A Synopsis of Sardinian Studies: Why Is it Important to Work on Island Orchids?


 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Current Knowledge
2.2. Local Conservation Status
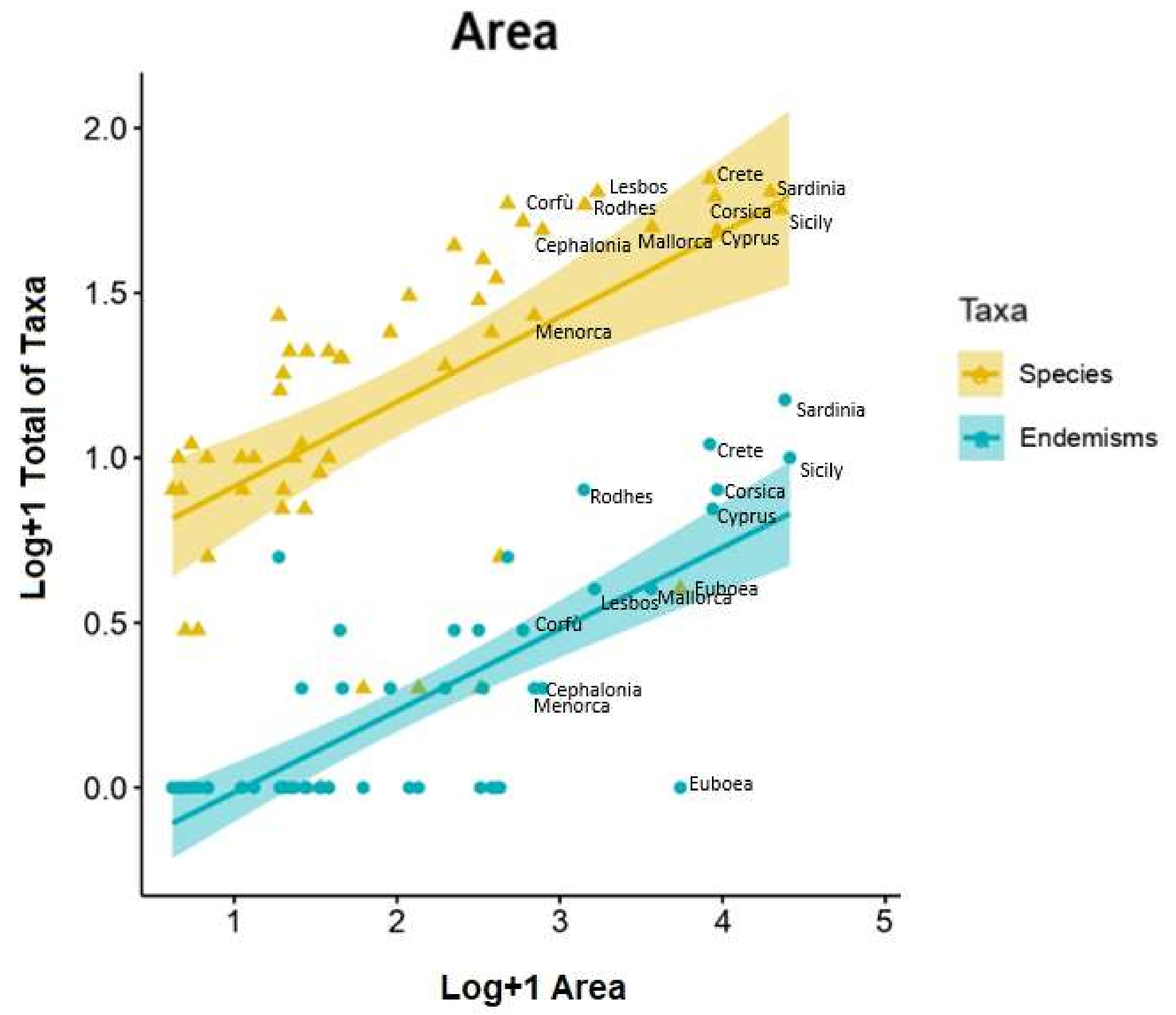
2.3. Species–Area and Species–Elevation Relationships
2.4. Key Shortfalls
3. Materials and Methods
3.1. Study Area
3.1.1. Topography
3.1.2. Geology
3.1.3. Climate
3.1.4. Plant Diversity
3.1.5. Human Impact
3.2. Check-List of Sardinian Orchids
3.3. Assessment of Local Conservation Status
3.4. Species–Area and Species–Elevation Relationships
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Darwin, C. On the Origin of Species, 6th ed.; John Murray: London, UK, 1859. [Google Scholar]
- Wallace, A.R. The Malay Archipelago: The Land of the Oranguatan, and the Bird of Paradise. A Narrative of Travel, with Studies of Man and Nature; Macmillan Publishers: New York, NY, USA, 1869. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Losos, J.B.; Schluter, L. Analysis of an evolutionary species–area relationship. Nature 2000, 408, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Triantis, K.A.; Ladle, R.J. A general dynamic theory of oceanic island biogeography. J. Biogeogr. 2008, 35, 977–994. [Google Scholar] [CrossRef]
- Mayr, E. Criteria of subspecies, species and genera in ornithology. Ann. N. Y. Acad. Sci. 1943, 44, 133–139. [Google Scholar] [CrossRef]
- Bramwell, D.; Caujapé-Castells, J. The Biology of Island Floras; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Traveset, A.; Fernández-Palacios, J.M.; Kueffer, C.; Bellingham, P.J.; Morden, C.; Drake, D.R. Introduction to the special issue: Advances in island plant biology since Sherwin Carlquist’s Island Biology. AoB PLANTS 2016, 8, plv148. [Google Scholar] [CrossRef]
- Darwin, C. On the Various Contrivances by Which British and Foreign Orchids are Fertilised by Insects, and on the Good Effects of Intercrossing; John Murray: London, UK, 1862. [Google Scholar]
- Harbaugh, D.T.; Wagner, W.L.; Allan, G.J.; Zimmer, E.A. The Hawaiian Archipelago is a stepping stone for dispersal in the Pacific: An example from the plant genus Melicope (Rutaceae). J. Biogeogr. 2009, 36, 230–241. [Google Scholar] [CrossRef]
- Ackerman, J.D.; Trejo-Torres, J.C.; Crespo-Chuy, Y. Orchids of the West Indies: Predictability of diversity and endemism. J. Biogeogr. 2007, 34, 779–786. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- McCain, C.M.; Grytnes, J.A. Elevational Gradients in Species Richness. In Encyclopedia of Life Sciences (ELS); John Wiley and Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Kerr, J.T.; Packer, L. Habitat heterogeneity as a determinant of mammal species richness in high-energy regions. Nature 1997, 385, 252–254. [Google Scholar] [CrossRef]
- Rahbek, C.; Graves, G.R. Multiscale assessment of patterns of avian species richness. Proc. Natl. Acad. Sci. USA 2001, 98, 4534–4539. [Google Scholar] [CrossRef]
- Jetz, W.; Rahbek, C. Geographic range size and determinants of avian species richness. Science 2002, 297, 1548–1551. [Google Scholar] [CrossRef]
- Kreft, H.; Jetz, W. Global patterns and determinants of vascular plant diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 5925–5930. [Google Scholar] [CrossRef]
- Médail, F.; Quézel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; van Dijk, P.P.; Rhodin, A.G.J.; Nash, S.D. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Ecoregions. Chelonian Conserv. Biol. 2004, 14, 200. [Google Scholar]
- Quézel, P. Definition of the Mediterranean region and the origin of its flora. Geobotany 1985, 7, 9–24. [Google Scholar]
- Greuter, W. Botanical diversity, endemism, rarity, and extinction in the mediterranean area: An analysis based on the published volumes of med-checklist. Bot. Chron. 1991, 10, 63–79. [Google Scholar]
- Kadereit, J.W.; Greibler, E.M.; Comes, H.P. Quaternary diversification in European alpine plants: Pattern and process. Philos Trans. R. Soc. B 2004, 59, 265–274. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Chase, M.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; Berg, C.V.D.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef]
