Semi-Targeted Metabolomics to Validate Biomarkers of Grape Downy Mildew Infection Under Field Conditions

 ,
,
Abstract
1. Introduction
2. Results and Discussion
2.1. Metabolomics Profiling of Downy Mildew Infected Grape Leaves
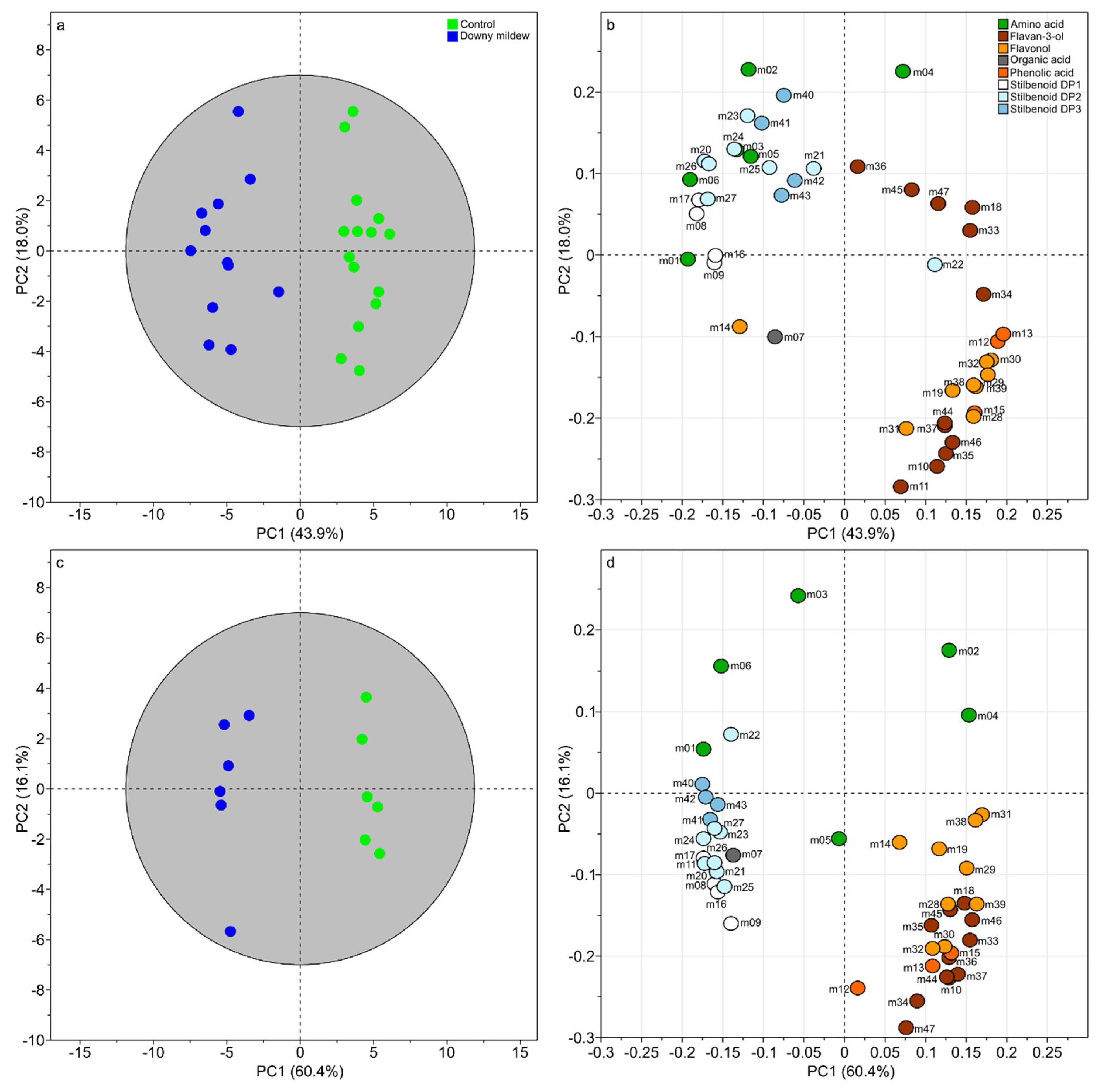
2.2. Unsupervised Multivariate Statistics for a Brief Overview of Metabolic Changes Upon Natural Downy Mildew Infection Under Field Condtions
2.3. Supervised Multivariate Statistics with “Infection Status” as Discriminant Variable
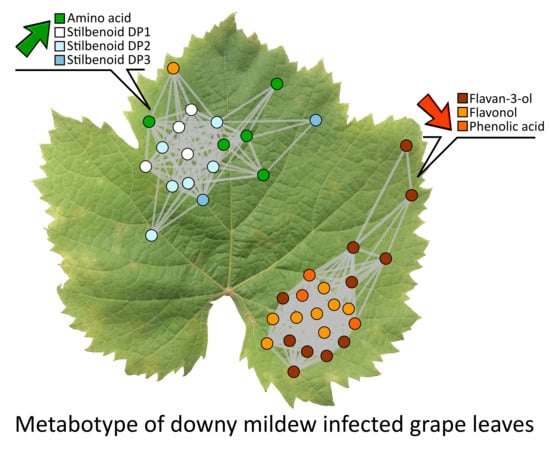
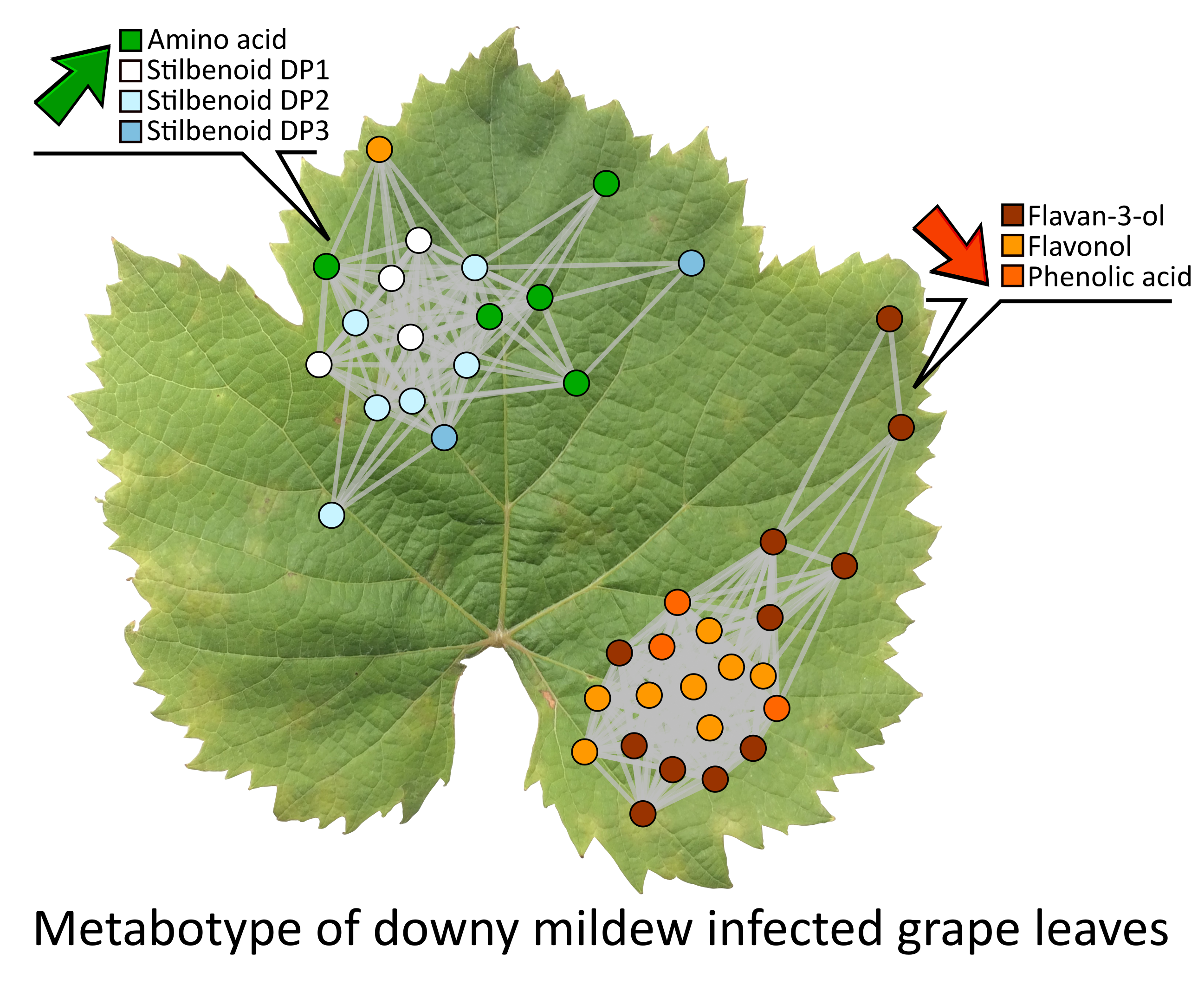
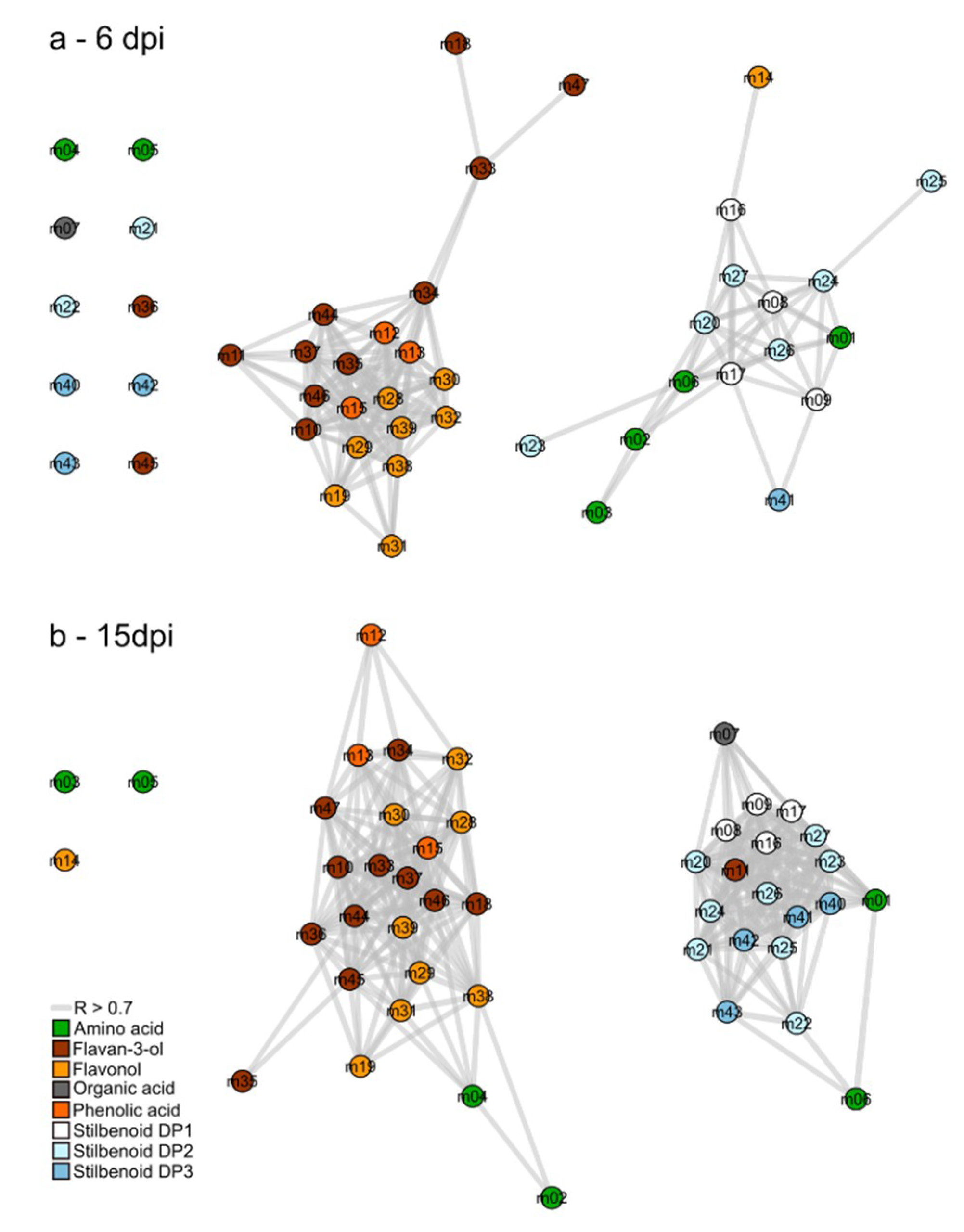
2.4. Metabolic Network Analysis of Grape Leaves Upon Downy Mildew
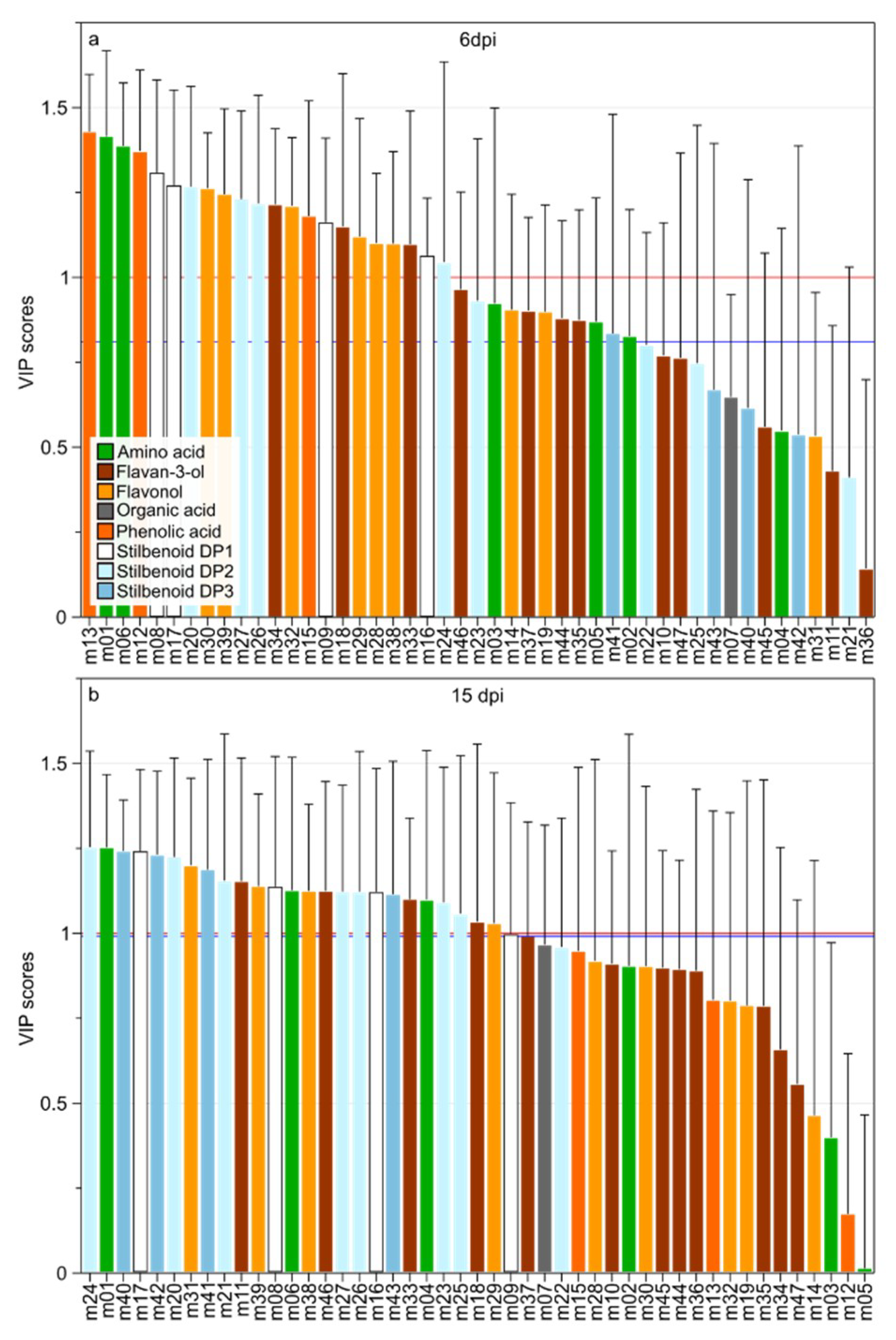
2.5. Variable Selection for the Identification of the Infection Biomarkers
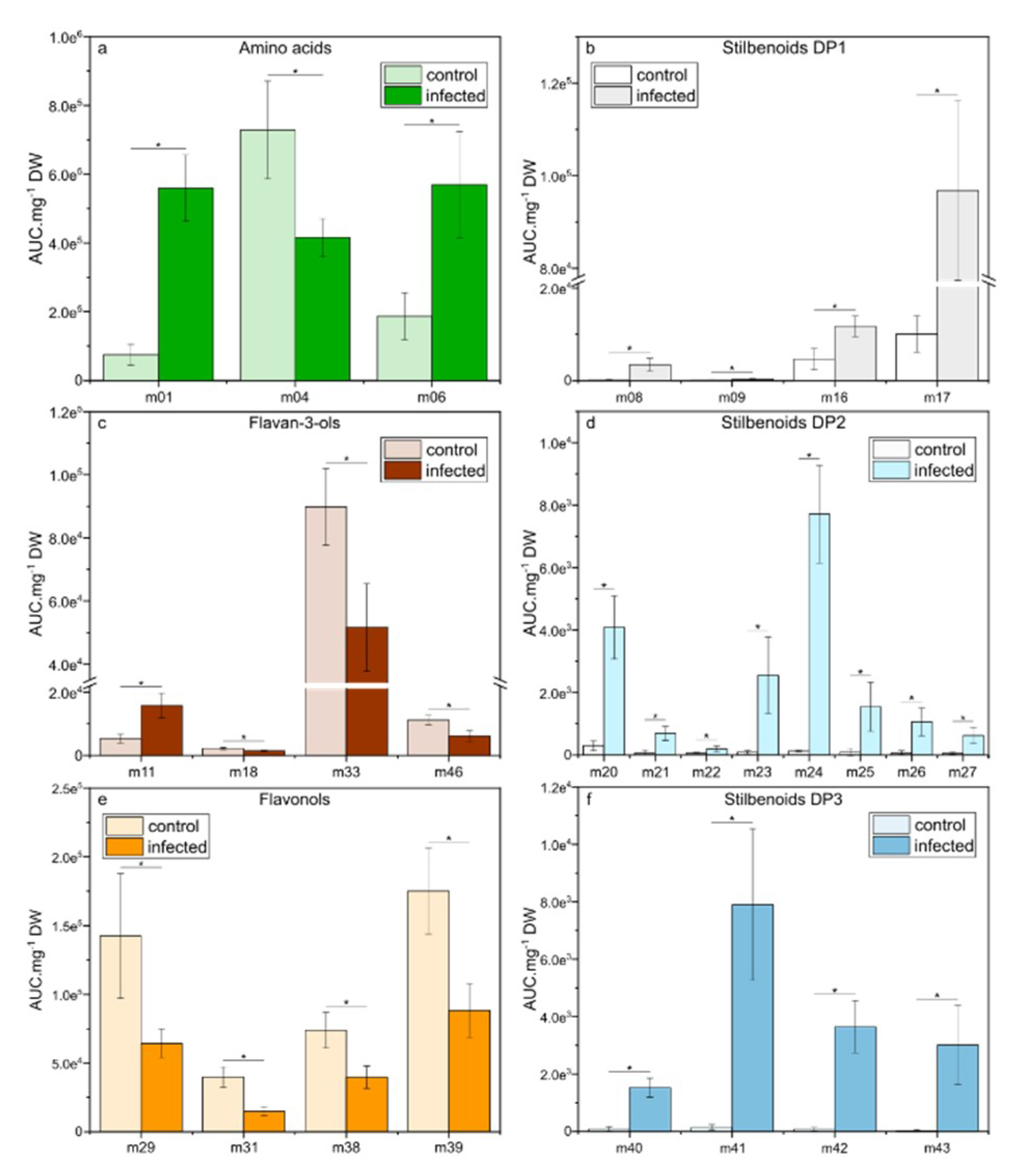
2.6. Changes in Relative Concentration of the Selected Metabolites in Grape Leaves Upon Downy Mildew Infection
3. Discussion
4. Materials and Methods
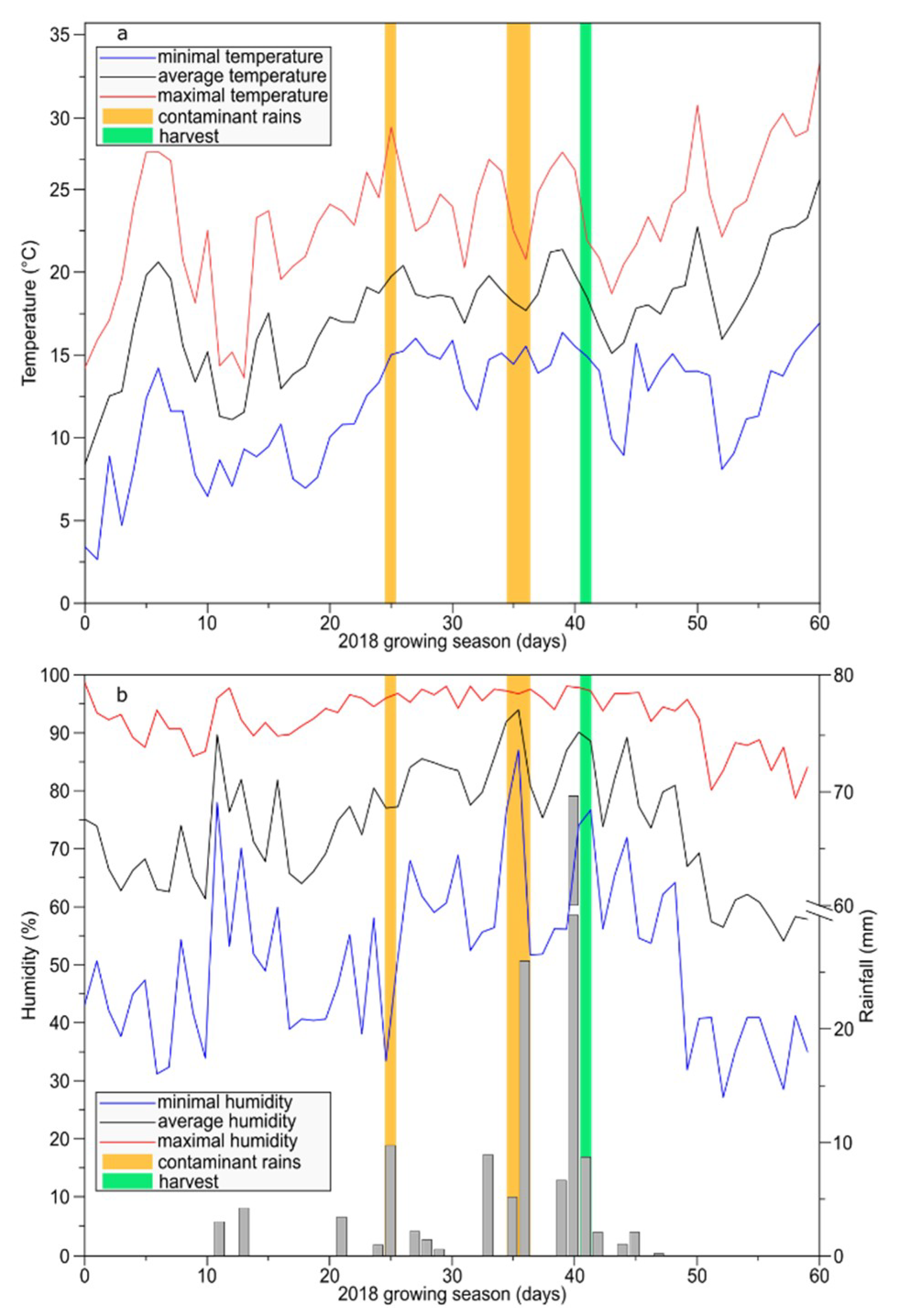
4.1. Plant Material and Growth Conditions
4.2. Chemicals
4.3. UPLC-MS Analyses
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar] [CrossRef]
- Runkle, J.; Flocks, J.; Economos, J.; Dunlop, A.L. A systematic review of Mancozeb as a reproductive and developmental hazard. Environ. Int. 2017, 99, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Juang, K.W.; Lee, Y.I.; Lai, H.Y.; Wang, C.H.; Chen, B.C. Copper accumulation, translocation, and toxic effects in grapevine cuttings. Environ. Sci. Pollut. Res. 2012, 19, 1315–1322. [Google Scholar] [CrossRef]
- Komárek, M.; Čadková, E.; Chrastný, V.; Bordas, F.; Bollinger, J.C. Contamination of vineyard soils with fungicides: A review of environmental and toxicological aspects. Environ. Int. 2010, 36, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Dagostin, S.; Schärer, H.J.; Pertot, I.; Tamm, L. Are there alternatives to copper for controlling grapevine downy mildew in organic viticulture? Crop Prot. 2011, 30, 776–788. [Google Scholar] [CrossRef]
- Jacometti, M.A.; Wratten, S.D.; Walter, M. Review: Alternatives to synthetic fungicides for Botrytis cinerea management in vineyards. Aust. J. Grape Wine Res. 2010, 16, 154–172. [Google Scholar] [CrossRef]
- Watt, M.; Fiorani, F.; Usadel, B.; Rascher, U.; Muller, O.; Schurr, U. Phenotyping: New windows into the plant for breeders. Annu. Rev. Plant Biol. 2020, 71, 689–712. [Google Scholar] [CrossRef]
