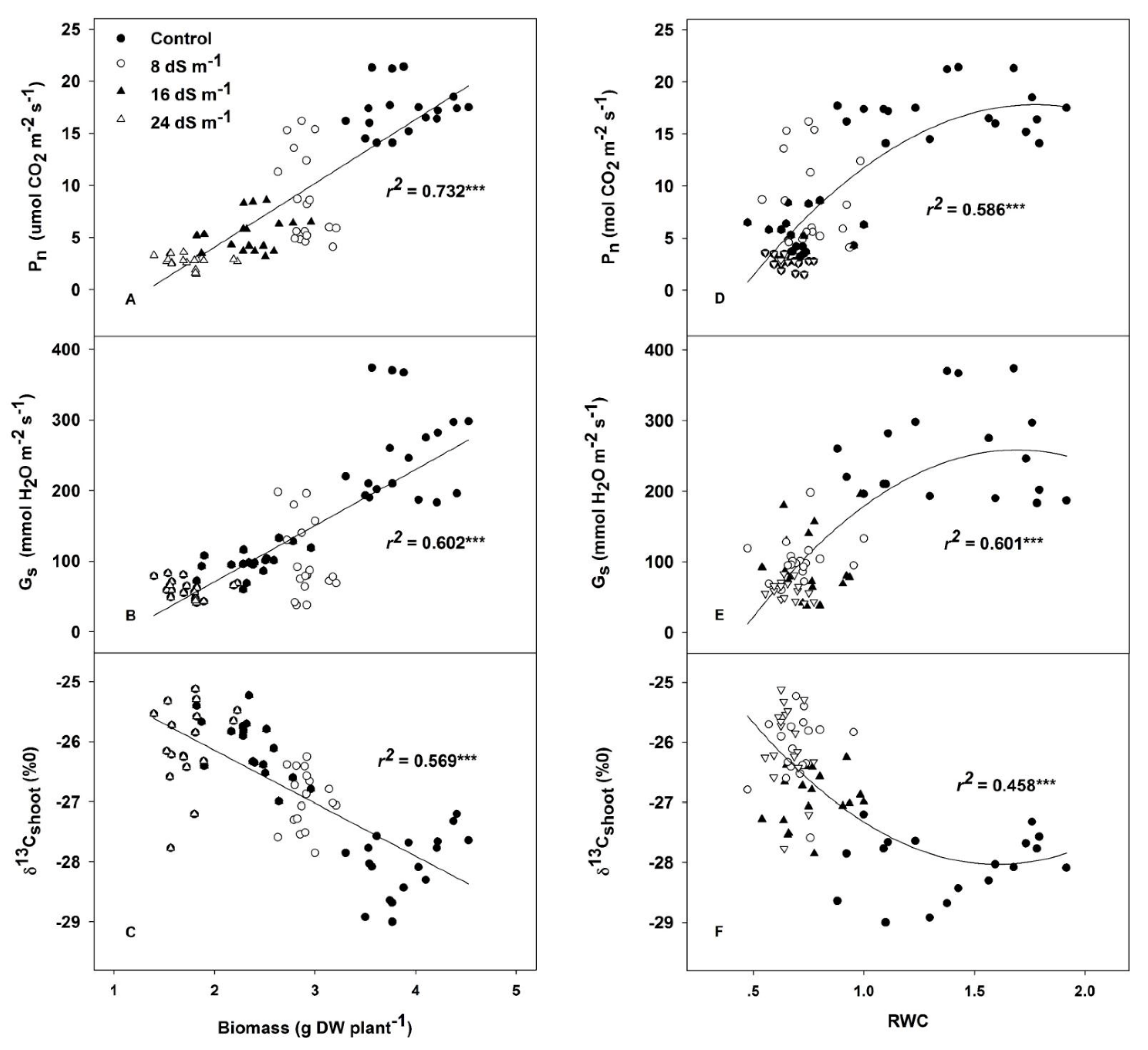
2.2. Gas Exchange Parameters and the Related Water Status
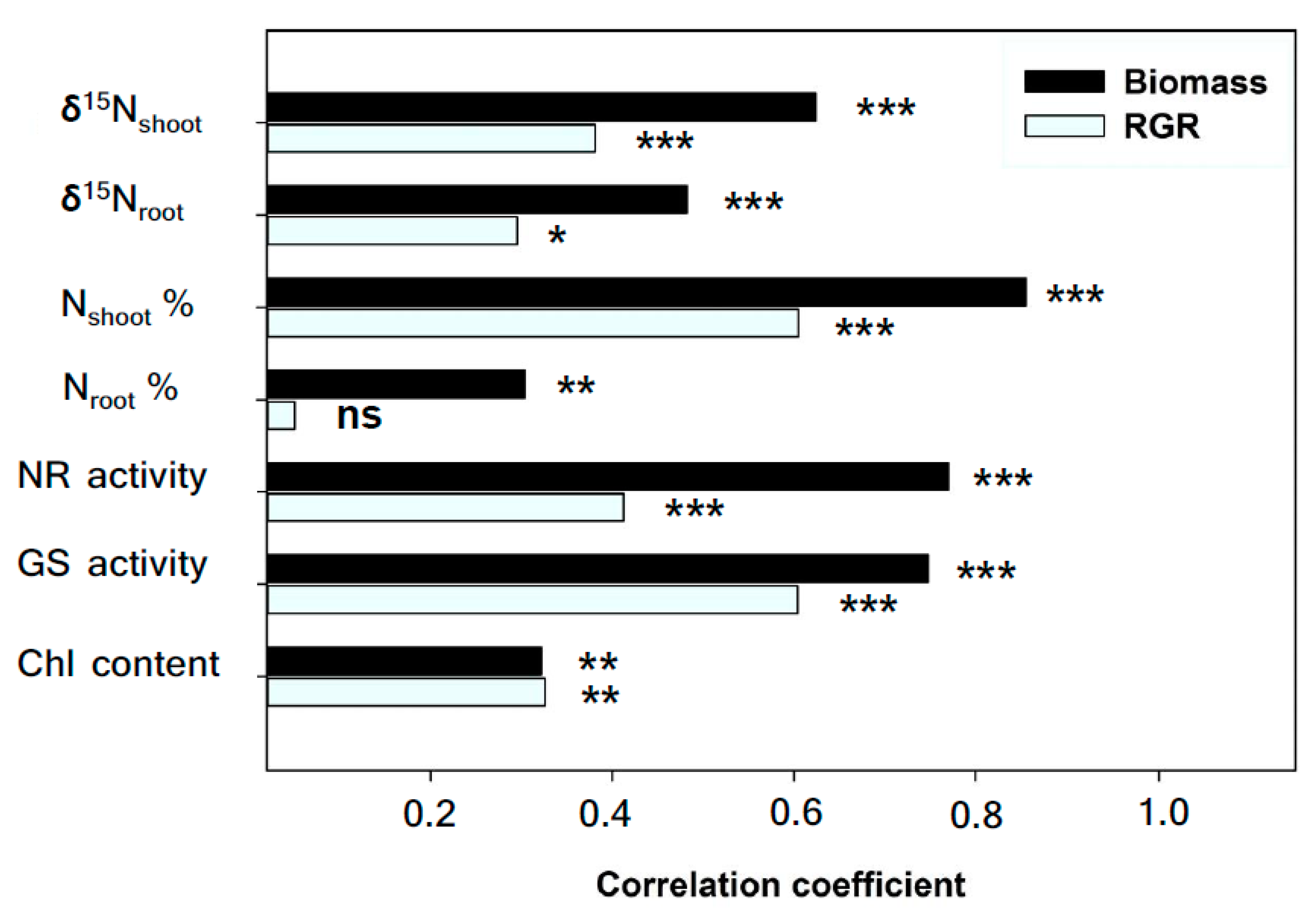
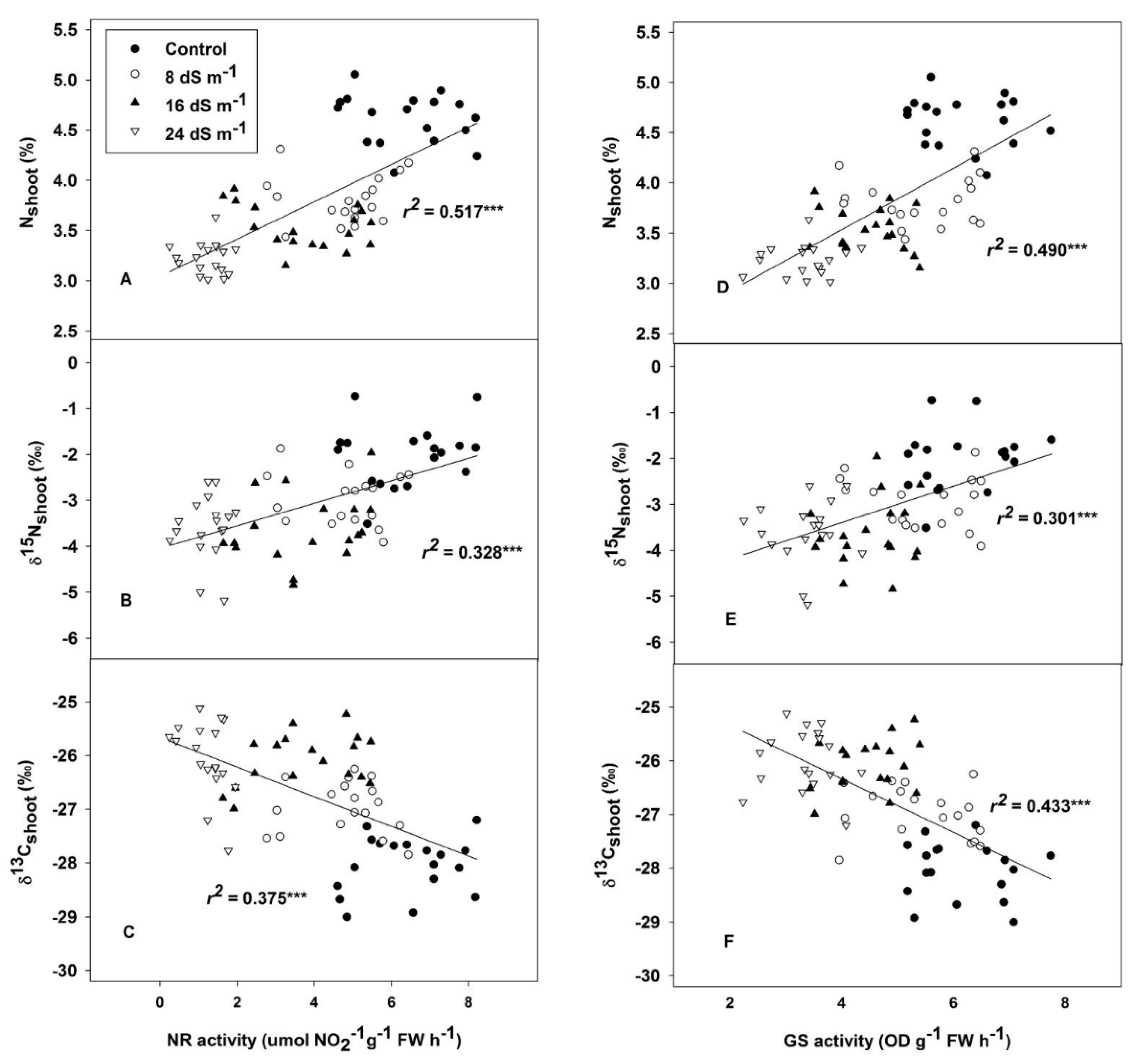
2.3. Nitrogen Concentration, Nitrogen Stable Isotopes and Related Enzymes
2.4. Ion Concentration Determination
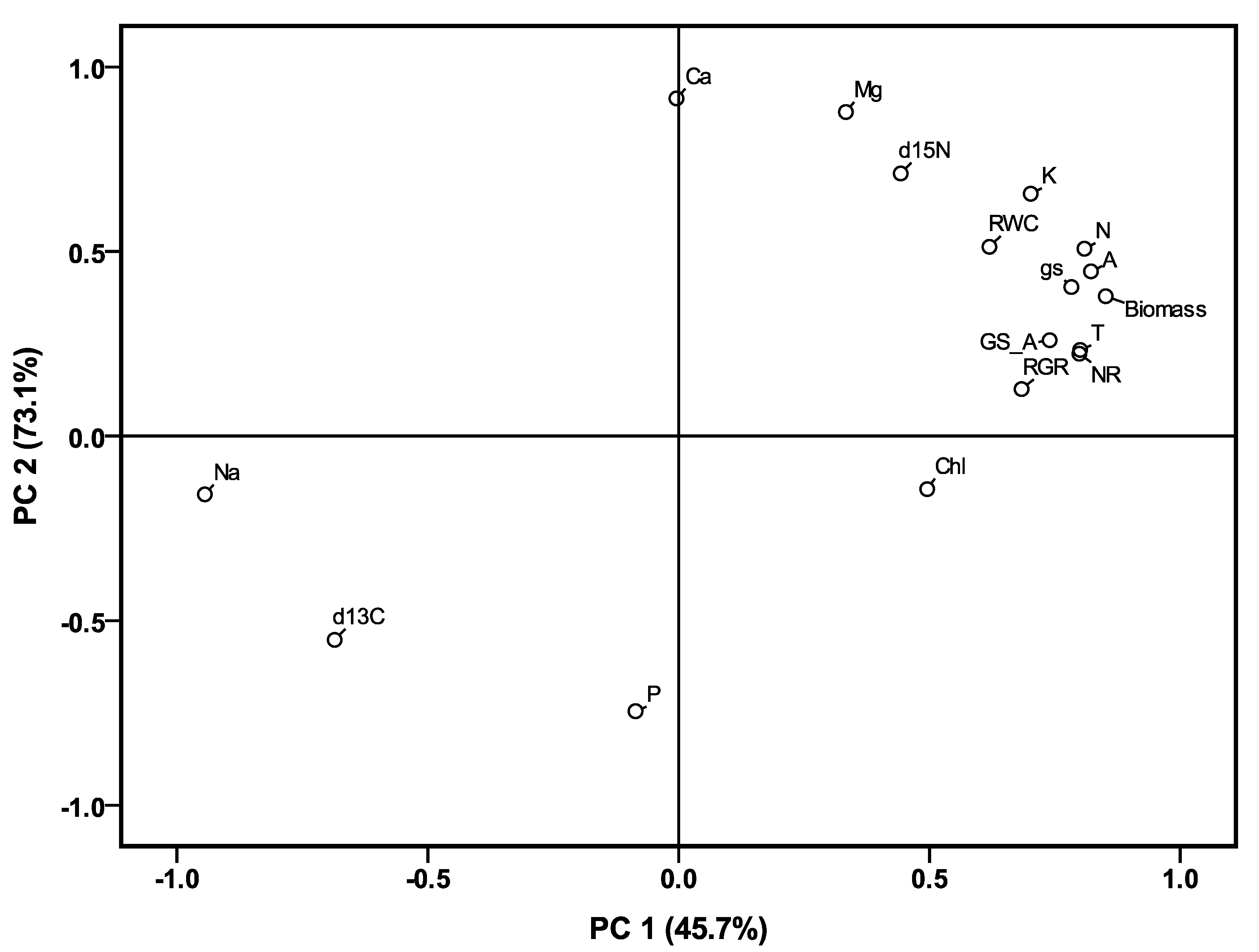
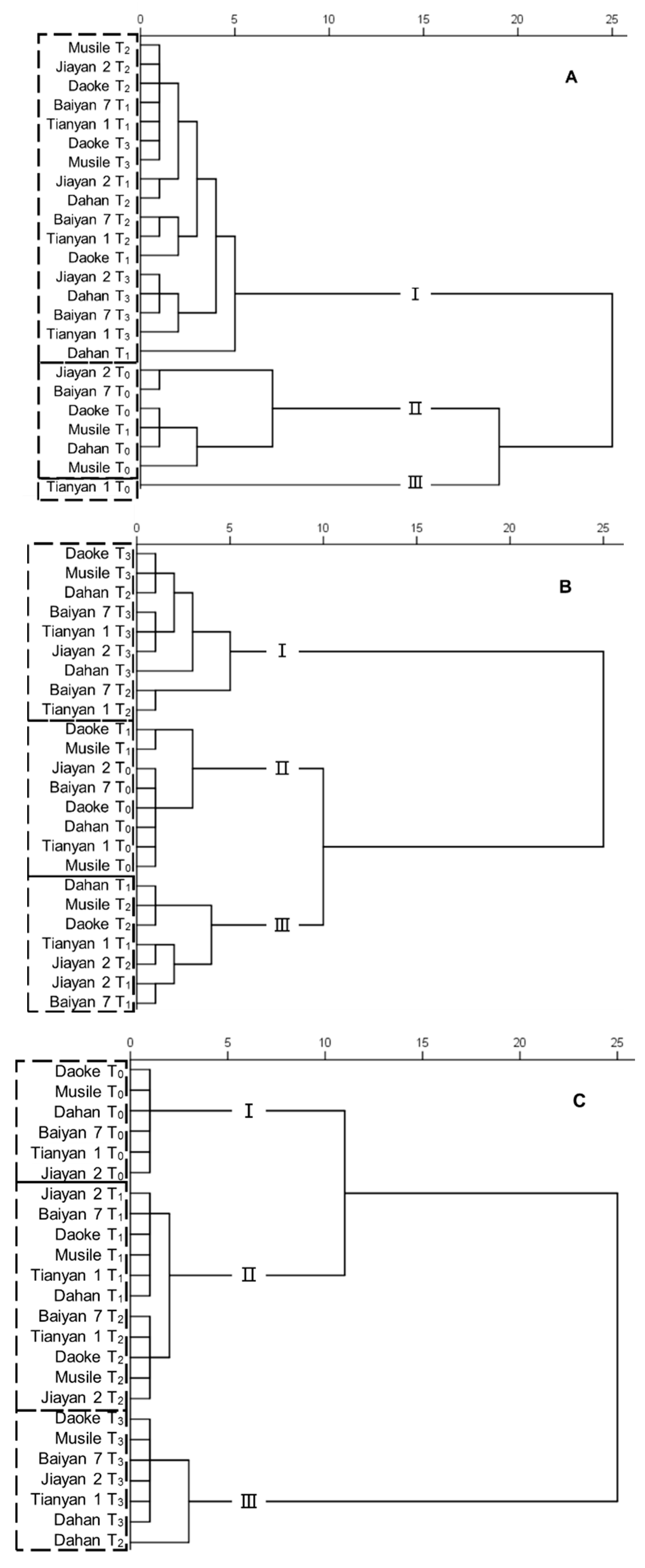
2.5. Overall Effects of Treatments and Genotypes on Agronomic and Physiological Traits
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Growth Parameters
4.3. Photosynthetic Gas Exchange Measurements
4.4. N Concentration and Stable Isotope Signatures in Total Organic Matter
4.5. NR and GS Enzyme Activity Determinations
4.6. Ion Concentration Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- MARA (Ministry of Agriculture and Rural Affairs of P.R.C.), 2018. Available online: http://www.moa.gov.cn/xw/bmdt/201801/t20180129_6135949.htm (accessed on 3 February 2020).
- Yang, C.; Wang, G.; Wang, M. Production and trade of wild oat forage in China. Pratacult. Sci. 2017, 5, 1129–1135. [Google Scholar]
- Chawade, A.; Lindén, P.; Bräutigam, M.; Jonsson, R.; Jonsson, A.; Moritz, T.; Olsson, O. Development of a model system to identify differences in spring and winter oat. PLoS ONE 2012, 7, e29792. [Google Scholar] [CrossRef] [PubMed]
- Nevo, E.; Chen, G. Drought and salt tolerances in wild relatives for wheat and barley improvement. Plant Cell Environ. 2010, 33, 670–685. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef]
- Chamekh, Z.; Ayadi, S.; Karmous, C.; Trifa, Y.; Amara, H.; Boudabbous, K.; Yousfi, S.; Serret, M.D.; Araus, J.L. Comparative effect of salinity on growth, grain yield, water use efficiency, δ13C and δ15N of landraces and improved durum wheat varieties. Plant Sci. 2016, 251, 44–53. [Google Scholar] [CrossRef]
- Yousfi, S.; Serret, M.D.; Márquez, A.J.; Voltas, J.; Araus, J.L. Combined use of δ13C, δ18O and δ15N tracks nitrogen metabolism and genotypic adaptation of durum wheat to salinity and water deficit. New Phytol. 2012, 194, 230–244. [Google Scholar] [CrossRef]
- Yousfi, S.; Serret, M.D.; Araus, J.L. Comparative response of δ13C, δ18O and δ15N in durum wheat exposed to salinity at the vegetative and reproductive stages. Plant Cell Environ. 2013, 36, 1214–1227. [Google Scholar] [CrossRef]
