Maize Tolerance against Drought and Chilling Stresses Varied with Root Morphology and Antioxidative Defense System

 ,
, 
 ,
,  and
and 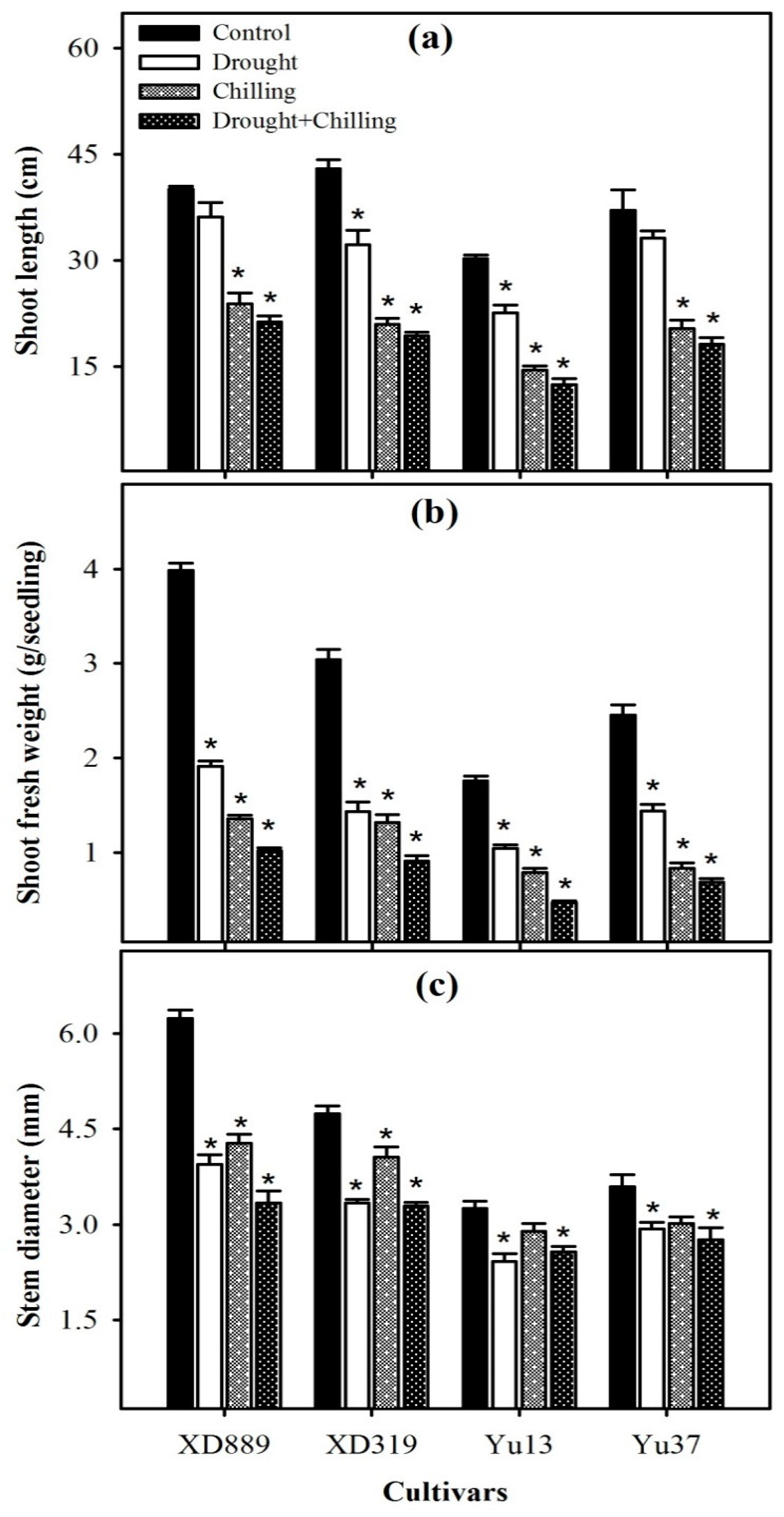{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