- Kennelly, M.M.; Gadoury, D.M.; Wilcox, W.F.; Magarey, P.A.; Seem, R.C. Primary infection, lesion productivity, and survival of sporangia in the grapevine downy mildew pathogen Plasmopara viticola. Phytopathology 2007, 97, 512–522. [Google Scholar] [CrossRef]
- Polesani, M.; Desario, F.; Ferrarini, A.; Zamboni, A.; Pezzotti, M.; Kortekamp, A.; Polverari, A. cDNA-AFLP analysis of plant and pathogen genes expressed in grapevine infected with Plasmopara viticola. Bmc Genom. 2008, 9, 1–14. [Google Scholar] [CrossRef]
- Gamm, M.; Héloir, M.C.; Bligny, R.; Vaillant-Gaveau, N.; Trouvelot, S.; Alcaraz, G.; Frettinger, P.; Clément, C.; Pugin, A.; Wendehenne, D.; et al. Changes in carbohydrate metabolism in Plasmopara viticola-infected grapevine leaves. Mol. Plant-Microbe Interact. 2011, 24, 1061–1073. [Google Scholar] [CrossRef]
- Mattivi, F.; Vrhovsek, U.; Malacarne, G.; Masuero, D.; Zulini, L.; Stefanini, M.; Moser, C.; Velasco, R.; Guella, G. Profiling of resveratrol oligomers, important stress metabolites, accumulating in the leaves of hybrid Vitis vinifera (Merzling × Teroldego) genotypes infected with Plasmopara viticola. J. Agric. Food Chem. 2011, 59, 5364–5375. [Google Scholar] [CrossRef] [PubMed]
- Chitarrini, G.; Soini, E.; Riccadonna, S.; Franceschi, P.; Zulini, L.; Masuero, D.; Vecchione, A.; Stefanini, M.; Di Gaspero, G.; Mattivi, F.; et al. Identification of biomarkers for defense response to Plasmopara viticola in a resistant grape variety. Front. Plant Sci. 2017, 8, 1524. [Google Scholar] [CrossRef] [PubMed]
- Pezet, R.; Gindro, K.; Viret, O.; Spring, J.L. Glycosylation and oxidative dimerization of resveratrol are respectively associated to sensitivity and resistance of grapevine cultivars to downy mildew. Physiol. Mol. Plant Pathol. 2004, 65, 297–303. [Google Scholar] [CrossRef]
- Pezet, R.; Perret, C.; Jean-Denis, J.B.; Tabacchi, R.; Gindro, K.; Viret, O. δ-viniferin, a resveratrol dehydrodimer: One of the major stilbenes synthesized by stressed grapevine leaves. J. Agric. Food Chem. 2003, 51, 5488–5492. [Google Scholar] [CrossRef] [PubMed]
- Dai, G.H.; Andary, C.; Mondolot-Cosson, L.; Boubals, D. Histochemical responses of leaves of in vitro plantlets of Vitis spp. to infection with Plasmopara viticola. Phytopathology 1995, 85, 149–154. [Google Scholar] [CrossRef]