- Aranjuelo, I.; Cabrera-Bosquet, L.; Morcuende, R.; Avice, J.C.; Nogués, S.; Araus, J.L.; Martínez-Carrasco, R.; Pérez, P. Does ear C sink strength contribute to overcoming photosynthetic acclimation of wheat plants exposed to elevated CO2? J. Exp. Bot. 2011, 62, 3957–3969. [Google Scholar] [CrossRef]
- Zhou, B.; Serret, M.D.; Elazab, A.; Bort Pie, J.; Araus, J.L.; Aranjuelo, I.; Sanz-Sáez, Á. Wheat ear carbon assimilation and nitrogen remobilization contribute significantly to grain yield. J. Integr. Plant Biol. 2016, 58, 914–926. [Google Scholar] [CrossRef]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Parisi, D.; Verlotta, A.; Fuggi, A. Nitrogen metabolism in durum wheat under salinity: Accumulation of proline and glycine betaine. Funct. Plant Biol. 2008, 35, 412–426. [Google Scholar] [CrossRef]
- Botella, M.A.; Cruz, C.; Martins-Louçao, M.A.; Cerdá, A. Nitrate reductase activity in wheat seedlings as affected by NO3−/NH4+ ratio and salinity. J. Plant Physiol. 1993, 142, 531–536. [Google Scholar] [CrossRef]
- Irshad, M.; Honna, T.; Eneji, A.; Yamamoto, S. Wheat response to nitrogen source under saline conditions. J. Plant Nutr. 2002, 25, 2603–2612. [Google Scholar] [CrossRef]
- Debouba, M.; Maâroufi-Dghimi, H.; Suzuki, A.; Ghorbel, M.H.; Gouia, H. Changes in growth and activity of enzymes involved in nitrate reduction and ammonium assimilation in tomato seedlings in response to NaCl stress. Ann. Bot. 2007, 99, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Luque, E.; Chamizo-Ampudia, A.; Llamas, A.; Galvan, A.; Fernandez, E. Understanding nitrate assimilation and its regulation in microalgae. Front. Plant Sci. 2015, 6, 899. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, D.; Gill, H.S.; Pfluegl, G.M.; Rotstein, S.H. Structure–function relationships of glutamine synthetases. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 2000, 1477, 122–145. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Winter, K.; Turner, B.L. Plant δ15N correlates with the transpiration efficiency of nitrogen acquisition in tropical trees. Plant Physiol. 2009, 151, 1667–1676. [Google Scholar] [CrossRef]
- Craine, J.M.; Brookshire, E.; Cramer, M.D.; Hasselquist, N.J.; Koba, K.; Marin-Spiotta, E.; Wang, L. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil 2015, 396, 1–26. [Google Scholar] [CrossRef]
- Serret, M.; Ortiz-Monasterio, I.; Pardo, A.; Araus, J. The effects of urea fertilisation and genotype on yield, nitrogen use efficiency, δ15N and δ13C in wheat. Ann. Appl. Biol. 2008, 153, 243–257. [Google Scholar] [CrossRef]