Shoot Growth
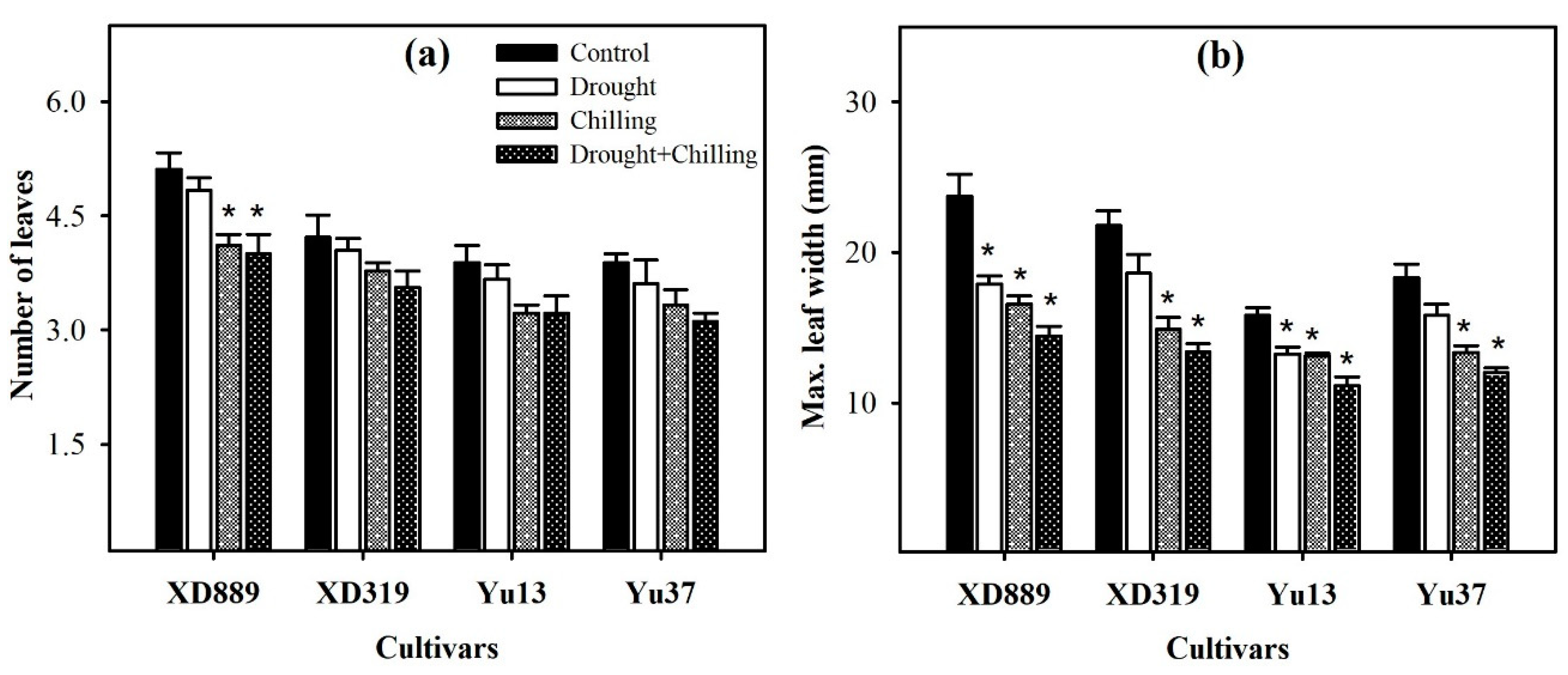
2.2. Leaf Growth
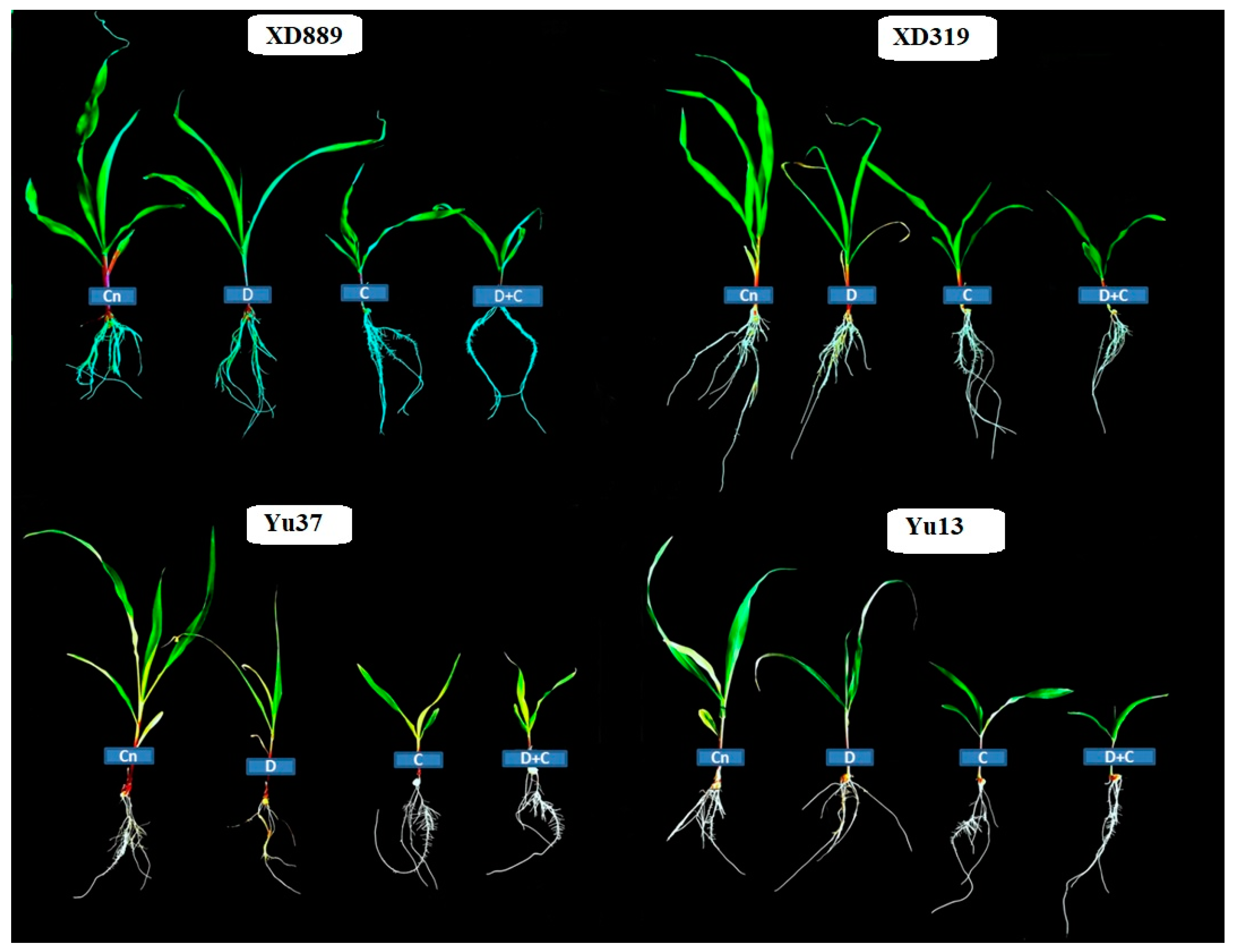
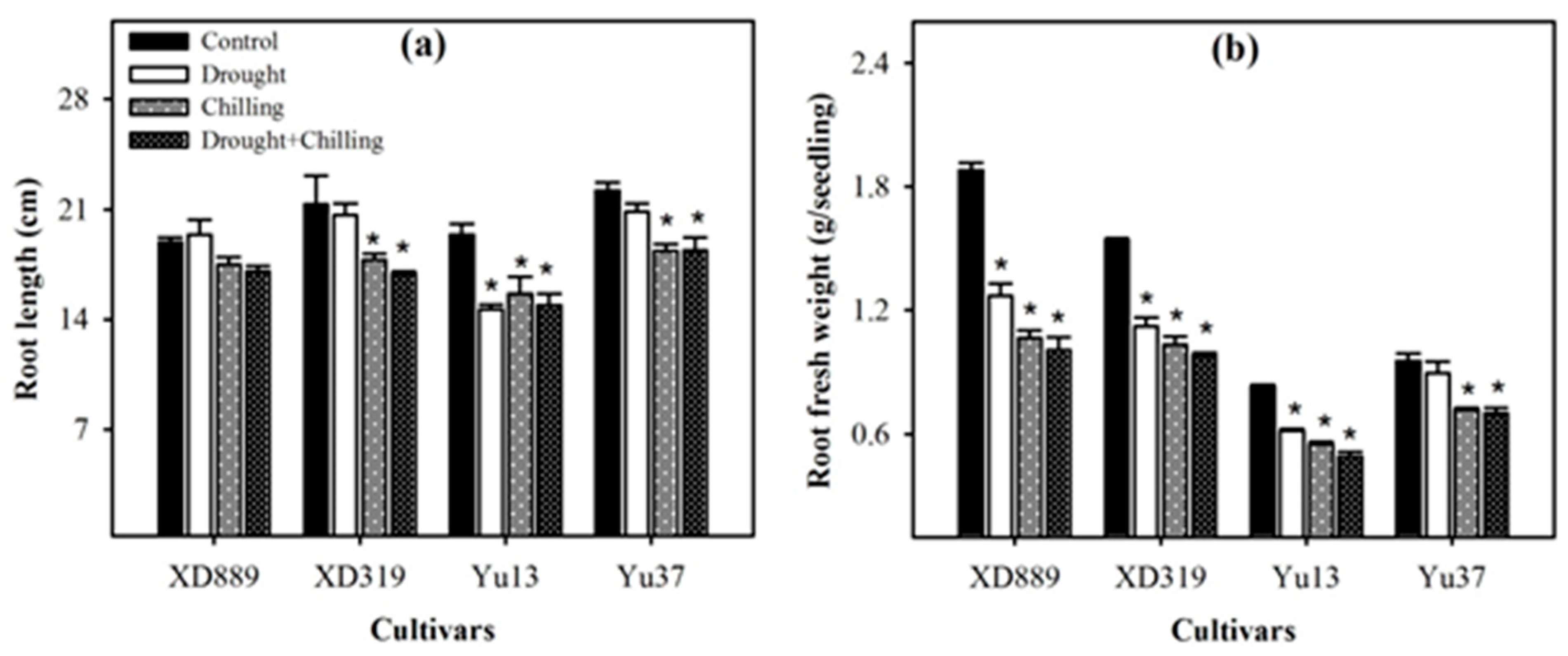
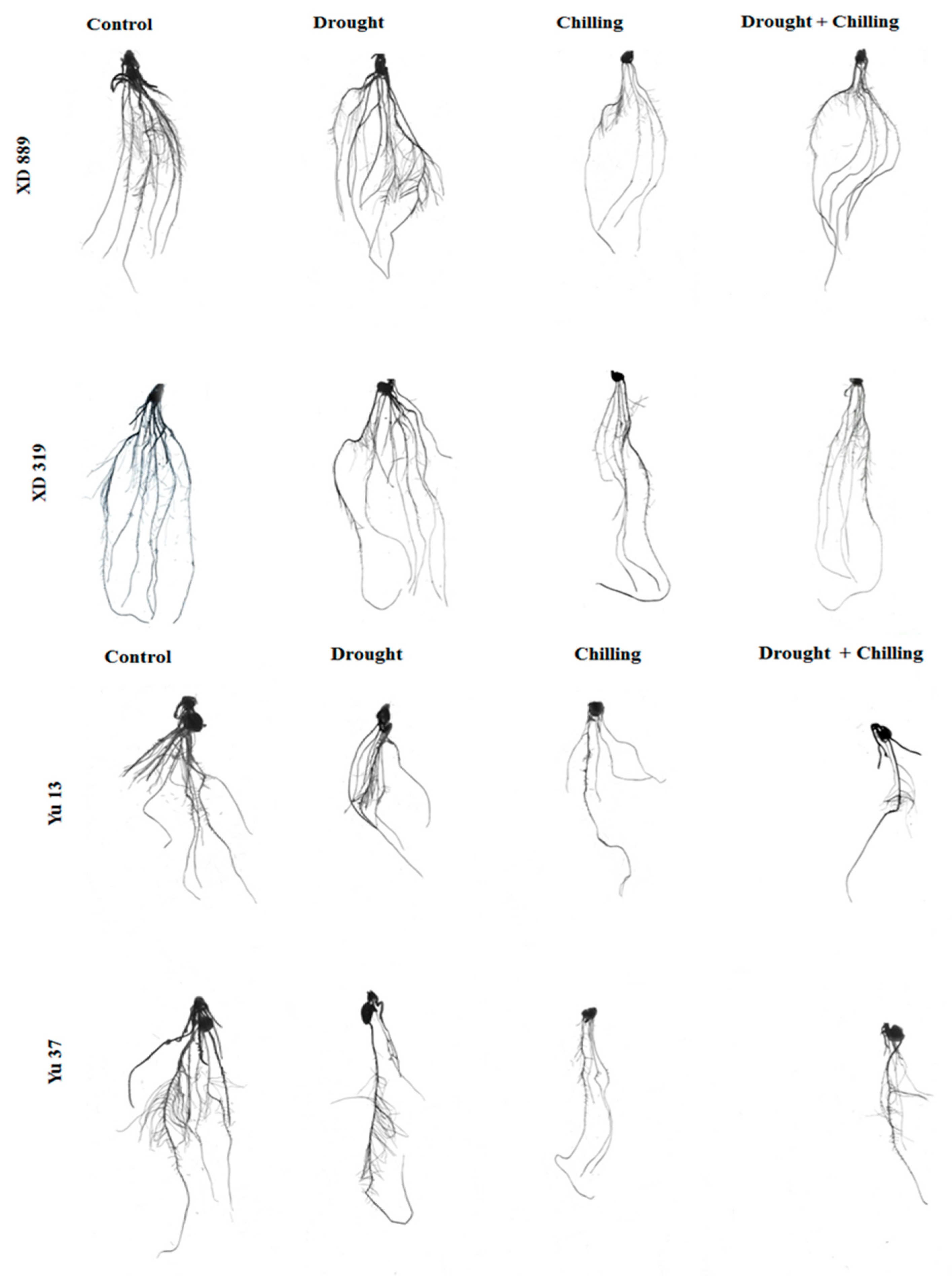