- Gindro, K.; Spring, J.L.; Pezet, R.; Richter, H.; Viret, O. Histological and biochemical criteria for objective and early selection of grapevine cultivars resistant to Plasmopara viticola. Vitis–J. Grapevine Res. 2006, 45, 191–196. [Google Scholar]
- Pawlus, A.D.; Waffo-Téguo, P.; Shaver, J.; Mérillon, J.M. Stilbenoid chemistry from wine and the genus Vitis, a review. OENO One 2012, 46, 57. [Google Scholar] [CrossRef]
- Rivière, C.; Pawlus, A.D.; Mérillon, J.M. Natural stilbenoids: Distribution in the plant kingdom and chemotaxonomic interest in Vitaceae. Nat. Prod. Rep. 2012, 29, 1317. [Google Scholar] [CrossRef]
- Pezet, R.; Gindro, K.; Viret, O.; Richter, H. Effects of resveratrol, viniferins and pterostilbene on Plasmopara viticola zoospore mobility and disease development. Vitis–J. Grapevine Res. 2004, 43, 145–148. [Google Scholar]
- Schnee, S.; Queiroz, E.F.; Voinesco, F.; Marcourt, L.; Dubuis, P.; Wolfender, J.; Gindro, K. Vitis vinifera canes, a new source of antifungal compounds against Plasmopara viticola, Erysiphe necator and Botrytis cinerea. J. Agric. Food Chem. 2013, 61, 5459–5467. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics–The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yin, Y. Strategies for large-scale targeted metabolomics quantification by Liquid Chromatography-Mass Spectrometry. Analyst 2016, 141, 6362–6373. [Google Scholar] [CrossRef] [PubMed]
- Krauss, M.; Singer, H.; Hollender, J. LC-high resolution MS in environmental analysis: From target screening to the identification of unknowns. Anal. Bioanal. Chem. 2010, 397, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Flamini, R.; De Rosso, M.; De Marchi, F.; Dalla Vedova, A.; Panighel, A.; Gardiman, M.; Maoz, I.; Bavaresco, L. An innovative approach to grape metabolomics: Stilbene profiling by suspect screening analysis. Metabolomics 2013, 9, 1243–1253. [Google Scholar] [CrossRef]
- Billet, K.; Houillé, B.; De Bernonville, T.D.; Besseau, S.; Oudin, A.; Courdavault, V.; Delanoue, G.; Guérin, L.; Clastre, M.; Giglioli-Guivarc’h, N.; et al. Field-based metabolomics of Vitis vinifera L. stems provides new insights for genotype discrimination and polyphenol metabolism structuring. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- De Rosso, M.; Mayr, C.M.; Girardi, G.; Vedova, A.D.; Flamini, R. High-resolution mass spectrometry metabolomics of grape chemical markers to reveal use of not-allowed varieties in the production of Amarone and Recioto wines. Metabolomics 2018, 14, 124. [Google Scholar] [CrossRef]