- Evans, R.; Bloom, A.; Sukrapanna, S.; Ehleringer, J. Nitrogen isotope composition of tomato (Lycopersicon esculentum Mill. cv. T-5) grown under ammonium or nitrate nutrition. Plant Cell Environ. 1996, 19, 1317–1323. [Google Scholar] [CrossRef]
- Liu, T.; Wang, B.; Xiao, H.; Wang, R.; Yang, B.; Cao, Q.; Cao, Y. Differentially improved soil microenvironment and seedling growth of Amorpha fruticosa by plastic, sand and straw mulching in a saline wasteland in northwest China. Ecol. Eng. 2018, 122, 126–134. [Google Scholar] [CrossRef]
- Baltruschat, H.; Fodor, J.; Harrach, B.D.; Niemczyk, E.; Barna, B.; Gullner, G.; Janeczko, A.; Kogel, K.H.; Schäfer, P.; Schwarczinger, I. Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants. New Phytol. 2008, 180, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Yousfi, S.; Serret, M.D.; Voltas, J.; Araus, J.L. Effect of salinity and water stress during the reproductive stage on growth, ion concentrations, Δ13C, and δ15N of durum wheat and related amphiploids. J. Exp. Bot. 2010, 61, 3529–3542. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Ma, B.; Ren, C. Growth, gas exchange, chlorophyll fluorescence, and ion content of naked oat in response to salinity. Crop Sci. 2007, 47, 123–131. [Google Scholar] [CrossRef]
- Ruan, Y.; Hu, Y.; Schmidhalter, U. Insights on the role of tillering in salt tolerance of spring wheat from detillering. Environ. Exp. Bot. 2008, 64, 33–42. [Google Scholar] [CrossRef]
- Wu, H.; Shabala, L.; Barry, K.; Zhou, M.; Shabala, S. Ability of leaf mesophyll to retain potassium correlates with salinity tolerance in wheat and barley. Physiol. Plant. 2013, 149, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Zan, W.; Geng, Z.; Xue-min, W.; Hong-wen, G. Growth, ion content and photosynthetic responses of two Elytrigia Desv. species seedlings to salinity stress. Afr. J. Biotechnol. 2011, 10, 7390–7396. [Google Scholar]
- Sun, Z.; Ren, L.; Fan, J.; Li, Q.; Wang, K.; Guo, M.; Wang, L.; Li, J.; Zhang, G.; Yang, Z. Salt response of photosynthetic electron transport system in wheat cultivars with contrasting tolerance. Plant Soil Environ. 2016, 62, 515–521. [Google Scholar]
- Kiani-Pouya, A.; Rasouli, F.; Bazihizina, N.; Zhang, H.; Hedrich, R.; Shabala, S. A large-scale screening of quinoa accessions reveals an important role of epidermal bladder cells and stomatal patterning in salinity tolerance. Environ. Exp. Bot. 2019, 168, 103885. [Google Scholar] [CrossRef]