- Bateman, R.M.; Bradshaw, E.; Devey, D.S.; Glover, B.J.; Malmgren, S.; Sramkó, G.; Thomas, M.M.; Rudall, P.J. Species arguments: Clarifying competing concepts of species delimitation in the pseudo-copulatory orchid genus Ophrys. Bot. J. Linn. Soc. 2011, 165, 336–347. [Google Scholar] [CrossRef]
- Devey, D.S.; Bateman, R.M.; Fay, M.F.; Hawkins, J.A. Friends or Relatives? Phylogenetics and Species Delimitation in the Controversial European Orchid Genus Ophrys. Ann. Bot. 2008, 101, 385–402. [Google Scholar] [CrossRef]
- Gögler, J.; Stökl, J.; Cortis, P.; Beyrle, H.; Lumaga, M.R.B.; Cozzolino, S.; Ayasse, M. Increased divergence in floral morphology strongly reduces gene flow in sympatric sexually deceptive orchids with the same pollinator. Evol. Ecol. 2015, 29, 703–717. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Araujo, M.B.; Jepson, P.; Ladle, R.J.; Watson, J.; Willis, K.J. Conservation Biogeography: Assessment and prospect. Divers. Distrib. 2005, 11, 3–23. [Google Scholar] [CrossRef]
- Blasi, C.; Marignani, M.; Copiz, R.; Fipaldini, M.; Bonacquisti, S.; Del Vico, E.; Rosati, L.; Zavattero, L. Important Plant Areas in Italy: From data to mapping. Biol. Conserv. 2011, 144, 220–226. [Google Scholar] [CrossRef]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Riddle, B.R.; Ladle, R.J.; Lourie, S.A.; Whittaker, R.J. Basic Biogeography: Estimating Biodiversity and Mapping Nature. In Conservation Biogeography; Ladle, R.J., Whittaker, R.J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2011; pp. 45–92. [Google Scholar]
- De Lange, P.; Norton, D.A.; Courtney, S.P.; Heenan, P.B.; Barkla, J.W.; Cameron, E.K.; Hitchmough, R.; Townsend, A.J. Threatened and uncommon plants of New Zealand (2008 revision). N. Z. J. Bot. 2009, 47, 61–96. [Google Scholar] [CrossRef]
- Marignani, M.; Blasi, C. Looking for important plant areas: Selection based on criteria, complementarity, or both? Biodivers. Conserv. 2012, 21, 1853–1864. [Google Scholar] [CrossRef]
- Humphreys, A.M.; Govaerts, R.; Ficinski, S.Z.; Nic Lughadha, E.; Vorontsova, M.S. Global dataset shows geography and life form predict modern plant extinction and rediscovery. Nat. Ecol. Evol. 2019, 3, 1043–1047. [Google Scholar] [CrossRef]
- Mansion, G.; Selvi, F.; Guggisberg, A.; Conti, E. Origin of Mediterranean insular endemics in the Boraginales: Integrative evidence from molecular dating and ancestral area reconstruction. J. Biogeogr. 2009, 36, 1282–1296. [Google Scholar] [CrossRef]
- Hsu, K.J.; Ryan, W.B.F.; Cita, M.B.; Hsü, K.J. Late Miocene Desiccation of the Mediterranean. Nature 1973, 242, 240–244. [Google Scholar] [CrossRef]
- Petit, R.J.; Aguinagalde, I.; De Beaulieu, J.-L.; Bittkau, C.; Brewer, S.; Cheddadi, R.; Ennos, R.; Fineschi, S.; Grivet, D.; Lascoux, M.; et al. Glacial Refugia: Hotspots But Not Melting Pots of Genetic Diversity. Science 2003, 300, 1563–1565. [Google Scholar] [CrossRef]
- Magri, D.; Fineschi, S.; Bellarosa, R.; Buonamici, A.; Sebastiani, F.; Schirone, B.; Simeone, M.C.; Vendramin, G.G. The distribution of Quercus suberchloroplast haplotypes matches the palaeogeographical history of the western Mediterranean. Mol. Ecol. 2007, 16, 5259–5266. [Google Scholar] [CrossRef]
- Pavarese, G.; Tranchida-Lombardo, V.; Cogoni, A.; Cristaudo, A.; Cozzolino, S. Where do Sardinian orchids come from: A putative African origin for the insular population of Platanthera bifolia var. kuenkelei? Bot. J. Linn. Soc. 2011, 167, 466–475. [Google Scholar] [CrossRef]
- Scrugli, A. Orchidee Spontanee della Sardegna Origine, Morfologia, Riproduzione, Impollinazione, Ibridazione, Anomalie, Fioritura; Edizioni Della Torre: Cagliari, Italy, 1990. [Google Scholar]
- Lai, R. Aggiornamento Corologico, Tassonomico, Nomenclaturale della Flora Orchidologica della Sardegna. Dottorato di Ricerca in Botanica Ambientale e Applicata. Ciclo XX; Università degli Studi di Cagliari: Cagliari, Italy, 2008. [Google Scholar]
- Hortal, J.; De Bello, F.; Diniz-Filho, J.A.; Lewinsohn, T.M.; Lobo, J.M.; Ladle, R.J. Seven Shortfalls that Beset Large-Scale Knowledge of Biodiversity Seven shortfalls biodiversity that beset knowledge on. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 523–549. [Google Scholar] [CrossRef]
