Improved Root Growth by Liming Aluminum-Sensitive Rice Cultivar or Cultivating an Aluminum-Tolerant One Does Not Enhance Fertilizer Nitrogen Recovery Efficiency in an Acid Paddy Soil


 ,
,
Abstract
1. Introduction
2. Results
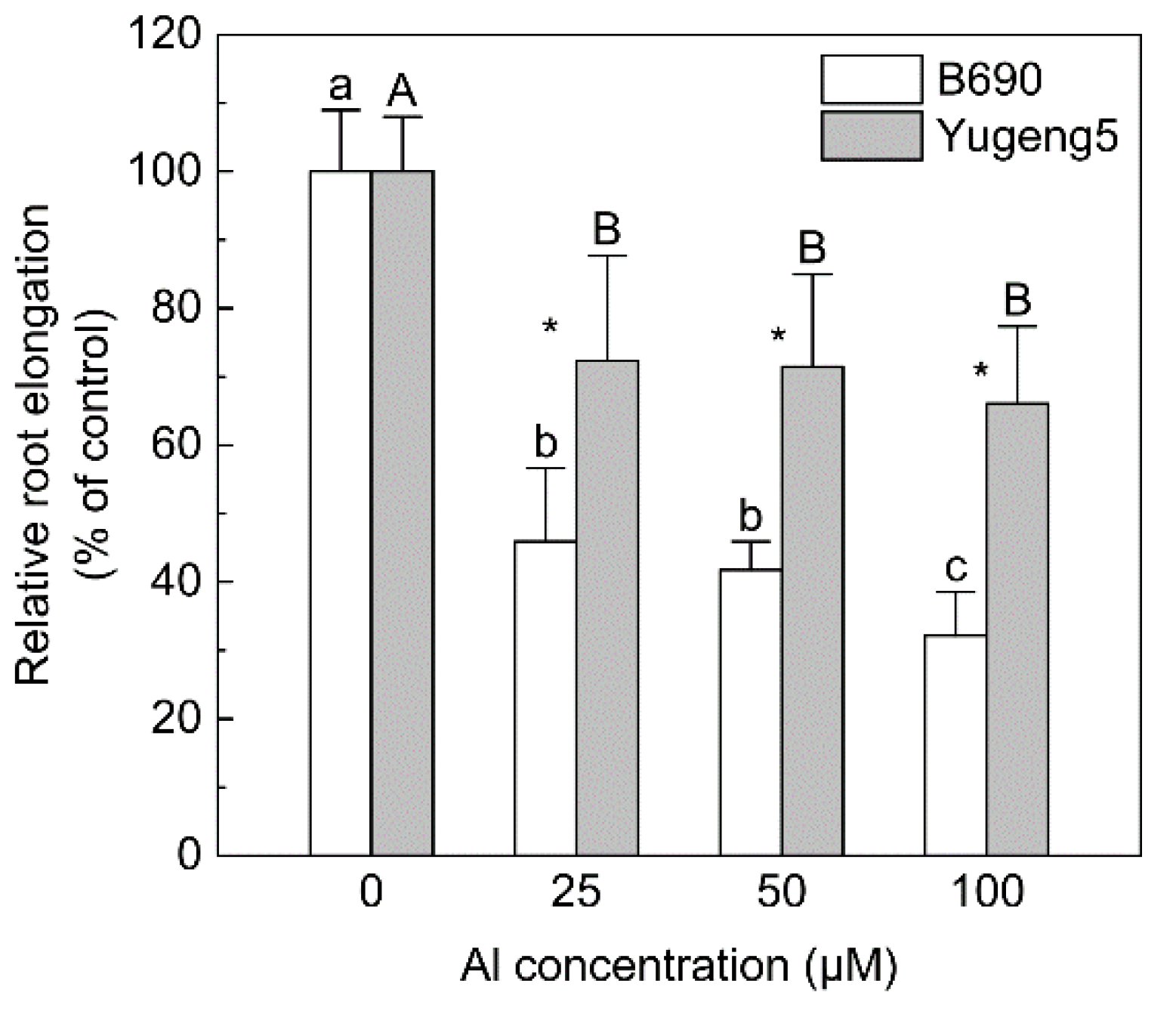
2.1. Comparison of Al Tolerance between Two Rice Cultivars
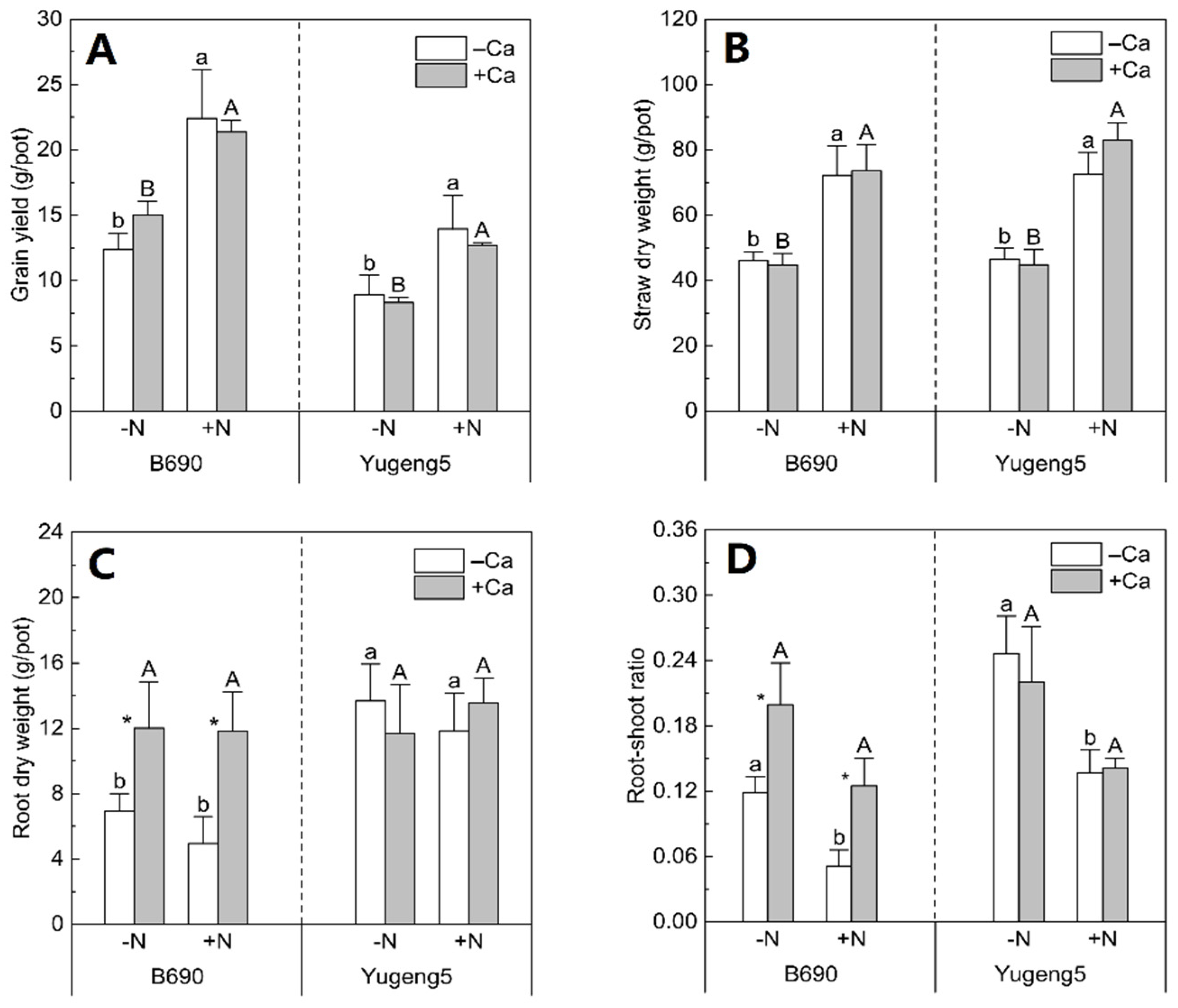
2.2. Effects of Lime and N on Biomass of Two Rice Cultivars
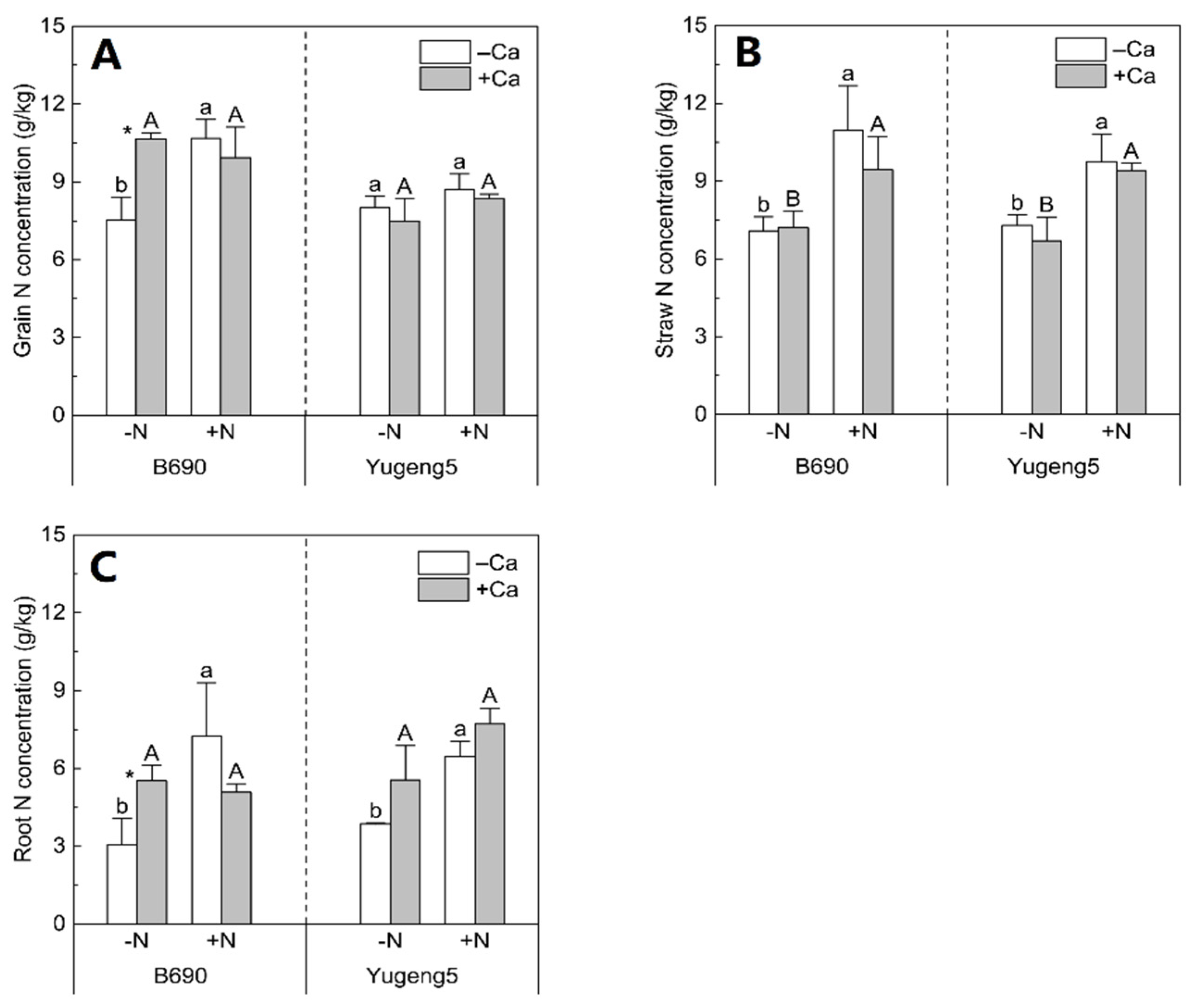
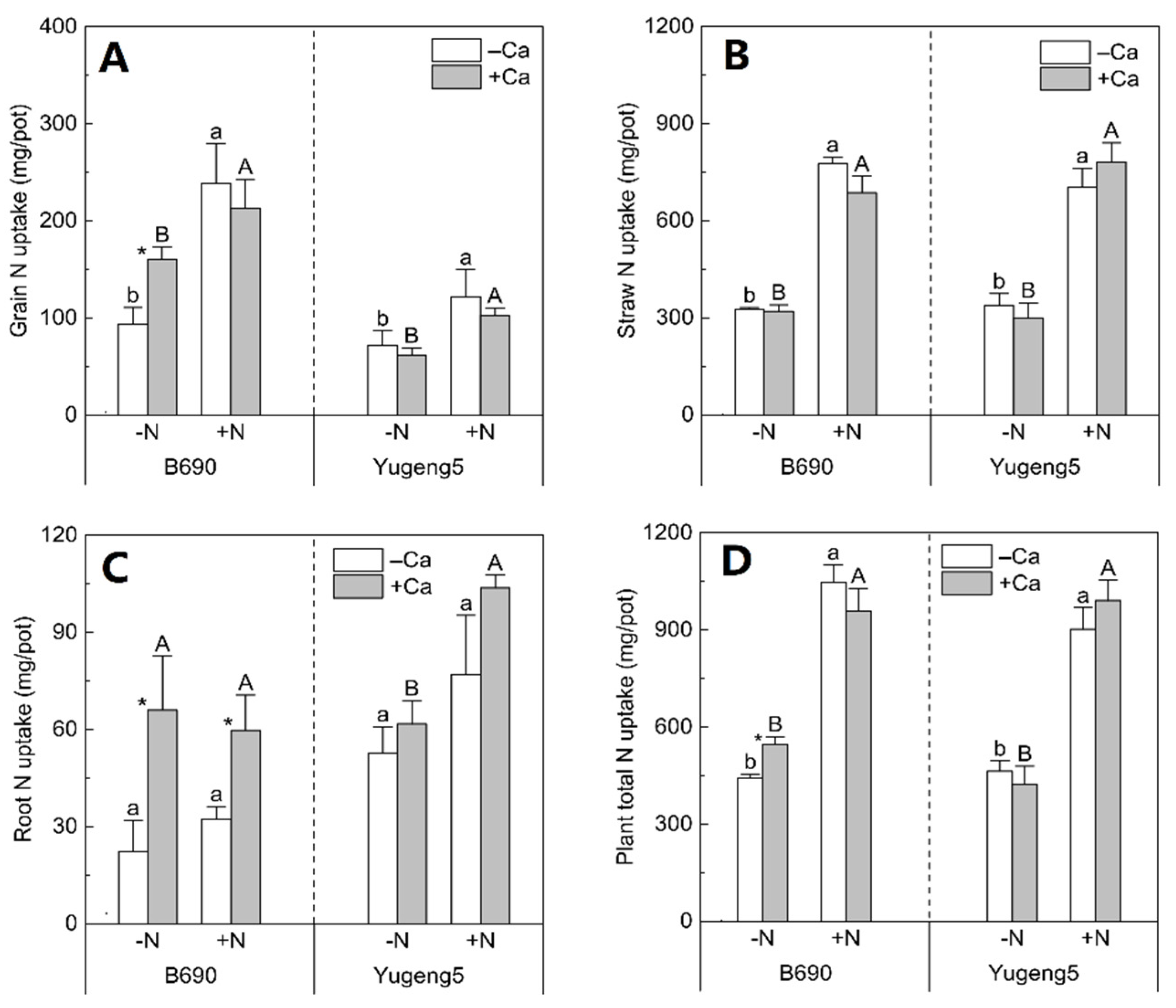
2.3. Effects of Lime and N on N Uptake of Two Rice Cultivars
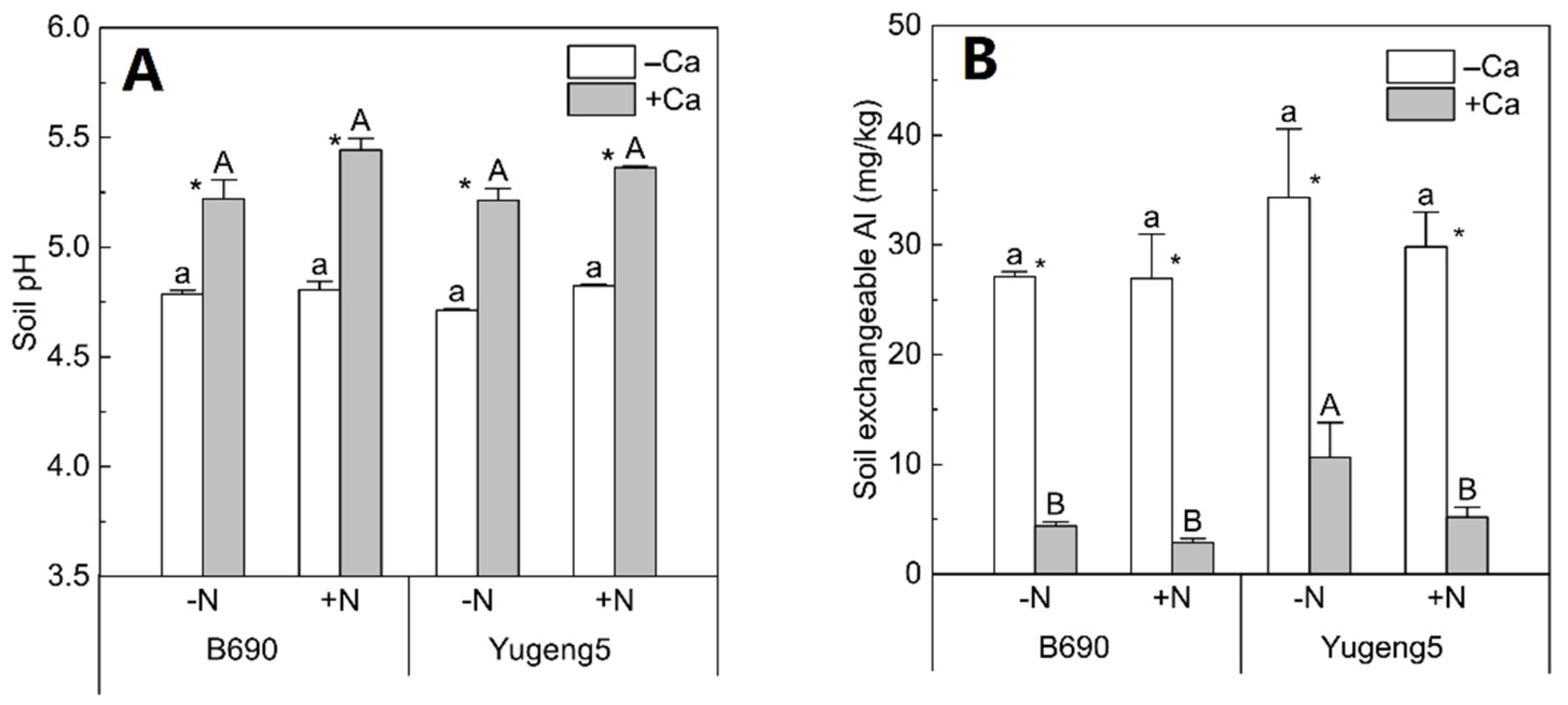
2.4. Effects of Lime and N on Soil pH and Exchangeable Al
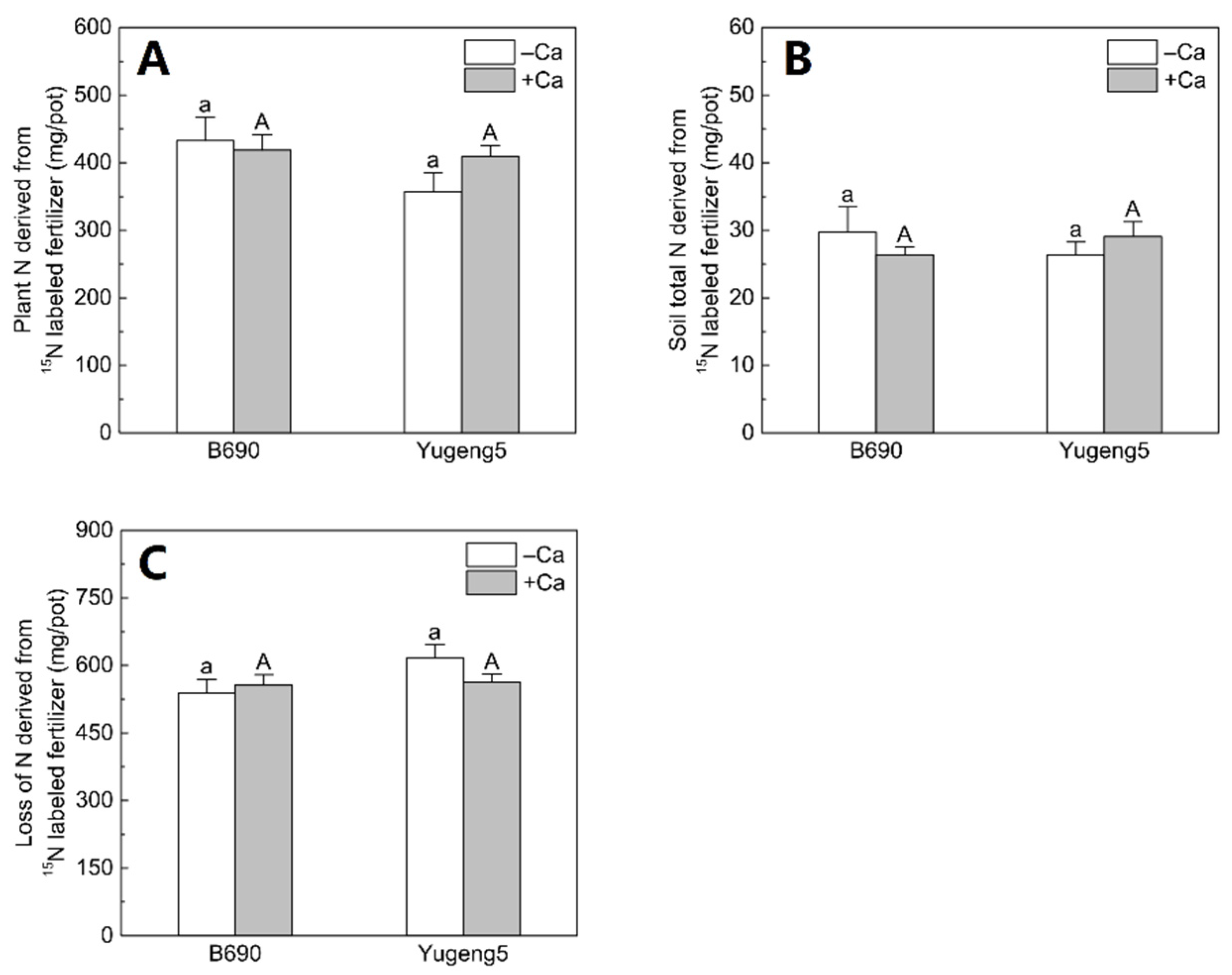
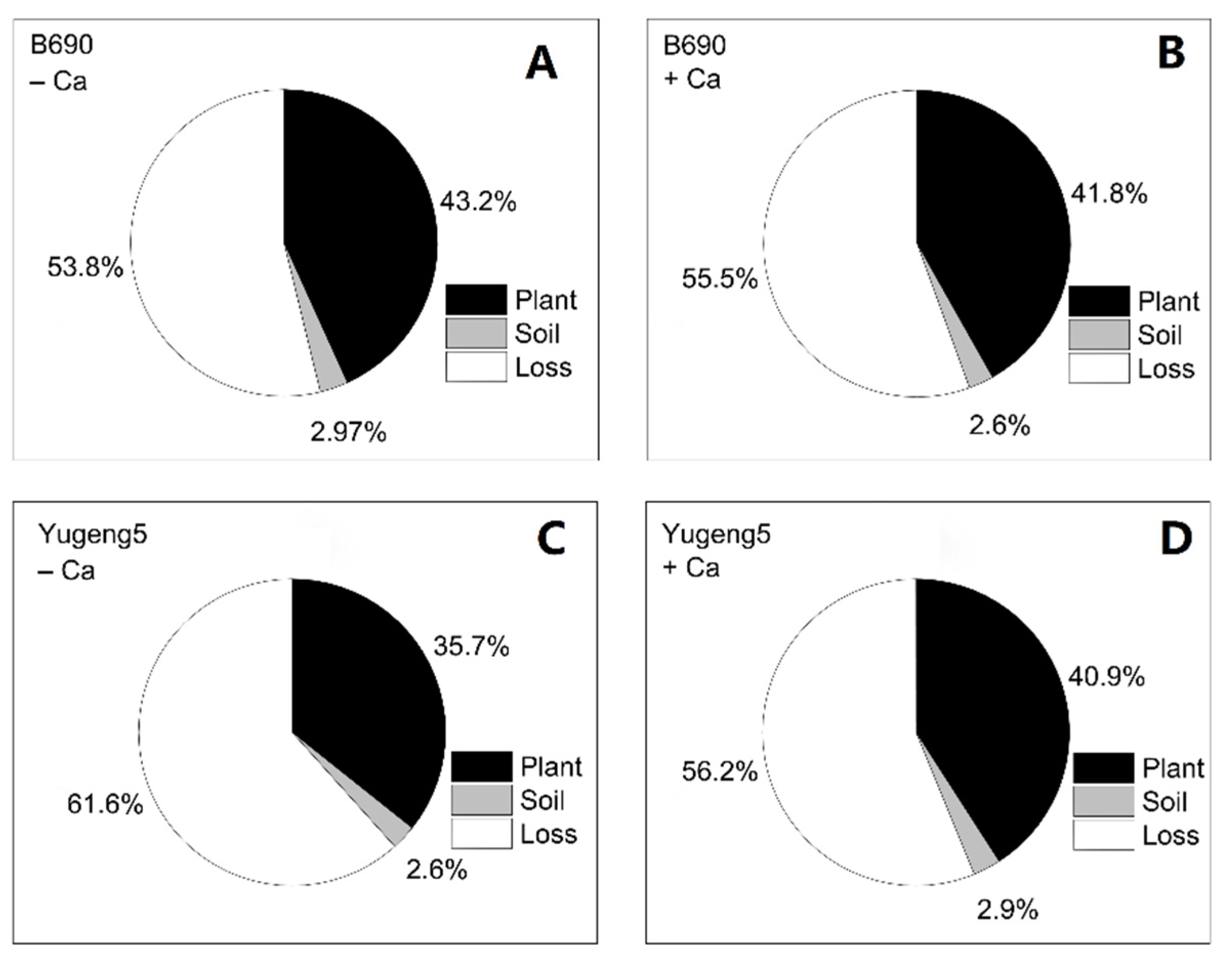
2.5. Fate of 15N-Labeled Fertilizer N in Soil–Plant Systems
2.6. Fertilizer N Recovery Efficiency
3. Discussion
3.1. Liming Improved Growth of Roots but Not Aboveground Parts of an Al-Sensitive Rice Cultivar
3.2. Liming Enhanced N Uptake of Al-Sensitive Rice Cultivar in the Absence of N Fertilizer
3.3. Cultivation of an Al-Tolerant Rice Cultivar Did Not Improve FNRE
4. Materials and Methods
4.1. Hydroponic Experiment
4.2. Pot Experiment
4.3. Analytical Methods
4.4. Calculation Methods for Fertilizer N Loss and FNRE
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Von Uexküll, H.R.; Mutert, E. Global extent, development and economic impact of acid soils. Plant Soil 1995, 171, 1–15. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Chen, R.F.; Shen, R.F. Coadaptation of plants to multiple stresses in acidic soils. Soil Sci. 2014, 179, 503–513. [Google Scholar] [CrossRef]
- Kochian, L.V.; Piñeros, M.A.; Hoekenga, O.A. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant Soil 2005, 274, 175–195. [Google Scholar] [CrossRef]