- Barbour, M.M.; Warren, C.R.; Farquhar, G.D.; Forrester, G.; Brown, H. Variability in mesophyll conductance between barley genotypes, and effects on transpiration efficiency and carbon isotope discrimination. Plant Cell Environ. 2010, 33, 1176–1185. [Google Scholar] [CrossRef]
- Rajabi, A.; Ober, E.S.; Griffiths, H. Genotypic variation for water use efficiency, carbon isotope discrimination, and potential surrogate measures in sugar beet. Field Crop. Res. 2009, 112, 172–181. [Google Scholar] [CrossRef]
- Rebetzke, G.; Condon, A.G.; Richards, R.; Farquhar, G. Selection for reduced carbon isotope discrimination increases aerial biomass and grain yield of rainfed bread wheat. Crop Sci. 2002, 42, 739–745. [Google Scholar] [CrossRef]
- Morgan, J.A.; LeCain, D.R.; McCaig, T.N.; Quick, J.S. Gas exchange, carbon isotope discrimination, and productivity in winter wheat. Crop Sci. 1993, 33, 178–186. [Google Scholar] [CrossRef]
- Agastian, P.; Kingsley, S.; Vivekanandan, M. Effect of salinity on photosynthesis and biochemical characteristics in mulberry genotypes. Photosynthetica 2000, 38, 287–290. [Google Scholar] [CrossRef]
- Elsheery, N.I.; Cao, K.-F. Gas exchange, chlorophyll fluorescence, and osmotic adjustment in two mango cultivars under drought stress. Acta Physiol. Plant. 2008, 30, 769–777. [Google Scholar] [CrossRef]
- Hirel, B.; Chardon, F.; Durand, J. The contribution of molecular physiology to the improvement of nitrogen use efficiency in crops. J. Crop Sci. Biotechnol. 2007, 10, 123–132. [Google Scholar]
- Katiyar, S.; Dubey, R. Influence of NaCl salinity on behaviours of nitrate reductase and nitrite reductase in rice seedlings differing in salt tolerance. J. Agron. Crop Sci. 1992, 169, 289–297. [Google Scholar] [CrossRef]
- Reda, M.; Migocka, M.; Kłobus, G. Effect of short-term salinity on the nitrate reductase activity in cucumber roots. Plant Sci. 2011, 180, 783–788. [Google Scholar] [CrossRef]
- Botella, M.A.; Martínez, V.; Nieves, M.; Cerdá, A. Effect of salinity on the growth and nitrogen uptake by wheat seedlings. J. Plant Nutr. 1997, 20, 793–804. [Google Scholar] [CrossRef]
- Ullrich, W.R. Salinity and nitrogen nutrition. In Salinity: Environment-Plants-Molecules; Springer, Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 229–248. [Google Scholar]