- Brown, J.H.; Lomolino, M.V. Biogeography; Sinauer Associates: Sunderland, MA, USA, 1998; 691p. [Google Scholar]
- Cortis, P.; Vereecken, N.J.; Schiestl, F.; Lumaga, M.R.B.; Scrugli, A.; Cozzolino, S. Pollinator convergence and the nature of species’ boundaries in sympatric Sardinian Ophrys (Orchidaceae). Ann. Bot. 2008, 104, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.-T.; Schuiteman, A.; Chase, M.; Li, J.-W.; Chung, S.-W.; Hsu, T.-C.; Jin, X.-H. Phylogenetics of subtribe Orchidinae s.l. (Orchidaceae; Orchidoideae) based on seven markers (plastid matK, psaB, rbcL, trnL-F, trnH-psba, and nuclear nrITS, Xdh): Implications for generic delimitation. BMC Plant Biol. 2017, 17, 222. [Google Scholar] [CrossRef]
- Diniz-Filho, J.A.F.; Loyola, R.; Raia, P.; Mooers, A.O.; Bini, L.M. Darwinian shortfalls in biodiversity conservation. Trends Ecol. Evol. 2013, 28, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.M.; Sramkó, G.; Paun, O. Integrating restriction site-associated DNA sequencing (RAD-seq) with morphological cladistic analysis clarifies evolutionary relationships among major species groups of bee orchids. Ann. Bot. 2018, 121, 85–105. [Google Scholar] [CrossRef]
- Gögler, J.; Twele, R.; Francke, W.; Ayasse, M. Two phylogenetically distinct species of sexually deceptive orchids mimic the sex pheromone of their single common pollinator, the cuckoo bumblebee Bombus vestalis. Chemoecology 2011, 21, 243–252. [Google Scholar] [CrossRef]
- Scrugli, A.; Cogoni, A.; Pessei, A. Eine neue interspezifische Hybride aus Sardinien: Ophrys × panattensis Scrugli, Cogoni et Pessei, nothosp. nov. Ophrys morisii (Martelli) Soó × Ophrys scolopax Cav. Die Orchid. 1992, 43, 224–227. [Google Scholar]
- Marignani, M.; Bacchetta, G.; Bagella, S.; Caria, M.C.; Delogu, F.; Farris, E.; Fenu, G.; Filigheddu, R.; Blasi, C. Is time on our side? Strengthening the link between field efforts and conservation needs. Biodivers. Conserv. 2014, 23, 421–431. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Díaz, S.; Purvis, A.; Cornelissen, J.H.C.; Mace, G.M.; Donoghue, M.J.; Ewers, R.M.; Jordano, P.; Pearse, W.D. Functional traits, the phylogeny of function, and ecosystem service vulnerability. Ecol. Evol. 2013, 3, 2958–2975. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, J.G.; Berrigan, D.; Hill, C.E.; Hoang, A.; Gibert, P.; Beerli, P.; Hoekstra, J.M. The Strength of Phenotypic Selection in Natural Populations. Am. Nat. 2001, 157, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, G.; Pontecorvo, C.; Vacca, R. La flora del Monte Arcuentu (Sardegna sud occidentale). Webbia 2007, 62, 175–204. [Google Scholar] [CrossRef]
- Gögler, J.; Stökl, J.; Sramkova, A.; Twele, R.; Francke, W.; Cozzolino, S.; Cortis, P.; Scrugli, A.; Ayasse, M. Ménage à trois-Two endemic species of deceptive orchids and one pollinator species. Evolution 2009, 63, 2222–2234. [Google Scholar] [CrossRef] [PubMed]
- Lussu, M.; De Agostini, A.; Cogoni, A.; Marignani, M.; Cortis, P. Does size really matter? A comparative study on floral traits in orchids with two different pollination strategies. Plant Biol. 2019, 21, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Carmignani, L.; Oggiano, G.; Funedda, A.; Conti, P.; Pasci, S. The geological map of Sardinia (Italy) at 1:250,000 scale. J. Maps 2015, 12, 826–835. [Google Scholar] [CrossRef]
- Canu, S.; Rosati, L.; Fiori, M.; Motroni, A.; Filigheddu, R.; Farris, E. Bioclimate map of Sardinia (Italy). J. Maps 2014, 11, 711–718. [Google Scholar] [CrossRef]
- Spoor, F. The human fossils from Corbeddu Cave, Sardinia: A reappraisal. Deinsea 1999, 7, 297–302. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Bacchetta, G.; Fenu, G.; Mattana, E. A checklist of the exclusive vascular flora of Sardinia with priority rankings for conservation. An. Jardín Botánico Mad. 2012, 69, 81–89. [Google Scholar] [CrossRef]
- Bajocco, S.; De Angelis, A.; Perini, L.; Ferrara, A.; Salvati, L. The Impact of Land Use/Land Cover Changes on Land Degradation Dynamics: A Mediterranean Case Study. Environ. Manag. 2012, 49, 980–989. [Google Scholar] [CrossRef] [PubMed]