- Ma, J.F.; Chen, Z.C.; Shen, R.F. Molecular mechanisms of Al tolerance in gramineous plants. Plant Soil 2014, 381, 1–12. [Google Scholar] [CrossRef]
- Kochian, L.V. Cellular mechanisms of aluminum toxicity and resistance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Ryan, P.R.; Ditomaso, J.M.; Kochian, L.V. Aluminum toxicity in roots: An investigation of spatial sensitivity and the role of the root cap. J. Exp. Bot. 1993, 44, 437–446. [Google Scholar] [CrossRef]
- Ma, J.F.; Shen, R.; Zhao, Z.; Wissuwa, M.; Takeuchi, Y.; Ebitani, T.; Yano, M. Response of rice to Al stress and identification of quantitative trait loci for Al tolerance. Plant Cell Physiol. 2002, 43, 652–659. [Google Scholar] [CrossRef]
- Garnett, T.; Conn, V.; Kaiser, B.N. Root based approaches to improving nitrogen use efficiency in plants. Plant Cell Environ. 2009, 32, 1272–1283. [Google Scholar] [CrossRef]
- Li, X.; Zeng, R.; Liao, H. Improving crop nutrient efficiency through root architecture modifications. J. Integr. Plant Biol. 2016, 58, 193–202. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Shen, R.F. Aluminum–nitrogen interactions in the soil–plant system. Front. Plant Sci. 2018, 9, 807. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Improving nutrient use efficiency of annual crops in Brazilian acid soils for sustainable crop production. Commun. Soil Sci. Plant Anal. 2001, 32, 1303–1319. [Google Scholar] [CrossRef]
- Zhu, Z.L.; Chen, D.L. Nitrogen fertilizer use in China—Contributions to food production, impacts on the environment and best management strategies. Nutr. Cycl. Agroecosys. 