- Parry, M.A.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.-G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.V.; Berry, J. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- Zhou, B.; Serret, M.D.; Pie, J.B.; Shah, S.S.; Li, Z. Relative contribution of nitrogen absorption, remobilization, and partitioning to the ear during grain filling in chinese winter wheat. Front. Plant Sci. 2018, 9, 1351. [Google Scholar] [CrossRef] [PubMed]
- Yousfi, S.; Serret, M.D.; Araus, J.L. Shoot δ15N gives a better indication than ion concentration or Δ13C of genotypic differences in the response of durum wheat to salinity. Funct. Plant Biol. 2009, 36, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D. δ15N as an integrator of the nitrogen cycle. Trends Ecol. Evol. 2001, 16, 153–162. [Google Scholar] [CrossRef]
- Zhou, B.; Sanz-Sáez, Á.; Elazab, A.; Shen, T.; Sánchez-Bragado, R.; Bort, J.; Serret, M.D.; Araus, J.L. Physiological traits contributed to the recent increase in yield potential of winter wheat from Henan Province, China. J. Integr. Plant Biol. 2014, 56, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Liu, C. Study on Agronomic Traits and Cropping Patterns of Forage Oats in Western Jilin Province; Northeast Normal University: Changchun, China, 2018. (In Chinese) [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 1–32. [Google Scholar]
- Al-Tamimi, N.; Brien, C.; Oakey, H.; Berger, B.; Saade, S.; Ho, Y.S.; Schmöckel, S.M.; Tester, M.; Negrão, S. Salinity tolerance loci revealed in rice using high-throughput non-invasive phenotyping. Nat. Commun. 2016, 7, 13342. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.; Rebetzke, G.; Farquhar, G. Improving intrinsic water-use efficiency and crop yield. Crop Sci. 2002, 42, 122–131. [Google Scholar]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Biomass (g DW Plant−1) | Plant Height (cm) | Tiller Number (Per Plant) | Culm Thickness (mm) | RGR (Per Day) | |
|---|---|---|---|---|---|
| Genotypes (G) | |||||
| Dahan | 2.54 a | 60.11 abc | 2.83 a | 0.47 a | 0.04 ab |
| Musile | 2.55 a | 56.98 a | 3.17 a | 0.54 b | 0.07 c |
| Tianyan 1 | 2.79 b | 65.89 d | 2.83 a | 0.52 b | 0.09 d |
| Baiyan 7 | 2.85 b | 61.81 cd | 2.67 a | 0.52 b | 0.07 c |
| Daoke | 2.81 b | 58.30 bc | 3.25 a | 0.55 b | 0.06 bc |
| Jiayan 2 | 2.80 b | 58.30 ab | 3.00 a | 0.53 b | 0.04 a |
| Treatments (T) | |||||
| T0 (Control) | 3.89 d | 71.35 d | 3.61 c | 0.58 c | 0.08 c |
| T1 (8 dS m−1) | 2.90 c | 64.96 c | 3.17 bc | 0.53 b | 0.07 c |
| T2 (16 dS m−1) | 2.36 b | 57.90 b | 2.94 b | 0.50 a | 0.06 b |