- Anesi, A.; Stocchero, M.; Dal Santo, S.; Commisso, M.; Zenoni, S.; Ceoldo, S.; Tornielli, G.B.; Siebert, T.E.; Herderich, M.; Pezzotti, M.; et al. Towards a scientific interpretation of the terroir concept: Plasticity of the grape berry metabolome. Bmc Plant Biol. 2015, 15, 191. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Moss, R.; Mao, Q.; Taylor, D.; Saucier, C. Investigation of monomeric and oligomeric wine stilbenoids in red wines by ultra-high-performance liquid chromatography/electrospray ionization quadrupole time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2013, 27, 1815–1827. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Masuero, D.; Gasperotti, M.; Franceschi, P.; Caputi, L.; Viola, R.; Mattivi, F. A versatile targeted metabolomics method for the rapid quantification of multiple classes of phenolics in fruits and beverages. J. Agric. Food Chem. 2012, 60, 8831–8840. [Google Scholar] [CrossRef]
- Püssa, T.; Floren, J.; Kuldkepp, P.; Raal, A. Survey of grapevine Vitis vinifera stem polyphenols by Liquid Chromatography-Diode Array Detection-Tandem Mass Spectrometry. J. Agric. Food Chem. 2006, 54, 7488–7494. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo-González, M.; Martínez-Carballo, E.; Cancho-Grande, B.; Santiago, J.L.; Martínez, M.C.; Simal-Gándara, J. Pattern recognition of three Vitis vinifera L. red grapes varieties based on anthocyanin and flavonol profiles, with correlations between their biosynthesis pathways. Food Chem. 2012, 130, 9–19. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Arapitsas, P.; Stefanini, M.; Flick, G.; Mattivi, F. Analysis of the phenolic composition of fungus-resistant grape varieties cultivated in Italy and Germany using UHPLC-MS/MS. J. Mass Spectrom. 2014, 49, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Garrido, I.; Bartolomé, B.; Gómez-Cordovés, C. Chemical characterization of commercial dietary ingredients from Vitis vinifera L. Anal. Chim. Acta 2006, 563, 401–410. [Google Scholar] [CrossRef]
- Teixeira, N.; Azevedo, J.; Mateus, N.; De Freitas, V. Proanthocyanidin screening by LC–ESI-MS of Portuguese red wines made with teinturier grapes. Food Chem. 2016, 190, 300–307. [Google Scholar] [CrossRef]
- Franceschi, P.; Masuero, D.; Vrhovsek, U.; Mattivi, F.; Wehrens, R. A benchmark spike-in data set for biomarker identification in metabolomics. J. Chemom. 2012, 26, 16–24. [Google Scholar] [CrossRef]