2002, 63, 117–127. [Google Scholar] [CrossRef]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.; Vitousek, P.M.; Zhang, F.S. Significant acidification in major Chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.Z.; Zhao, X.Q.; Yi, X.Y.; Chen, Z.C.; Dong, X.Y.; Chen, R.F.; Shen, R.F. Excessive application of nitrogen and phosphorus fertilizers induces soil acidification and phosphorus enrichment during vegetable production in Yangtze River Delta, China. Soil Use Manag. 2013, 29, 161–168. [Google Scholar] [CrossRef]
- Schroder, J.L.; Zhang, H.; Girma, K.; Raun, W.R.; Penn, C.J.; Payton, M.E. Soil acidification from long-term use of nitrogen fertilizers on winter wheat. Soil Sci. Soc. Am. J. 2011, 75, 957–964. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, M.; Song, W.; Wen, S.; Wang, B.; Zhu, C.; Shen, R. Impact of 25 years of inorganic fertilization on diazotrophic abundance and community structure in an acidic soil in southern China. Soil Biol. Biochem. 2017, 113, 240–249. [Google Scholar] [CrossRef]
- Afreh, D.; Zhang, J.; Guan, D.; Liu, K.; Song, Z.; Zheng, C.; Deng, A.; Feng, X.; Zhang, X.; Wu, Y.; et al. Long-term fertilization on nitrogen use efficiency and greenhouse gas emissions in a double maize cropping system in subtropical China. Soil Tillage Res. 2018, 180, 259–267. [Google Scholar] [CrossRef]
- Ghosh, B.C.; Bhat, R. Environmental hazards of nitrogen loading in wetland rice fields. Environ. Pollut. 1998, 102, 123–126. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Methodology for evaluation of lowland rice genotypes for nitrogen use efficiency. J. Plant Nutr. 2003, 26, 1315–1333. [Google Scholar] [CrossRef]
- Peng, S.; Buresh, R.J.; Huang, J.; Zhong, X.; Zou, Y.; Yang, J.; Wang, G.; Liu, Y.; Hu, R.; Tang, Q.; et al. Improving nitrogen fertilization in rice by site-specific N management. A review. Agron. Sustain. Dev. 2010, 30, 649–656. [Google Scholar] [CrossRef]