| T3 (24 dS m−1) | 1.73 a | 49.66 a | 2.11 a | 0.48 a | 0.03 a |
| Level of Significance | |||||
| G | 0.000 | 0.000 | 0.152 | 0.000 | 0.000 |
| T | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| G × T | 0.000 | 0.000 | 0.048 | 0.075 | 0.928 |
| Genotypes | Biomass (g) | Biomass Relative to Control (%) | ||
|---|---|---|---|---|
| Control | T1 | T2 | T3 | |
| Dahan | 3.46 a | 82.24 b | 62.12 ab | 49.78 a |
| Musile | 3.63 ab | 76.52 b | 62.72 ab | 42.54 a |
| Tianyan 1 | 3.73 ab | 79.64 b | 74.79 b | 44.10 a |
| Baiyan 7 | 3.92 cd | 78.66 b | 61.20 ab | 40.20 a |
| Daoke | 4.22 cd | 67.86 a | 58.50 ab | 50.94 a |
| Jiayan 2 | 4.38 d | 66.26 a | 48.23 a | 43.70 a |
| Sum of square (G) | 1.80 *** | 650.13 *** | 1089.22 * | 268.4 ns |
| RWC (#) | Pn (μmol CO2 m−2 s−1) | Gs (mmol H2O m−2 s−1) | E (mmol H2O m−2 s−1) | WUEi (μmol CO2 mmol H2O−1) | Ci/Ca (µmol mol−1) | δ13Cshoot (‰) | δ13Croot (‰) | |
|---|---|---|---|---|---|---|---|---|
| Genotypes(G) | ||||||||
| Dahan | 0.81 a | 7.67 a | 90.50 a | 2.43 a | 3.19 b | 0.57 a | −26.72 b | −26.76 bc d |
| Musile | 0.88 a | 8.52 bc | 144.08 c | 3.67 b | 2.27 a | 0.61 a | −26.84 b | −27.15 a |
| Tianyan 1 | 0.90 a | 8.78 c | 158.67 d | 3.63 b | 2.27 a | 0.62 a | −27.30 a | −26.96 a bc |
| Baiyan 7 | 0.89 a | 8.37 bc | 125.58 b | 3.13 ab | 2.54 a | 0.57 a | −26.63 b | −26.65 c d |
| Daoke | 0.92 a | 9.79 d | 122.00 b | 3.38 b | 2.72 ab | 0.61 a | −26.81 b | −27.08 ab |
| Jiayan 2 | 0.88 a | 7.93 ab | 129.33 b | 3.11 ab | 2.24 a | 0.60 a | −26.35 b | −26.53 d |
| Treatments(T) | ||||||||
| T0 (Control) | 1.40 b | 17.22 d | 253.3 c | 5.33 c | 3.37 b | 0.65 b | −28.03 a | −27.13 a |
| T1 (8 dS m−1) | 0.76 a | 8.68 c | 100.8 b | 3.16 b | 3.00 b | 0.61 b | −26.95 b | −26.72 bc |
| T2 (16 dS m−1) | 0.71 a | 5.39 b | 98.4 b | 2.82 b | 1.96 a | 0.56 a | −26.08 c | −26.63 c |
| T3 (24 dS m−1) | 0.66 a | 2.72 a | 60.8 a | 1.57 a | 1.82 a | 0.56 a | −26.04 c | −26.95 ab |
| Level of Significance | ||||||||
| G | 0.875 | 0.000 | 0.000 | 0.001 | 0.001 | 0.473 | 0.000 | 0.000 |
| T | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.004 | 0.000 | 0.000 |
| G × T | 0.991 | 0.000 | 0.000 | 0.000 | 0.099 | 0.000 | 0.081 | 0.665 |
| δ15Nshoot (‰) | δ15Nroot (‰) | Nshoot (%) | Nroot (%) | Chl (SPAD Units) | NR Activity (μmol NO2− g−1 FW h−1) | GS Activity (OD g−1 FW h−1) | |
|---|---|---|---|---|---|---|---|
| Genotypes (G) | |||||||