- Malacarne, G.; Vrhovsek, U.; Zulini, L.; Masuero, D.; Cestaro, A.; Stefanini, M.; Delledonne, M.; Velasco, R.; Guella, G.; Mattivi, F.; et al. Resistance to Plasmopara viticola is associated with a complex pattern of stilbenoids and with specific host transcriptional responses. Bmc Plant Biol. 2011, 11, 1–13. [Google Scholar] [CrossRef]
- Vezzulli, S.; Malacarne, G.; Masuero, D.; Vecchione, A.; Dolzani, C.; Goremykin, V.; Mehari, Z.H.; Banchi, E.; Velasco, R.; Stefanini, M.; et al. The Rpv3-3 haplotype and stilbenoid induction mediate downy mildew resistance in a grapevine interspecific population. Front. Plant Sci. 2019, 10, 1–23. [Google Scholar] [CrossRef]
- Gabaston, J.; Cantos-Villar, E.; Biais, B.; Waffo-Teguo, P.; Renouf, E.; Corio-Costet, M.F.; Richard, T.; Mérillon, J.M. Stilbenes from Vitis vinifera L. waste: A sustainable tool for controlling Plasmopara viticola. J. Agric. Food Chem. 2017, 65, 2711–2718. [Google Scholar] [CrossRef]
- Grenville-Briggs, L.J.; West, P.V. The biotrophic stages of oomycete–plant interactions. In Advances in Applied Microbiology; Laskin, A.L., Bennett, J.W., Gadd, G.M., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005; Volume 57, pp. 217–243. ISBN 0065-2164. [Google Scholar]
- Ciaffi, M.; Paolacci, A.R.; Paolocci, M.; Alicandri, E.; Bigini, V.; Badiani, M.; Muganu, M. Transcriptional regulation of stilbene synthases in grapevine germplasm differentially susceptible to downy mildew. Bmc Plant Biol. 2019, 19, 1–18. [Google Scholar] [CrossRef]
- Maes, W.H.; Steppe, K. Perspectives for remote sensing with Unmanned Aerial Vehicles in precision agriculture. Trends Plant Sci. 2019, 24, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Latouche, G.; Bellow, S.; Poutaraud, A.; Meyer, S.; Cerovic, Z.G. Influence of constitutive phenolic compounds on the response of grapevine (Vitis vinifera L.) leaves to infection by Plasmopara viticola. Planta 2013, 237, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Bellow, S.; Latouche, G.; Brown, S.C.; Poutaraud, A.; Cerovic, Z.G. In vivo localization at the cellular level of stilbene fluorescence induced by Plasmopara viticola in grapevine leaves. J. Exp. Bot. 2012, 63, 3697–3708. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Novales, J.; Garde-Cerdán, T.; Tardáguila, J.; Gutiérrez-Gamboa, G.; Pérez-Álvarez, E.P.; Diago, M.P. Assessment of amino acids and total soluble solids in intact grape berries using contactless Vis and NIR spectroscopy during ripening. Talanta 2019, 199, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Gago, J.; Fernie, A.R.; Nikoloski, Z.; Tohge, T.; Martorell, S.; Escalona, J.M.; Ribas-Carbó, M.; Flexas, J.; Medrano, H. Integrative field scale phenotyping for investigating metabolic components of water stress within a vineyard. Plant Methods 2017, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kerkech, M.; Hafiane, A.; Canals, R. Vine disease detection in UAV multispectral images using optimized image registration and deep learning segmentation approach. Comput. Electron. Agric. 2020, 174, 105446. [Google Scholar] [CrossRef]
- Steimetz, E.; Trouvelot, S.; Gindro, K.; Bordier, A.; Poinssot, B.; Adrian, M.; Daire, X. Influence of leaf age on induced resistance in grapevine against Plasmopara viticola. Physiol. Mol. Plant Pathol. 2012, 79, 89–96. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; ISBN 3-900051-07-0. [Google Scholar]
- Csárdi, G.; Nepusz, T. The igraph software package for complex network research. Interjournal Complex Syst. 2006, 1695, 1–9. [Google Scholar] [CrossRef]
- Fruchterman, T.M.J.; Reingold, E.M. Graph drawing by force-directed placement. Softw. Pract. Exp. 1991, 21, 1129–1164. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Retention Time (min) | Phenolic Class | Compound Assignment | Molecular Ion | Product Ions | λmax (nm) | Authentication |
|---|---|---|---|---|---|---|---|
| m01 | 1.07 | Amino acid | L-proline | 116 [M+H]+ | 70 | Standard | |
| m02 | 1.67 | Amino acid | L-leucine | 132 [M+H]+ | 86 | Standard | |
| m03 | 1.80 | Amino acid | L-isoleucine | 132 [M+H]+ | 86 | Standard | |
| m04 | 2.78 | Amino acid | L-phenylalanine | 166 [M+H]+ | 120 | Standard | |
| m05 | 1.46 | Amino acid | L-tyrosine | 182 [M+H]+ | 165 | Standard | |
| m06 | 4.12 | Amino acid | L-tryptophan | 205 [M+H]+ | 188 | Standard | |
| m07 | 1.44 | Organic acid | Citric acid | 191 [M-H]− | 111 | 327 | Standard |
| m08 | 10.48 | Stilbenoid DP1 | E-resveratrol | 227 [M-H]− | 143, 185 | 305, 317 | Standard |
| m09 | 8.72 | Stilbenoid DP1 | E-piceatannol | 243 [M-H]− | 159, 201 | 322 | Standard |
| m10 | 5.05 | Flavan-3-ol | Catechin | 289 [M-H]− | 203, 123 | 229, 278 | Standard |