- Zhou, X.; Zhou, S.; Xu, M.; Gilles, C. Evolution characteristics and influence factors of acidification in paddy soil of southern China. Sci. Agric. Sin. 2015, 48, 4811–4817. [Google Scholar]
- Zhao, X.Q.; Shen, R.F.; Sun, Q.B. Ammonium under solution culture alleviates aluminum toxicity in rice and reduces aluminum accumulation in roots compared with nitrate. Plant Soil 2009, 315, 107–121. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Guo, S.W.; Shinmachi, F.; Sunairi, M.; Noguchi, A.; Hasegawa, I.; Shen, R.F. Aluminium tolerance in rice is antagonistic with nitrate preference and synergistic with ammonium preference. Ann. Bot. 2013, 111, 69–77. [Google Scholar] [CrossRef]
- Goulding, K.W.T. Soil acidification and the importance of liming agricultural soils with particular reference to the United Kingdom. Soil Use Manag. 2016, 32, 390–399. [Google Scholar] [CrossRef]
- Ikeda, M.; Yamanishi, T. Accumulation of nitrogen supplied as ammonium in the root tips of aluminum-stressed wheat cultivars differing in aluminum sensitivity. J. Fac. Agric. Kyushu Univ. 1999, 44, 33–38. [Google Scholar]
- Famoso, A.N.; Clark, R.T.; Shaff, J.E.; Craft, E.; McCouch, S.R.; Kochian, L.V. Development of a novel aluminum tolerance phenotyping platform used for comparisons of cereal aluminum tolerance and investigations into rice aluminum tolerance mechanisms. Plant Physiol. 2010, 153, 1678–1691. [Google Scholar] [CrossRef]
- Hoyt, P.B.; Nyborg, M. Toxic metals in acid soil: I. Estimation of plant-available aluminum. Soil Sci. Soc. Am. J. 1971, 35, 236–240. [Google Scholar] [CrossRef]
- Lynch, J. Root architecture and plant productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, H.; Zhang, J. Root morphology and physiology in relation to the yield formation of rice. J. Integr. Agric. 2012, 11, 920–926. [Google Scholar] [CrossRef]