| Dahan | −3.03 a | −4.42 a | 3.66 a | 2.04 ab | 43.15 a | 4.09 bc | 5.02 a |
| Musile | −3.32 a | −3.43 b | 3.84 ab | 2.00 ab | 50.60 b | 4.41 c | 5.11 a |
| Tianyan 1 | −2.80 a | −4.49 a | 4.01 b | 1.94 ab | 44.83 a | 2.78 a | 5.09 a |
| Baiyan 7 | −3.01 a | −3.74 ab | 3.74 a | 1.79 a | 48.16 b | 3.78 b | 4.94 a |
| Daoke | −2.97 a | −4.57 a | 3.77 a | 2.15 b | 49.08 b | 4.94 d | 4.42 a |
| Jiayan 2 | −3.14 a | −3.61 ab | 3.73 a | 1.86 ab | 43.96 a | 4.32 ab | 4.55 a |
| Treatments (T) | |||||||
| T0 (Control) | −2.04 c | −3.05 b | 4.62 d | 2.16 b | 48.18 b | 6.42 d | 6.16 d |
| T1 (8 dS m−1) | −2.95 b | −4.01 a | 3.80 c | 1.97 ab | 46.76 b | 4.80 c | 5.45 c |
| T2 (16 dS m−1) | −3.63 a | −4.54 a | 3.53 b | 1.79 a | 47.82 b | 3.76 b | 4.50 b |
| T3 (24 dS m−1) | −3.59 a | −4.59 a | 3.22 a | 1.93 a | 43.76 a | 1.22 a | 3.31 a |
| Level of Significance | |||||||
| G | 0.544 | 0.002 | 0.001 | 0.024 | 0.000 | 0.000 | 0.027 |
| T | 0.000 | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 0.000 |
| G × T | 0.244 | 0.169 | 0.460 | 0.563 | 0.002 | 0.000 | 0.006 |
| Shoot | Root | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ca2+ | Mg2+ | K+ | Na+ | P | K+/Na+ | Ca2+/Na+ | Ca2+ | Mg2+ | K+ | Na+ | P | K+/Na+ | Ca2+/Na+ | |
| Genotypes (G) | ||||||||||||||
| Dahan | 0.080 b | 0.074 ab | 1.253 a | 1.951 b | 0.145 a | 15.66 a | 0.924 a | 0.207 a | 0.071 a | 0.411 abc | 0.591 a | 0.148 a | 1.754 a | 0.93 a |
| Musile | 0.076 ab | 0.068 a | 1.246 a | 1.666 ab | 0.148 a | 24.53 a | 1.461 a | 0.223 a | 0.080 a | 0.486 bc | 0.721 a | 0.173 ab | 1.953 a | 1.02 a |
| Tianyan 1 | 0.080 b | 0.080 b | 1.317 a | 1.668 ab | 0.171 b | 20.11 a | 1.152 a | 0.234 a | 0.076 a | 0.398 ab | 0.649 a | 0.184 b | 2.022 a | 1.44 ab |
| Jiayan 2 | 0.063 a | 0.065 a | 1.274 a | 1.716 ab | 0.167 b | 41.50 b | 1.871 a | 0.254 a | 0.076 a | 0.378 a | 0.603 a | 0.171 ab | 2.072 a | 1.60 ab |
| Daoke | 0.072 ab | 0.068 a | 1.242 a | 1.492 a | 0.148 a | 16.20 a | 0.918 a | 0.213 a | 0.075 a | 0.496 c | 0.714 a | 0.163 ab | 1.916 a | 0.97 a |
| Baiyan 7 | 0.065 a | 0.066 a | 1.252 a | 1.544 a | 0.159 ab | 22.28 a | 1.020 a | 0.263 a | 0.075 a | 0.379 a | 0.551 a | 0.160 ab | 2.243 a | 1.93 b |
| Treatments (T) | ||||||||||||||
| T0 (Control) | 0.092 c | 0.088 b | 1.714 c | 0.023 a | 0.130 a | 91.564 b | 4.791 b | 0.452 c | 0.092d | 0.692 c | 0.107 a | 0.155 a | 6.700 b | 4.632 b |