- Zucca, C.; Canu, A.; Previtali, F. Soil degradation by land use change in an agropastoral area in Sardinia (Italy). Catena 2010, 83, 46–54. [Google Scholar] [CrossRef]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Bonisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY—A global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Delforge, P. Orchids of Europe, North Africa and the Middle East, 1st ed.; Timber Press: Portland, OR, USA, 2006. [Google Scholar]
- Delforge, P. Guide des Orchidées d’Europe, d’Afrique du Nord et du Proche-Orient; Delachaux et Niestlé: Lonay, France, 2016. [Google Scholar]
- GIROS. Orchidee d’Italia. Ediz. Illustrata; Il Castello: Milano, Italy, 2009. [Google Scholar]
- GIROS. Orchidee d’Italia, 2nd ed; Il Castello: Milano, Italy, 2016. [Google Scholar]
- Kühn, R.; Pedersen, H.; Cribb, P. Field Guide to the Orchids of Europe and the Mediterranean; Royal Botanic Gardens, Kew: Richmond, UK, 2019. [Google Scholar]
- Liu, Q.; Chen, J.; Corlett, R.T.; Fan, X.; Yu, N.; Yang, H.; Gao, J.-Y. Orchid conservation in the biodiversity hotspot of southwestern China. Conserv. Biol. 2015, 29, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Hobohm, C. (Ed.) Endemism in Vascular Plants Hobohm; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Matthews, T.J.; Triantis, K.A.; Whittaker, R.J.; Guilhaumon, F. Sars: An R package for fitting, evaluating and comparing species—Area relationship models. Ecography 2019, 42, 1446–1455. [Google Scholar] [CrossRef]
- Wallace, A.R. On the tendency of varieties to depart indefinitely from the original type. J. Proc. Linn. Soc. 1858, 3, 53–62. [Google Scholar]
- Hamilton, T.H.; Rubinoff, I.; Barth, R.H.; Bush, G.L. Species Abundance: Natural Regulation of Insular Variation. Science 1963, 142, 1575–1577. [Google Scholar] [CrossRef]
- Johnson, M.P.; Raven, P.H. Species Number and Endemism: The Galapagos Archipelago Revisited. Science 1973, 179, 893–895. [Google Scholar] [CrossRef]
- Robinson, G.R.; Yurlina, M.E.; Handel, S.N. A Century of Change in the Staten Island Flora: Ecological Correlates of Species Losses and Invasions. Bull. Torrey Bot. Club 1994, 121, 119–129. [Google Scholar] [CrossRef]
- Aggemyr, E.; Cousins, S.A.O. Landscape structure and land use history influence changes in island plant composition after 100 years. J. Biogeogr. 2012, 39, 1645–1656. [Google Scholar] [CrossRef]
- Chiarucci, A.; Fattorini, S.; Foggi, B.; Landi, S.; Lazzaro, L.; Podani, J.; Simberloff, D. Plant recording across two centuries reveals dramatic changes in species diversity of a Mediterranean archipelago. Sci. Rep. 2017, 7, 5415. [Google Scholar] [CrossRef] [PubMed]
- Naczk, A.M.; Kolanowska, M. Glacial Refugia and Future Habitat Coverage of Selected Dactylorhiza Representatives (Orchidaceae). PLoS ONE 2015, 10, e0143478. [Google Scholar] [CrossRef] [PubMed]
- Pillon, Y.; Fay, M.F.; Hedrén, M.; Bateman, R.M.; Devey, D.S.; Shipunov, A.B.; Van Der Bank, M.; Chase, M.W.; Chase, M.; van der, B.W. Evolution and temporal diversification of western European polyploid species complexes in Dactylorhiza (Orchidaceae). Taxon 2007, 56, 1185–1208. [Google Scholar] [CrossRef]
- Pedersen, H. Ærenlund Dactylorhiza majalis s.l. (Orchidaceae) in acid habitats: Variation patterns, taxonomy, and evolution. Nord. J. Bot. 2002, 22, 641–665. [Google Scholar] [CrossRef]
- Thompson, J.D.; Lavergne, S.; Affre, L.; Gaudeul, M.; Debussche, M. Ecological differentiation of Mediterranean endemic plants. Taxon 2005, 54, 967–976. [Google Scholar] [CrossRef]
- Biddick, M.; Hendriks, A.; Burns, K.C. Plants obey (and disobey) the island rule. Proc. Natl. Acad. Sci. USA 2019, 116, 17632–17634. [Google Scholar] [CrossRef]
- Foster, J.B. Evolution of Mammals on Islands. Nature 1964, 202, 234–235. [Google Scholar] [CrossRef]
- Lomolino, M.V.; Van Der Geer, A.A.; Lyras, G.A.; Palombo, M.R.; Sax, D.F.; Rozzi, R. Of mice and mammoths: Generality and antiquity of the island rule. J. Biogeogr. 2013, 40, 1427–1439. [Google Scholar] [CrossRef]
- Bateman, R.M. Implications of next-generation sequencing for the systematics and evolution of the terrestrial orchid genus Epipactis, with particular reference to the British Isles. Kew Bull. 2020, 75, 4. [Google Scholar] [CrossRef]