- Yamazaki, K. Root system formation and its relation to grain yield in rice plants. Korean J. Crop Sci. 1989, 34, 449–459. [Google Scholar]
- Yamazki, K.; Harada, J. The root system formation and its possible bearings on grain yield in rice plants. Jpn. Agric. Res. Q. 1982, 15, 153–160. [Google Scholar]
- Chang, C.S.; Sung, J.M. Nutrient uptake and yield responses of peanuts and rice to lime and fused magnesium phosphate in an acid soil. Field Crop Res. 2004, 89, 319–325. [Google Scholar] [CrossRef]
- Gallais, A.; Coque, M. Genetic variation and selection for nitrogen use efficiency in maize: A synthesis. Maydica 2005, 50, 531–547. [Google Scholar]
- Fageria, N.K.; Knupp, A.M. Influence of lime and gypsum on growth and yield of upland rice and changes in soil chemical properties. J. Plant Nutr. 2014, 37, 1157–1170. [Google Scholar] [CrossRef]
- Ai, C.; Liang, G.; Sun, J.; He, P.; Tang, S.; Yang, S.; Zhou, W.; Wang, X. The alleviation of acid soil stress in rice by inorganic or organic ameliorants is associated with changes in soil enzyme activity and microbial community composition. Biol. Fertil. Soils. 2015, 51, 465–477. [Google Scholar] [CrossRef]
- Liao, P.; Huang, S.; van Gestel, N.C.; Zeng, Y.; Wu, Z.; van Groenigen, K.J. Liming and straw retention interact to increase nitrogen uptake and grain yield in a double rice-cropping system. Field Crop Res. 2018, 216, 217–224. [Google Scholar] [CrossRef]
- Adams, F.; Martin, J.B. Liming effects on nitrogen use and efficiency. In Nitrogen in Crop Production, 1st ed.; Hauck, R.D., Ed.; American Society of Agronomy: Madison, WI, USA, 1984; pp. 417–426. [Google Scholar]
- Nyborg, M.; Hoyt, P.B. Effects of soil acidity and liming on mineralization of soil nitrogen. Can. J. Soil Sci. 1978, 58, 331–338. [Google Scholar] [CrossRef]