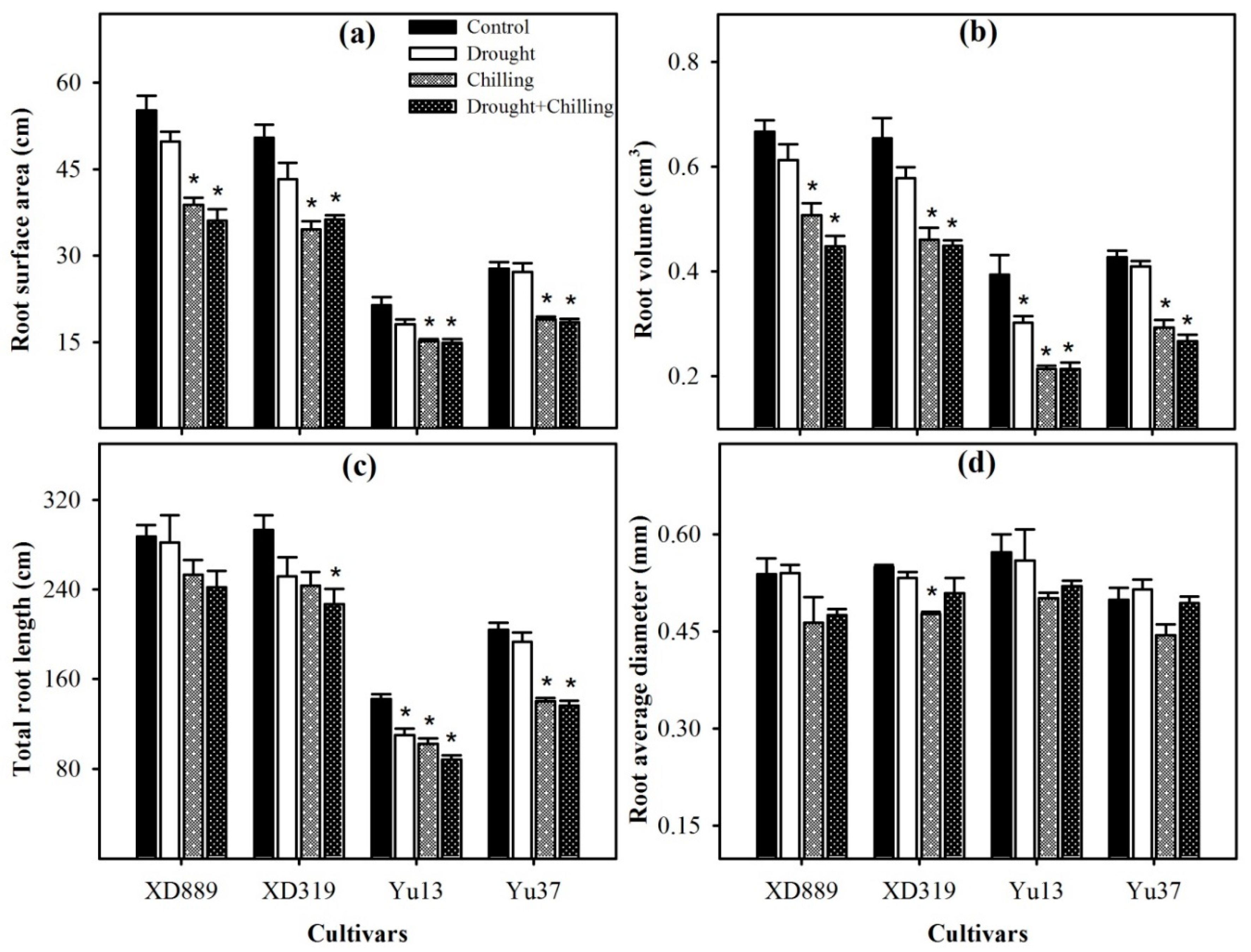
2.3. Root Growth
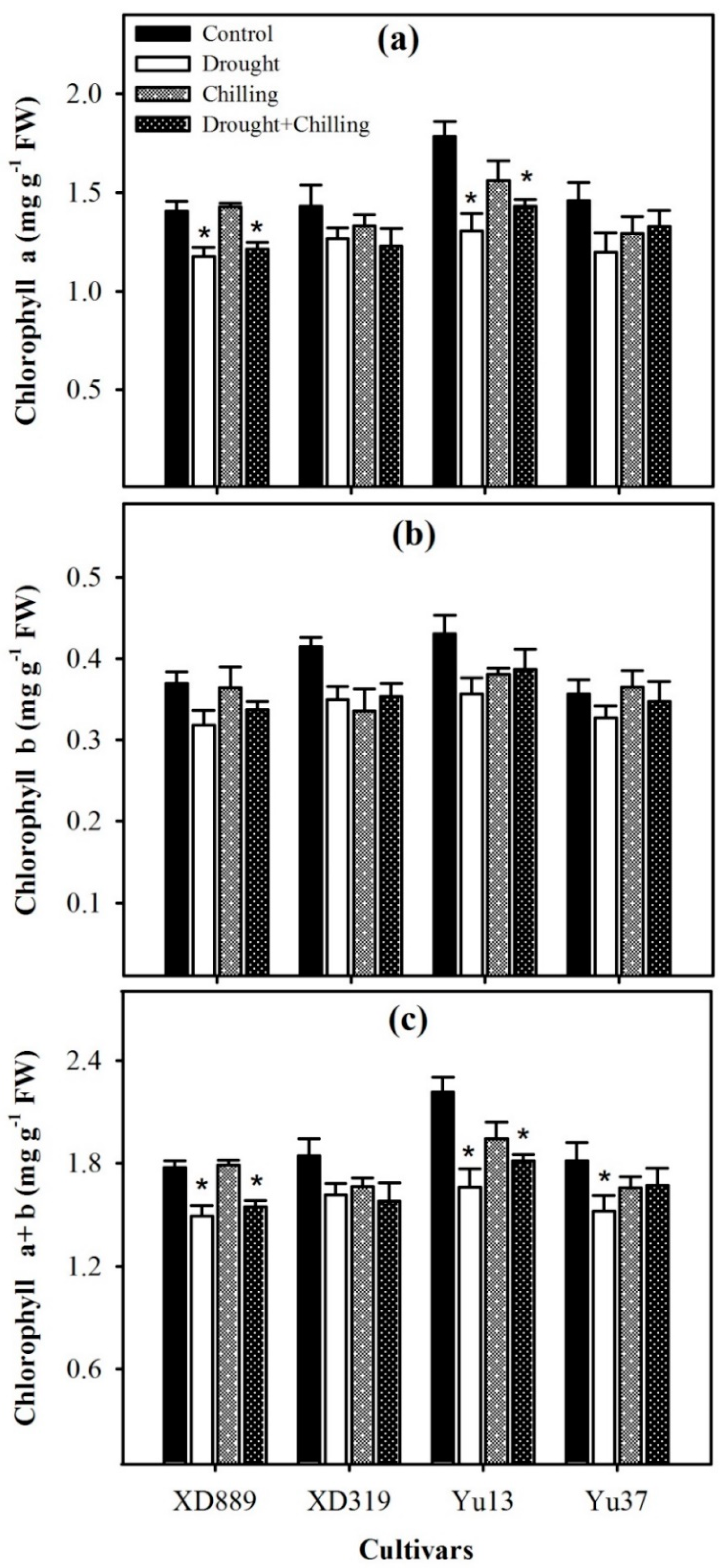
2.4. Photosynthetic Pigments
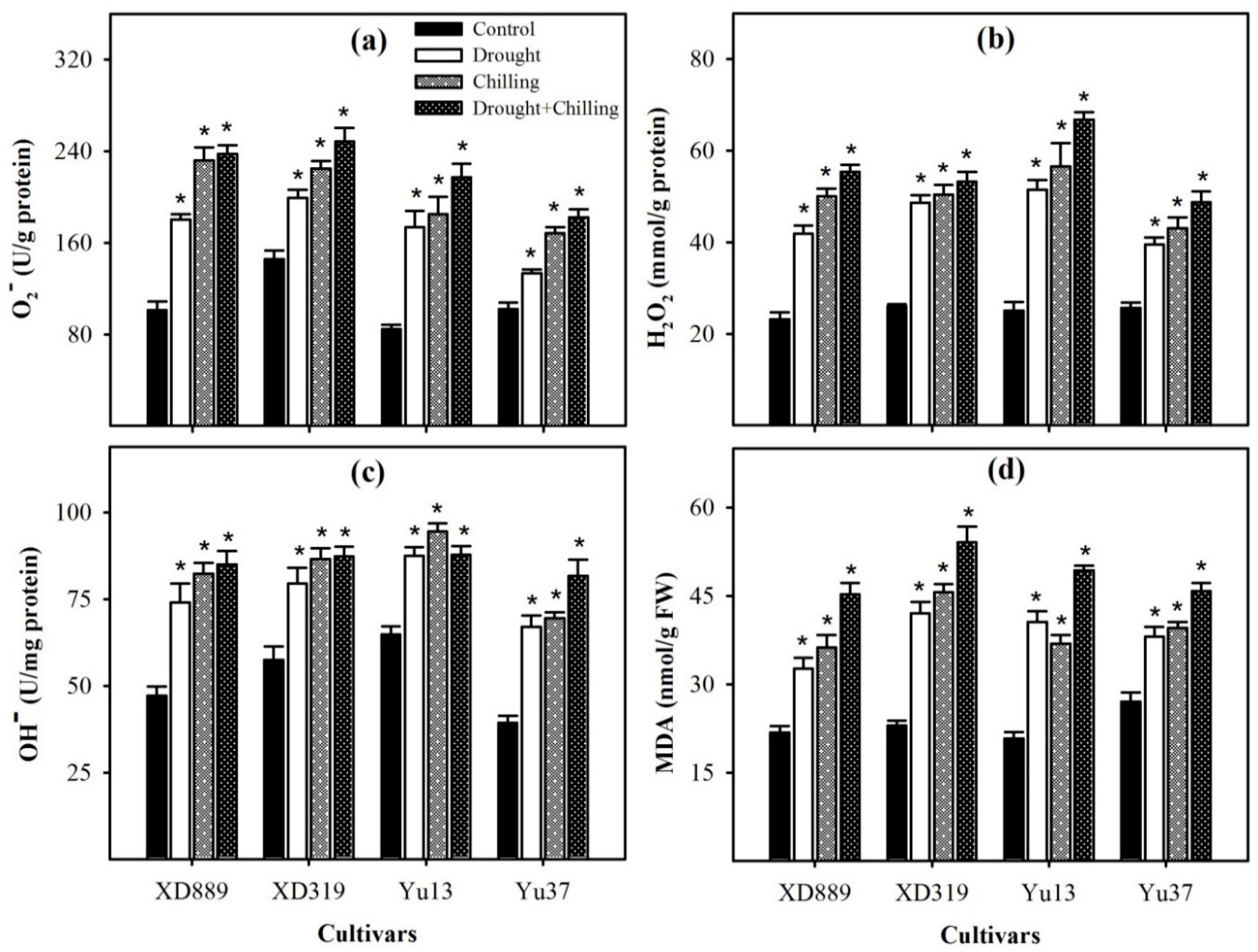