- Iliadou, E.; Panitsa, M.; Dimopoulos, P. Flora and factors affecting species diversity in the islet groups of the protected “ Natura 2000” sites of the Amvrakikos Gulf and Mesologgi Lagoon (Ionian area, Greece). Willdenowia 2014, 44, 439–450. [Google Scholar] [CrossRef]
- Vogt-Schilb, H.; Pradel, R.; Geniez, P.; Hugot, L.; Delage, A.; Richard, F.; Schatz, B. Responses of orchids to habitat change in Corsica over 27 years. Ann. Bot. 2016, 118, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Fenu, G.; Fois, M.; Cañadas, E.M.; Bacchetta, G. Using endemic-plant distribution, geology and geomorphology in biogeography: The case of Sardinia (Mediterranean Basin). Syst. Biodivers. 2014, 12, 181–193. [Google Scholar] [CrossRef]
- Prather, L.A.; Alvarez-Fuentes, O.; Mayfield, M.H.; Ferguson, C.J. Implications of the decline in plant collecting for systematic and floristic research. Syst. Bot. 2004, 29, 216–220. [Google Scholar] [CrossRef]
- Robbirt, K.M.; Davy, A.J.; Hutchings, M.J.M.; Roberts, D.L. Validation of biological collections as a source of phenological data for use in climate change studies: A case study with the orchid Ophrys sphegodes. J. Ecol. 2011, 99, 235–241. [Google Scholar] [CrossRef]
- Lussu, M.; De Agostini, A.; Marignani, M.; Cogoni, A.; Cortis, P. Ophrys annae and Ophrys chestermanii: An impossible love between two orchid sister species. Nord. J. Bot. 2018, 36, e01798. [Google Scholar] [CrossRef]
- Sletvold, N.; Dahlgren, J.P.; Øien, D.-I.; Moen, A.; Ehrlén, J. Climate warming alters effects of management on population viability of threatened species: Results from a 30-year experimental study on a rare orchid. Glob. Chang. Biol. 2013, 19, 2729–2738. [Google Scholar] [CrossRef]
- Ongaro, S.; Martellos, S.; Bacaro, G.; De Agostini, A.; Cogoni, A.; Cortis, P. Distributional pattern of Sardinian Orchids under a Climate Change Distributional pattern of Sardinian Orchids under a Climate Change Scenario. Community Ecol. 2018, 19, 223–232. [Google Scholar] [CrossRef]
- Fenu, G.; Giotta, C.; Piccitto, M.; Scrugli, A.; Bacchetta, G. Dactylorhiza elata (Poir.) Soó subsp. sesquipedalis (Willd.) Soó. Italian Botanist. 2010, 42, 539–613. [Google Scholar]
- Pellegrino, G.; Bellusci, F. Effects of human disturbance on reproductive success and population viability of Serapias cordigera (Orchidaceae). Bot. J. Linn. Soc. 2014, 176, 408–420. [Google Scholar] [CrossRef]
- Coates, D.; Dixon, K. Current perspectives in plant conservation biology. Aust. J. Bot. 2007, 55, 187–193. [Google Scholar] [CrossRef]
- Swarts, N.; Dixon, K. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Parra-Tabla, V.; Vargas, C.F.; Naval, C.; Calvo, L.M.; Ollerton, J. Population Status and Reproductive Success of an Endangered Epiphytic Orchid in a Fragmented Landscape. Biotropica 2011, 43, 640–664. [Google Scholar] [CrossRef]
- Dorland, E.; Willems, J.H. High light availability alleviates the costs of reproduction in Ophrys insectifera (Orchidaceae). J. Eur. Orchid. 2006, 38, 501–518. [Google Scholar]
- Smithson, A. Pollinator limitation and inbreeding depression in orchid species with and without nectar rewards. New Phytol. 2005, 169, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Hegland, S.J.; Nielsen, A.; Lá.zaro, A.; Bjerknes, A.-L.; Totland, Ø. How does climate warming affect plant-pollinator interactions? Ecol. Lett. 2009, 12, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Bukovinszky, T.; Verheijen, J.; Zwerver, S.; Klop, E.; Biesmeijer, J.C.; Wäckers, F.L.; Prins, H.H.; Kleijn, D. Exploring the relationships between landscape complexity, wild bee species richness and reproduction, and pollination services along a complexity gradient in the Netherlands. Biol. Conserv. 2017, 214, 312–319. [Google Scholar] [CrossRef]
- Balfour, N.J.; Ollerton, J.; Castellanos, M.C.; Ratnieks, F.L. British phenological records indicate high diversity and extinction rates among late-summer-flying pollinators. Biol. Conserv. 2018, 222, 278–283. [Google Scholar] [CrossRef]
- Fay, M.F. Island plants, comparative morphology and genetic integrity. Bot. J. Linn. Soc. 2012, 170, 285–287. [Google Scholar] [CrossRef][Green Version]