- Bailey, J.S. Liming and nitrogen efficiency: Some effects of increased calcium supply and increased soil pH on nitrogen recovery by perennial ryegrass. Commun. Soil. Sci. Plant Anal. 1995, 26, 1233–1246. [Google Scholar] [CrossRef]
- Stevens, R.J.; Laughlin, R.J. Effects of lime and nitrogen fertilizer on two sward types over a 10-year period. J. Agric. Sci. 1996, 127, 451–461. [Google Scholar] [CrossRef]
- Liu, J.; Chen, F.; Olokhnuud, C.; Glass, A.D.M.; Tong, Y.; Zhang, F.; Mi, G. Root size and nitrogen-uptake activity in two maize (Zea mays) inbred lines differing in nitrogen-use efficiency. J. Plant Nutr. Soil Sci. 2009, 172, 230–236. [Google Scholar] [CrossRef]
- Kiba, T.; Krapp, A. Plant nitrogen acquisition under low availability: Regulation of uptake and root architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil. 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Robinson, D.; Rorison, I.H. Relationship between root morphology and nitrogen availability in a recent theoretical model describing nitrogen uptake from soil. Plant Cell Environ. 1983, 6, 641–647. [Google Scholar] [CrossRef]
- Wang, Y.; Mi, G.; Chen, F.; Zhang, J.; Zhang, F. Response of root morphology to nitrate supply and its contribution to nitrogen accumulation in maize. J. Plant Nutr. 2004, 27, 2189–2202. [Google Scholar] [CrossRef]
- Peng, Y.; Niu, J.; Peng, Z.; Zhang, F.; Li, C. Shoot growth potential drives N uptake in maize plants and correlates with root growth in the soil. Field Crop Res. 2010, 115, 85–93. [Google Scholar] [CrossRef]
- Weligama, C.; Tang, C.; Sale, P.W.G.; Conyers, M.K.; Liu, D.L. Application of nitrogen in NO3− form increases rhizosphere alkalisation in the subsurface soil layers in an acid soil. Plant Soil. 2010, 333, 403–416. [Google Scholar] [CrossRef]