| T1 (8 dS m−1) | 0.061 a | 0.066 a | 1.262 b | 1.178 b | 0.156 b | 1.076 a | 0.052 a | 0.184 b | 0.080 c | 0.415 b | 0.743 b | 0.152 a | 0.561 a | 0.259 a |
| T2 (16 dS m−1) | 0.062 a | 0.062 a | 1.081 a | 1.925 c | 0.185 c | 0.599 a | 0.032 a | 0.134 a | 0.069 b | 0.298 a | 0.747 b | 0.169 a | 0.402 a | 0.186 a |
| T3 (24 dS m−1) | 0.076 b | 0.065 a | 1.000 a | 3.565d | 0.154 b | 0.283 a | 0.021 a | 0.158 ab | 0.060 a | 0.295 a | 0.956 c | 0.190 b | 0.311 a | 0.177 a |
| Level of Significance | ||||||||||||||
| G | 0.002 | 0.000 | 0.650 | 0.024 | 0.000 | 0.000 | 0.031 | 0.104 | 0.139 | 0.001 | 0.075 | 0.008 | 0.754 | 0.011 |
| T | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| G × T | 0.128 | 0.120 | 0.428 | 0.156 | 0.012 | 0.000 | 0.004 | 0.216 | 0.023 | 0.359 | 0.683 | 0.238 | 0.933 | 0.003 |
| Models | Final Stepwise | R2 |
|---|---|---|
| Water status traits † | Biomass = 7.41 + 0.13 Pn + 0.17 E − 0.31 δ13CShoot + 0.47 δ13Croot | 0.82 * |
| N status traits ‡ | Biomass = −1.38 + 0.81 Nshoot + 0.09 NR + 0.14 GSactivity | 0.79 * |
| Water and N traits combined § | Biomass = 7.42 + 0.59 Nshoot + 0.54 Pn + 0.16 GSactivity + 0.31 δ13Croot | 0.83 ** |
| Total parameters # | Biomass = 9.91 − 0.42 Na+shoot + 9.37 Mg2+shoot − 0.47 Na+root + 0.23 δ13Croot − 3.67 Pshoot | 0.90 *** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadaqat Shah, S.; Li, Z.; Yan, H.; Shi, L.; Zhou, B. Comparative Study of the Effects of Salinity on Growth, Gas Exchange, N Accumulation and Stable Isotope Signatures of Forage Oat (Avena sativa L.) Genotypes. Plants 2020, 9, 1025. https://doi.org/10.3390/plants9081025
Sadaqat Shah S, Li Z, Yan H, Shi L, Zhou B. Comparative Study of the Effects of Salinity on Growth, Gas Exchange, N Accumulation and Stable Isotope Signatures of Forage Oat (Avena sativa L.) Genotypes. Plants. 2020; 9(8):1025. https://doi.org/10.3390/plants9081025
Chicago/Turabian StyleSadaqat Shah, Syed, Zhijian Li, Hong Yan, Lianxuan Shi, and Bangwei Zhou. 2020. "Comparative Study of the Effects of Salinity on Growth, Gas Exchange, N Accumulation and Stable Isotope Signatures of Forage Oat (Avena sativa L.) Genotypes" Plants 9, no. 8: 1025. https://doi.org/10.3390/plants9081025
APA StyleSadaqat Shah, S., Li, Z., Yan, H., Shi, L., & Zhou, B. (2020). Comparative Study of the Effects of Salinity on Growth, Gas Exchange, N Accumulation and Stable Isotope Signatures of Forage Oat (Avena sativa L.) Genotypes. Plants, 9(8), 1025. https://doi.org/10.3390/plants9081025