- Hinsley, A.; De Boer, H.J.; Fay, M.F.; Gale, S.W.; Gardiner, L.M.; Gunasekara, R.S.; Kumar, P.; Masters, S.; Metusala, D.; Roberts, D.L.; et al. A review of the trade in orchids and its implications for conservation. Bot. J. Linn. Soc. 2017, 186, 435–455. [Google Scholar] [CrossRef]
- Fenu, G.; Bacchetta, G.; Charalambos, S.C.; Fournaraki, C.; Del Galdo, G.P.G.; Gotsiou, P.; Kyratzis, A.; Piazza, C.; Vicens, M.; Pinna, M.S.; et al. An early evaluation of translocation actions for endangered plant species on Mediterranean islands. Plant Divers. 2019, 41, 94–104. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using species distribution models at local scale to guide the search of poorly known species: Review, methodological issues and future directions. Ecol. Model. 2018, 385, 124–132. [Google Scholar] [CrossRef]
- Fois, M.; Fenu, G.; Bacchetta, G. Estimating land market values from real estate offers: A replicable method in support of biodiversity conservation strategies. Ambio 2018, 48, 313–323. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| IUCN Red List | GenBank Items | TRY Plant Trait Database | ||||||
|---|---|---|---|---|---|---|---|---|
| Species | Global | Local | ObsNum | ObsGRNum | MeasNum | MeasGRNum | TraitNum | |
| Anacamptis | ||||||||
| A. collina | Least Concern | Least Concern | 6 | |||||
| A. fragrans | Least Concern | Least Concern | 2 | |||||
| A. laxiflora | Least Concern | Vulnerable | 70 | |||||
| A. longicornu | Least Concern | 38 | ||||||
| A. palustris | Least Concern | Critically Endangered | 24 | |||||
| A. papilionacea var. papilionacea | Least Concern | Least Concern | 4 | |||||
| A. papilionacea var. grandiflora | Least Concern | 4 | ||||||
| A. pyramidalis | Least Concern | Vulnerable | 25 | 244 | 31 | 447 | 87 | 120 |
| Cephalanthera | ||||||||
| C. damasonium | Least Concern | Vulnerable | 86 | |||||
| C. longifolia | Least Concern | Least Concern | 106 | 329 | 57 | 827 | 334 | 111 |
| C. rubra | Least Concern | Vulnerable | 13 | 1 | 469 | 4 | 113 | |
| Dactylorhiza | ||||||||
| D. elata subsp. sesquipedalis | Near Threatened | Critically Endangered | ||||||
| D. insularis | Vulnerable | 2 | ||||||
| Epipactis | ||||||||
| E. gracilis | Vulnerable | |||||||
| E. helleborine | Least Concern | 1734 | 509 | 117 | 1458 | 738 | 145 | |
| E. helleborine subsp. muelleri | Least Concern | Critically Endangered | 13 | 74 | 164 | 74 | ||
| E. microphylla | Least Concern | 41 | 100 | 12 | 236 | 8 | 81 | |
| E. palustris | Least Concern | Vulnerable | 162 | 271 | 25 | 427 | 25 | 120 |
| E. tremolsii | Vulnerable | 1 | ||||||
| Gennaria | ||||||||
| G. diphylla | Least Concern | Least Concern | 11 | |||||
| Himantoglossum | ||||||||
| H. robertianum | Least Concern | Least Concern | 41 | |||||
| H. hircinum | Least Concern | Endangered | ||||||
| Limodorum | ||||||||
| L. abortivum | Least Concern | Least Concern | 97 | 93 | 234 | |||
| L. trabutianum | Near Threatened | Vulnerable | 1 | |||||
| Neotinea | ||||||||
| N. lactea | Least Concern | Least Concern | 7 | |||||
| N. maculata | Least Concern | Least Concern | 16 | |||||
| N. tridentata | Least Concern | Vulnerable | 9 | |||||
| Neottia | ||||||||
| N. nidus-avis | Least Concern | Least Concern | 84 | 434 | 21 | 701 | 23 | 127 |
| N. ovata | Vulnerable | 34 | ||||||
| Ophrys | 84 | |||||||
| O. apifera | Least Concern | Least Concern | 102 | |||||
| O. bombyliflora | Least Concern | Least Concern | 16 | 1 | ||||
| O. eleonorae | Least Concern | |||||||
| O. exaltata subsp. morisii | Least Concern | 9 | ||||||
| O. funerea | Endangered | 5 | ||||||
| O. fusca | Least Concern | Least Concern | 79 | |||||
| O. fusca subsp. ortuabis | Endangered | 0 | ||||||
| O. garganica | Vulnerable | 64 | ||||||
| O. holoserica subsp. annae | Vulnerable | 0 | ||||||
| O. holoserica subsp. chestermanii | Least Concern | 0 | ||||||
| O. incubacea | Least Concern | 17 | ||||||
| O. lutea | Least Concern | Least Concern | 26 | |||||
| O. normanii | Endangered | Endangered | 0 | |||||
| O. panattensis | Endangered | 0 | ||||||
| Ophrys scolopax subsp. apiformis | Critically Endangered | 1 | ||||||
| O. scolopax. subsp. conradiae | Vulnerable | |||||||
| O. sicula | Least Concern | 11 | ||||||
| O. speculum | Least Concern | Least Concern | 18 | |||||
| O. sphegodes subsp. praecox | Vulnerable | 0 | ||||||