- Ogawa, S.; Selvaraj, M.G.; Fernando, A.J.; Lorieux, M.; Ishitani, M.; McCouch, S.; Arbelaez, J.D. N- and P-mediated seminal root elongation response in rice seedlings. Plant Soil. 2014, 375, 303–315. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.X.; Zhang, N.; Hu, B.; Jin, T.; Xu, H.; Qin, Y.; Yan, P.; Zhang, X.; Guo, X.; et al. NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 2019, 37, 676–684. [Google Scholar] [CrossRef]
- Craine, J.M.; Wedin, D.A.; Chapin III, F.S.; Reich, P.B. Relationship between the structure of root systems and resource use for 11 North American grassland plants. Plant Ecol. 2002, 165, 85–100. [Google Scholar] [CrossRef]
- Tian, Q.; Chen, F.; Zhang, F.; Mi, G. Genotypic difference in nitrogen acquisition ability in maize plants is related to the coordination of leaf and root growth. J. Plant Nutr. 2006, 29, 317–330. [Google Scholar] [CrossRef]
- Teo, Y.H.; Beyrouty, C.A.; Norman, R.J.; Gbur, E.E. Nutrient uptake relationship to root characteristics of rice. Plant Soil. 1995, 171, 297–302. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Vadez, V. Physiological traits for crop yield improvement in low N and P environments. Plant Soil. 2002, 245, 1–15. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Schrader, L.E.; Youngs, V.L. Analysis by digestion and colorimetric assay of total nitrogen in plant tissues high in nitrate. Crop Sci. 1974, 14, 854–856. [Google Scholar] [CrossRef]
- Zhang, H.Q.; Zhao, X.Q.; Chen, Y.L.; Zhang, L.Y.; Shen, R.F. Case of a stronger capability of maize seedlings to use ammonium being responsible for the higher 15N recovery efficiency of ammonium compared with nitrate. Plant Soil. 2019, 440, 293–309. [Google Scholar] [CrossRef]