| m11 | 6.20 | Flavan-3-ol | Epicatechin | 289 [M-H]− | 203, 123 | 229, 278 | Standard |
| m12 | 4.97 | Phenolic acid | Coutaric acid | 295 [M-H]− | 163, 149 | Standard | |
| m13 | 3.92 | Phenolic acid | Caftaric acid | 311 [M-H]− | 179, 149 | Standard | |
| m14 | 3.15 | Flavonol | Rhamnetin | 315 [M-H]− | 165, 121 | Standard | |
| m15 | 5.40 | Phenolic acid | Fertaric acid | 325 [M-H]− | 193, 149 | Standard | |
| m16 | 6.55 | Stilbenoid DP1 | Resveratrol glucoside | 389 [M-H]− | 227, 185 | 279 | Moss et al. (2013) [29] |
| m17 | 7.96 | Stilbenoid DP1 | E-piceid | 389 [M-H]− | 227, 185 | 279 | Standard |
| m18 | 7.86 | Flavan-3-ol | Catechin gallate | 441 [M-H]− | 289, 169 | Vrhovsek et al. (2012) [30] | |
| m19 | 9.34 | Flavonol | Kaempferol 3-O-glucoside | 449 [M-H]− | 287, 153 | Standard | |
| m20 | 9.72 | Stilbenoid DP2 | Pallidol | 453 [M-H]− | 265, 359 | 232sh, 285 | Standard |
| m21 | 10.54 | Stilbenoid DP2 | Z-resveratrol dimer1 | 453 [M-H]− | 428, 265 | 225, 283 | Püssa et al. (2006) [31] |
| m22 | 11.41 | Stilbenoid DP2 | Z-resveratrol dimer2 | 453 [M-H]− | 428, 265 | 225, 283 | Püssa et al. (2006) [31] |
| m23 | 12.87 | Stilbenoid DP2 | Z-ε-viniferin | 453 [M-H]− | 347, 359, 225 | 204, 285 | Standard |
| m24 | 13.24 | Stilbenoid DP2 | E-ε-viniferin | 453 [M-H]− | 347, 359, 225 | 225sh, 323 | Standard |
| m25 | 14.31 | Stilbenoid DP2 | E-ω-viniferin | 453 [M-H]− | 347, 359, 225 | 225sh, 325 | [25,29] |
| m26 | 14.88 | Stilbenoid DP2 | E-δ-viniferin | 453 [M-H]− | 347, 359, 225 | 225sh, 327 | Standard |
| m27 | 15.80 | Stilbenoid DP2 | Z-resveratrol dimer3 | 453 [M-H]− | 428, 265 | 225, 283 | Püssa et al. (2006) [31] |
| m28 | 9.26 | Flavonol | Kaempferol 3-O-glucuronide | 461 [M-H]− | 205 | 265, 347 | Standard |
| m29 | 8.40 | Flavonol | Quercetin 3-O-glucoside | 463 [M-H]− | 301 | 256, 354 | Standard |
| m30 | 8.27 | Flavonol | Quercetin 3-O-glucuronide | 477 [M-H]− | 301 | 254, 355 | Standard |
| m31 | 7.28 | Flavonol | Myricetin 3-O-glucoside | 479 [M-H]− | 317, 271 | 262, 300sh, 353 | Standard |
| m32 | 8.27 | Flavonol | Myricetin hexoside | 479 [M-H]− | 317, 271 | 262, 300sh, 353 | Figueiredo-González et al. (2012) [32] |
| m33 | 4.53 | Flavan-3-ol | Procyanidin B1 | 577 [M-H]− | 289, 425, 407 | 280, 313 | Standard |
| m34 | 5.63 | Flavan-3-ol | Procyanidin B2 | 577 [M-H]− | 289, 425, 407 | 279 | Standard |
| m35 | 4.83 | Flavan-3-ol | Procyanidin B3 | 577 [M-H]− | 289, 425, 407 | 279 | Standard |