| O. subfusca subsp. liveranii | Least Concern | Least Concern | 0 | |||||
| O. tenthredinifera | Least Concern | Least Concern | ||||||
| Orchis | ||||||||
| O. anthropophora | Least Concern | Least Concern | 61 | |||||
| O. brancifortii | Least Concern | Endangered | 2 | |||||
| O.italica | Least Concern | Endangered | ||||||
| Orchis mascula subsp. ichnusae | Vulnerable | 0 | ||||||
| O. provincialis | Least Concern | Least Concern | 26 | 1 | ||||
| O. purpurea | Least Concern | Endangered | 29 | |||||
| Platanthera | ||||||||
| P. algeriensis | Near Threatened | Vulnerable | 4 | |||||
| P.kuenkelei subsp. kuenkelei var. sardoa | Critically Endangered | Critically Endangered | 8 | |||||
| Serapias | ||||||||
| S. cordigera | Least Concern | Least Concern | 12 | 2 | 3 | 2 | ||
| S. lingua | Least Concern | Least Concern | 19 | 6 | 17 | 9 | ||
| S. nurrica | Near Threatened | Vulnerable | 5 | |||||
| S. parviflora | Least Concern | Least Concern | 11 | 3 | 19 | 12 | ||
| Spiranthes | ||||||||
| S. aestivalis | Data Deficient | Vulnerable | 12 | |||||
| S. spiralis | Least Concern | Least Concern | 47 | |||||
| Hybrid | Parental Species | Distribution |
|---|---|---|
| Anacamptis × bornemannii Asch. | Anacamptis papilionacea × Anacamptis longicornu | W-Medit. |
| Anacamptis × caccabaria Verguin | Anacamptis laxiflora × Anacamptis papilionacea | Medit.-Atl. |
| Anacamptis × sarcidani Scrugli et Grasso | Anacamptis laxiflora × Anacamptis longicornu | Endem. SA |
| Ophrys × barbaricina M. Allard et M.P.Grasso | Ophrys speculum × Ophrys morisii | Endem. SA |
| Ophrys × cosana H. Baumann et Kunkele | Ophrys bombyliflora × Ophrys incubacea | W-Medit. |
| Ophrys × daissiorum (H. Baumann, Giotta, Künkele, Lorenz & Piccitto) P. Delforge | Ophrys holoserica subsp. chestermanii × Ophrys morisii | Endem. SA |
| Ophrys × domus-maria M.P. Grasso | Ophrys apifera × Ophrys morisii | Endem. SA |
| Ophrys × fernandii Rolfe | Ophrys bombyliflora × Ophrys speculum | W-Medit. |
| Ophrys × heraultii G. Keller ex Schrenk | Ophrys tenthredinifera× Ophrys speculum | Medit. |
| Ophrys × laconensis Scrugli et Grasso | Ophrys exaltata subsp. morisii × Ophrys tenthredinifera | Endem. SA |
| Ophrys × maladroxiensis Scrugli, Todde e Cogoni | Ophrys exaltata subsp. morisii × Ophrys holoserica subsp. annae | Endem. SA |
| Ophrys × manfredoniae O. & E. Danesch | Ophrys incubacea × Ophrys tenthredinifera | W-Medit. |
| Ophrys × sommieri E.G. Camus ex Cortesi | Ophrys bombyliflora × Ophrys tenthredinifera | Medit. |
| Ophrys × spanui P. Delforge | Ophrys holoserica subsp. annae × Ophrys tenthredinifera | Endem. SA-CO |
| Ophrys × sulcitana Scrugli, Todde e Cogoni | Ophrys holoserica subsp. annae × Ophrys bombyliflora | Endem. SA |
| Ophrys × tavignanensis H. & J.M. Mathé & M. Pena | Ophrys eleonorae × Ophrys incubacea | W-Medit. |
| Orchis × penzigiana A. Camus nsubsp. sardoa Scrugli et Grasso | Orchis provincialis × Orchis mascula subsp. ichnusae | Endem. SA |
| Serapias × ambigua Rouy | Serapias cordigera × Serapias lingua | Medit.-Atl. |
| Serapias × cortoghianae, Grasso M.P | Serapias nurrica × Serapias cordigera | W-Medit. |
| Serapias × semilingua E.G. Camus et al. | Serapias lingua × Serapias parviflora | Medit.-Atl. |
| Chorological Rank | Taxa |
|---|---|
| Endem. SA | Ophrys holoserica subsp. chestermanii |
| Endem. SA | Ophrys normanii |
| Endem. SA | Ophrys fusca subsp. ortuabis |
| Endem. SA | Ophrys panattensis |
| Endem. SA | Ophrys subfusca subsp. liveranii |
| Endem. SA-CO | Ophrys holoserica subsp. annae |
| Endem. SA-CO | Ophrys funerea |
| Endem. SA-CO | Ophrys exaltata subsp. morisii |
| Endem. SA-CO | Ophrys scolopax subsp. conradiae |
| Endem. SA-CO | Ophrys sphegodes subsp. praecox |
| Endem. SA-CO | Orchis mascula subsp. ichnusae |
| Endem. SA-SI | Orchis brancifortii |
| Endem. SA-TU | Platanthera kuenkelei subsp. kuenkelei var. sardoa |
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anacamptis | ||||||||||||||||||||||||
| A. collina | ||||||||||||||||||||||||
| A. fragrans | ||||||||||||||||||||||||
| A. laxiflora | ||||||||||||||||||||||||
| A. longicornu | ||||||||||||||||||||||||
| A. palustris | ||||||||||||||||||||||||