- Krupnik, T.J.; Six, J.; Ladha, J.K.; Paine, M.J.; van Kessel, C. An assessment of fertilizer nitrogen recovery efficiency by grain crops. In Agriculture and the Nitrogen Cycle, 1st ed.; Mosier, A.R., Syers, K.J., Freney, J.R., Eds.; Island Press: Covelo, CA, USA, 2004; pp. 193–207. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | TN (mg/g) | SOM (mg/g) | NO3−-N (mg/kg) | NH4+-N (mg/kg) | Available P (mg/kg) | Available K (mg/kg) |
|---|---|---|---|---|---|---|
| 4.9 | 1.2 | 11.3 | 2.4 | 4.4 | 29.3 | 94.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.Q.; Zhao, X.Q.; Chen, Y.L.; Wang, J.L.; Shen, R.F. Improved Root Growth by Liming Aluminum-Sensitive Rice Cultivar or Cultivating an Aluminum-Tolerant One Does Not Enhance Fertilizer Nitrogen Recovery Efficiency in an Acid Paddy Soil. Plants 2020, 9, 765. https://doi.org/10.3390/plants9060765
Zhang HQ, Zhao XQ, Chen YL, Wang JL, Shen RF. Improved Root Growth by Liming Aluminum-Sensitive Rice Cultivar or Cultivating an Aluminum-Tolerant One Does Not Enhance Fertilizer Nitrogen Recovery Efficiency in an Acid Paddy Soil. Plants. 2020; 9(6):765. https://doi.org/10.3390/plants9060765
Chicago/Turabian StyleZhang, Hao Qing, Xue Qiang Zhao, Yi Ling Chen, Jia Lin Wang, and Ren Fang Shen. 2020. "Improved Root Growth by Liming Aluminum-Sensitive Rice Cultivar or Cultivating an Aluminum-Tolerant One Does Not Enhance Fertilizer Nitrogen Recovery Efficiency in an Acid Paddy Soil" Plants 9, no. 6: 765. https://doi.org/10.3390/plants9060765
APA StyleZhang, H. Q., Zhao, X. Q., Chen, Y. L., Wang, J. L., & Shen, R. F. (2020). Improved Root Growth by Liming Aluminum-Sensitive Rice Cultivar or Cultivating an Aluminum-Tolerant One Does Not Enhance Fertilizer Nitrogen Recovery Efficiency in an Acid Paddy Soil. Plants, 9(6), 765. https://doi.org/10.3390/plants9060765