2.5. Reactive Oxygen Species and Lipid Peroxidation
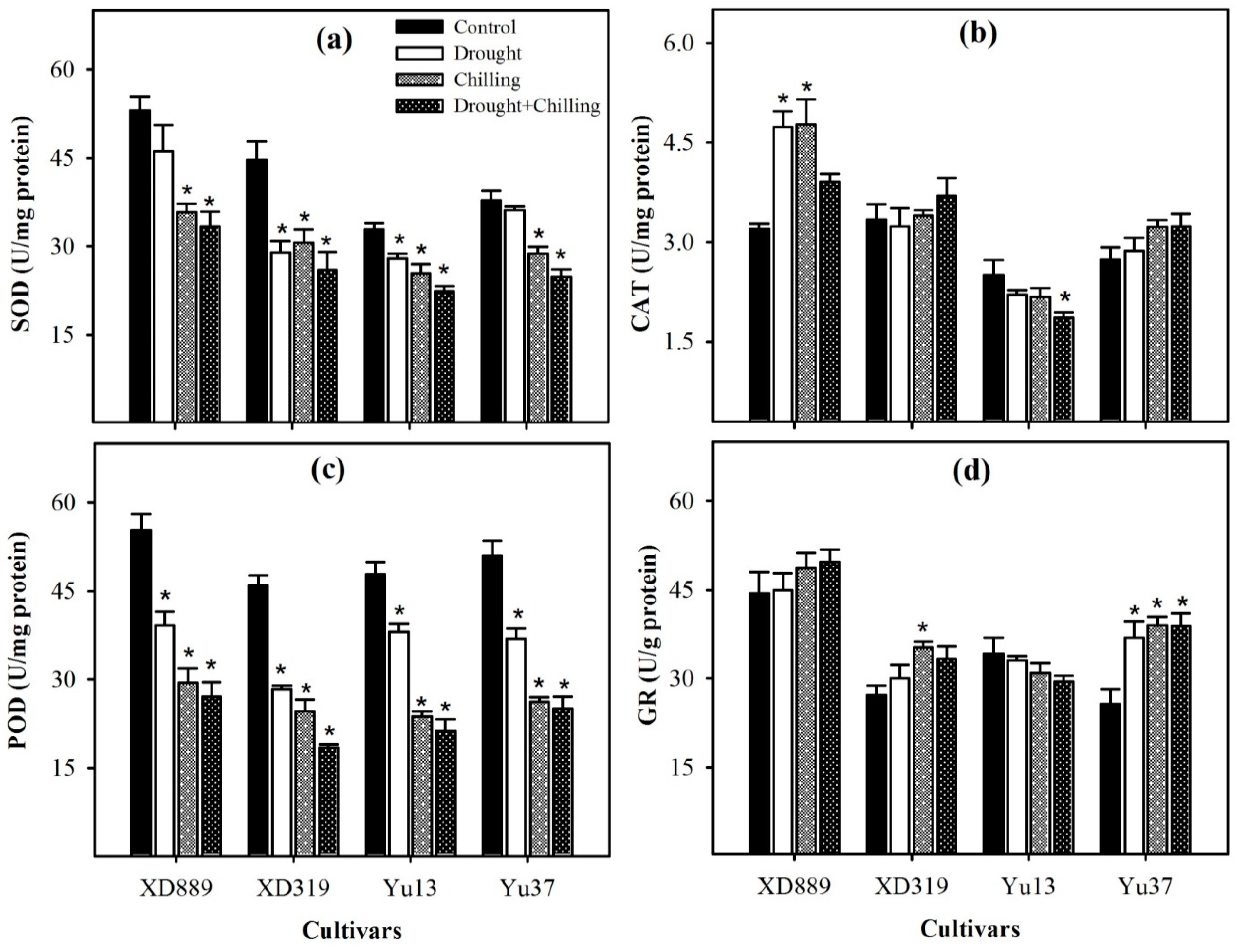
2.6. Enzymatic Antioxidants
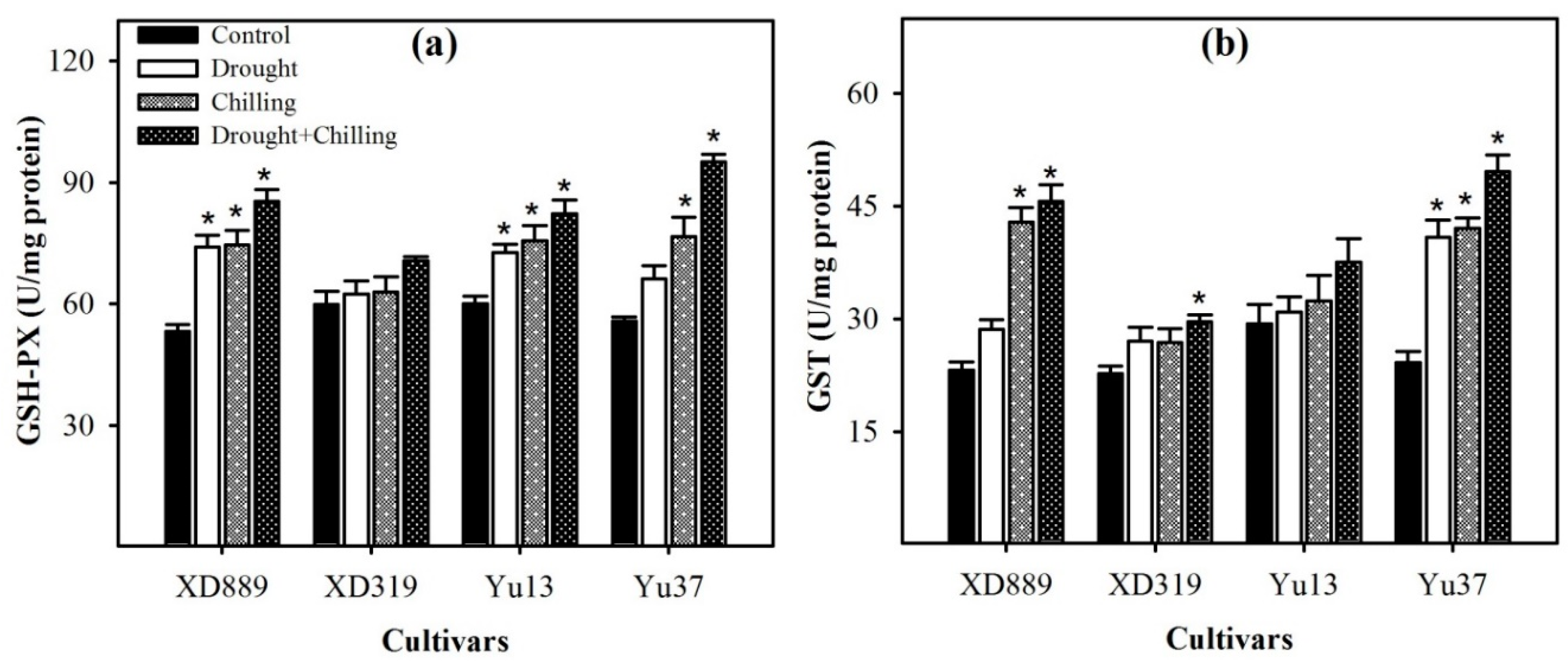
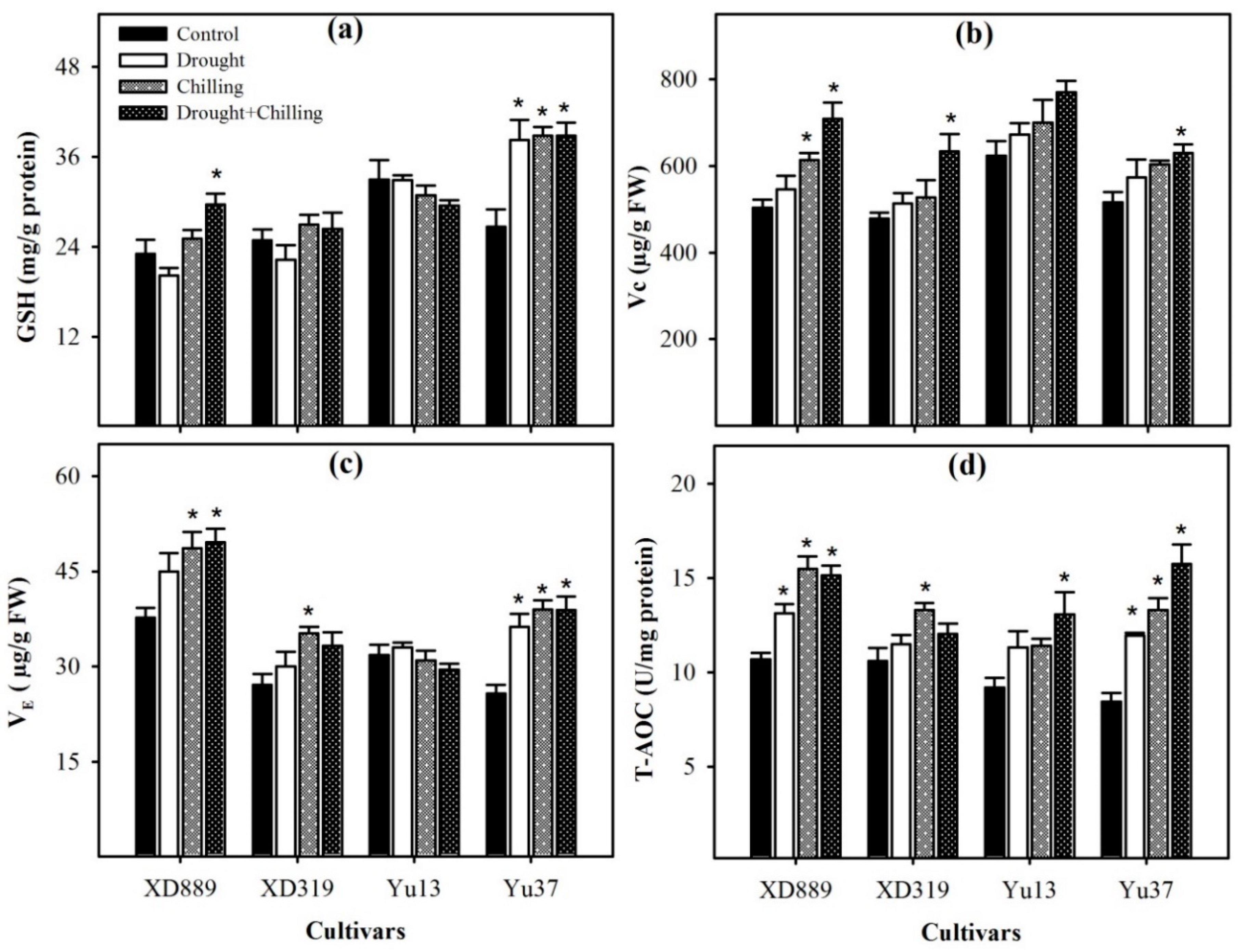