| A. papilionacea subsp. grandiflora | ||||||||||||||||||||||||
| A. papilionacea subsp. papilionacea | ||||||||||||||||||||||||
| A. pyramidalis | ||||||||||||||||||||||||
| Cephalanthera | ||||||||||||||||||||||||
| C. damasonium | ||||||||||||||||||||||||
| C. longifolia | ||||||||||||||||||||||||
| C. rubra | ||||||||||||||||||||||||
| Dactylorhiza | ||||||||||||||||||||||||
| D. elata subsp. sesquipedalis | ||||||||||||||||||||||||
| D. insularis | ||||||||||||||||||||||||
| Epipactis | ||||||||||||||||||||||||
| E. helleborine | ||||||||||||||||||||||||
| E. helleborine subsp. muelleri | ||||||||||||||||||||||||
| E. microphylla | ||||||||||||||||||||||||
| E. palustris | ||||||||||||||||||||||||
| E. persica subsp. exilis | ||||||||||||||||||||||||
| E. tremolsii | ||||||||||||||||||||||||
| Gennaria | ||||||||||||||||||||||||
| G. diphylla | ||||||||||||||||||||||||
| Himantoglossum | ||||||||||||||||||||||||
| H. robertianum | ||||||||||||||||||||||||
| H. hircinum | ||||||||||||||||||||||||
| Limodorum | ||||||||||||||||||||||||
| L. abortivum | ||||||||||||||||||||||||
| L. trabutianum | ||||||||||||||||||||||||
| Neotinea | ||||||||||||||||||||||||
| N. lactea | ||||||||||||||||||||||||
| N. maculata | ||||||||||||||||||||||||
| N. tridentata | ||||||||||||||||||||||||
| Neottia | ||||||||||||||||||||||||
| N. nidus-avis | ||||||||||||||||||||||||
| N. ovata | ||||||||||||||||||||||||
| Ophrys | ||||||||||||||||||||||||
| O. apifera | ||||||||||||||||||||||||
| O. bombyliflora | ||||||||||||||||||||||||
| O. eleonorae | ||||||||||||||||||||||||
| O. exaltata subsp. morisii | ||||||||||||||||||||||||
| O. funerea | ||||||||||||||||||||||||
| O. fusca | ||||||||||||||||||||||||
| O. fusca subsp. ortuabis | ||||||||||||||||||||||||
| O. garganica | ||||||||||||||||||||||||
| O. holoserica subsp. annae | ||||||||||||||||||||||||
| O. holoserica subsp. chestermanii | ||||||||||||||||||||||||
| O. incubacea | ||||||||||||||||||||||||
| O. lutea | ||||||||||||||||||||||||
| O. normanii | ||||||||||||||||||||||||
| O. panattensis | ||||||||||||||||||||||||
| O.scolopax subsp. apiformis | ||||||||||||||||||||||||
| O. scolopax. subsp. conradiae | ||||||||||||||||||||||||
| O. sicula | ||||||||||||||||||||||||
| O. speculum | ||||||||||||||||||||||||
| O. sphegodes subsp. praecox | ||||||||||||||||||||||||
| O. subfusca subsp. liveranii | ||||||||||||||||||||||||
| O. tenthredinifera | ||||||||||||||||||||||||
| Orchis | ||||||||||||||||||||||||
| O. anthropophora | ||||||||||||||||||||||||
| O. brancifortii | ||||||||||||||||||||||||
| O. italica | ||||||||||||||||||||||||
| O. mascula subsp. ichnusae | ||||||||||||||||||||||||
| O. provincialis | ||||||||||||||||||||||||
| O. purpurea | ||||||||||||||||||||||||
| Platanthera | ||||||||||||||||||||||||
| P. algeriensis | ||||||||||||||||||||||||
| P.kuenkelei subsp. kuenkelei var. sardoa | ||||||||||||||||||||||||
| Serapias | ||||||||||||||||||||||||
| S. cordigera | ||||||||||||||||||||||||
| S. lingua | ||||||||||||||||||||||||
| S. nurrica | ||||||||||||||||||||||||
| S. parviflora | ||||||||||||||||||||||||
| Spiranthes | ||||||||||||||||||||||||
| S. aestivalis | ||||||||||||||||||||||||
| S. spiralis | ||||||||||||||||||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lussu, M.; Marignani, M.; Lai, R.; Loi, M.C.; Cogoni, A.; Cortis, P. A Synopsis of Sardinian Studies: Why Is it Important to Work on Island Orchids? Plants 2020, 9, 853. https://doi.org/10.3390/plants9070853
Lussu M, Marignani M, Lai R, Loi MC, Cogoni A, Cortis P. A Synopsis of Sardinian Studies: Why Is it Important to Work on Island Orchids? Plants. 2020; 9(7):853. https://doi.org/10.3390/plants9070853
Chicago/Turabian StyleLussu, Michele, Michela Marignani, Roberta Lai, Maria Cecilia Loi, Annalena Cogoni, and Pierluigi Cortis. 2020. "A Synopsis of Sardinian Studies: Why Is it Important to Work on Island Orchids?" Plants 9, no. 7: 853. https://doi.org/10.3390/plants9070853
APA StyleLussu, M., Marignani, M., Lai, R., Loi, M. C., Cogoni, A., & Cortis, P. (2020). A Synopsis of Sardinian Studies: Why Is it Important to Work on Island Orchids? Plants, 9(7), 853. https://doi.org/10.3390/plants9070853