| m36 | 5.33 | Flavan-3-ol | Procyanidin B4 | 577 [M-H]− | 289, 425, 407 | 279 | Ehrhardt et al. (2014) [33] |
| m37 | 6.86 | Flavan-3-ol | Procyanidin B5 | 577 [M-H]− | 289, 425, 407 | 279 | Ehrhardt et al. (2014) [33] |
| m38 | 9.03 | Flavonol | Kaempferol 3-O-rutinoside | 593 [M-H]− | 285, 255 | 265, 348 | Standard |
| m39 | 8.11 | Flavonol | Quercetin 3-O-rutinoside | 609 [M-H]− | 447, 301 | 256, 354 | Standard |
| m40 | 13.36 | Stilbenoid DP3 | Resveratrol trimer1 | 679 [M-H]− | 585, 491 | Moss et al. (2013) [29] | |
| m41 | 13.57 | Stilbenoid DP3 | E-miyabenol C | 679 [M-H]− | 573, 451 | Standard | |
| m42 | 13.98 | Stilbenoid DP3 | Resveratrol trimer2 | 679 [M-H]− | 585, 491 | Moss et al. (2013) [29] | |
| m43 | 14.78 | Stilbenoid DP3 | Resveratrol trimer3 | 679 [M-H]− | 585, 491 | Moss et al. (2013) [29] | |
| m44 | 5.13 | Flavan-3-ol | Procyanidin trimer | 865 [M-H]− | 664, 576, 289 | Monagas et al. (2006) [34] | |
| m45 | 6.37 | Flavan-3-ol | Procyanidin C1 | 865 [M-H]− | 664, 576 | Standard | |
| m46 | 4.31 | Flavan-3-ol | Prodelphinidin trimer1 | 881 [M-H]− | 592, 305 | Teixera et al., (2016) [35] | |
| m47 | 5.95 | Flavan-3-ol | Prodelphinidin trimer2 | 881 [M-H]− | 592, 576 | Teixera et al., (2016) [35] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Billet, K.; Malinowska, M.A.; Munsch, T.; Unlubayir, M.; Adler, S.; Delanoue, G.; Lanoue, A. Semi-Targeted Metabolomics to Validate Biomarkers of Grape Downy Mildew Infection Under Field Conditions. Plants 2020, 9, 1008. https://doi.org/10.3390/plants9081008
Billet K, Malinowska MA, Munsch T, Unlubayir M, Adler S, Delanoue G, Lanoue A. Semi-Targeted Metabolomics to Validate Biomarkers of Grape Downy Mildew Infection Under Field Conditions. Plants. 2020; 9(8):1008. https://doi.org/10.3390/plants9081008
Chicago/Turabian StyleBillet, Kévin, Magdalena Anna Malinowska, Thibaut Munsch, Marianne Unlubayir, Sophie Adler, Guillaume Delanoue, and Arnaud Lanoue. 2020. "Semi-Targeted Metabolomics to Validate Biomarkers of Grape Downy Mildew Infection Under Field Conditions" Plants 9, no. 8: 1008. https://doi.org/10.3390/plants9081008
APA StyleBillet, K., Malinowska, M. A., Munsch, T., Unlubayir, M., Adler, S., Delanoue, G., & Lanoue, A. (2020). Semi-Targeted Metabolomics to Validate Biomarkers of Grape Downy Mildew Infection Under Field Conditions. Plants, 9(8), 1008. https://doi.org/10.3390/plants9081008