2.7. Non-Enzymatic Antioxidants and Total Antioxidant Activity
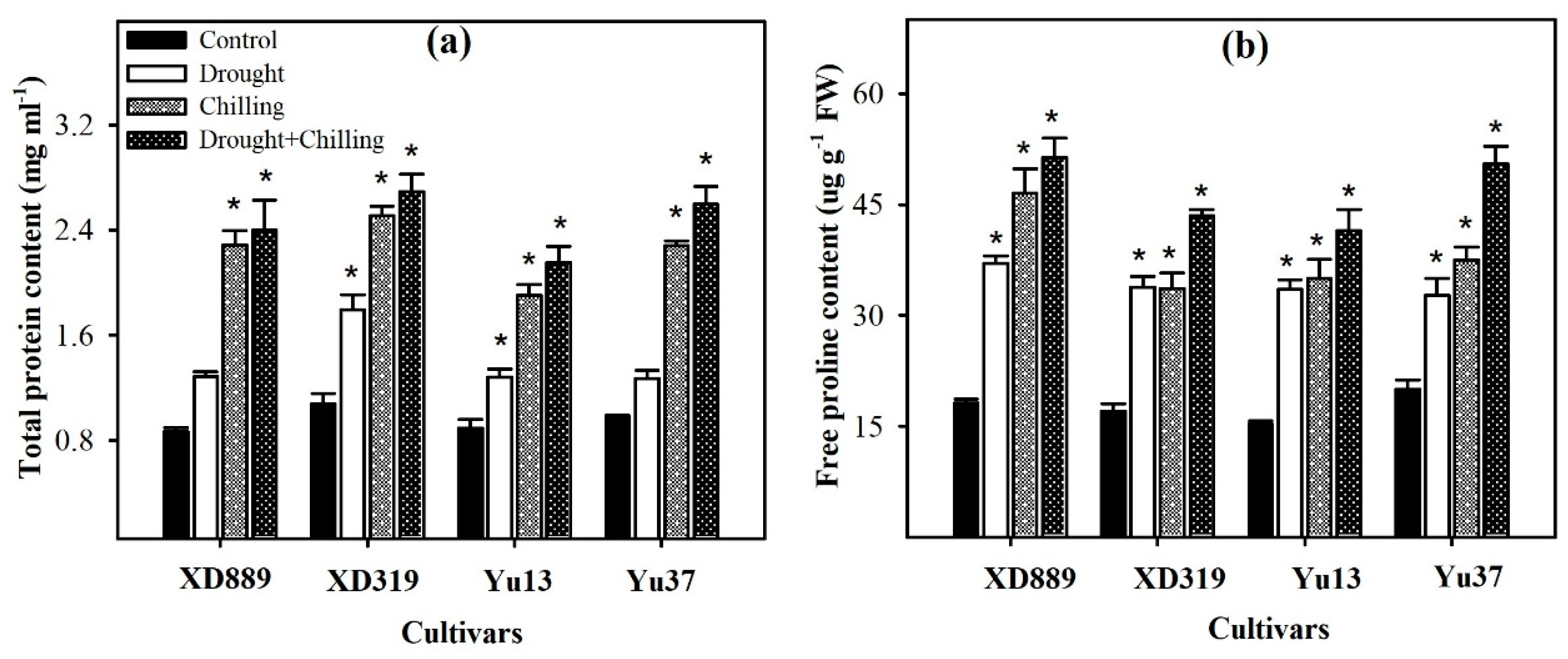
2.8. Metabolites Accumulation
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, and Treatments
4.2. Observations
4.2.1. Measurement of Growth Parameters
4.2.2. Photosynthetic Pigments
4.2.3. Lipid Peroxidation and ROS Activity
4.2.4. Estimation of Enzymatic and Non-Enzymatic Antioxidants
4.2.5. Assay of Osmolyte Accumulation Profiles
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, W.; Chen, Q.; Hussain, S.; Mei, J.; Dong, H.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Pre-sowing Seed Treatments in Direct-seeded Early Rice: Consequences for Emergence, Seedling Growth and Associated Metabolic Events under Chilling Stress. Sci. Rep. 2016, 6, 19637. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and Drought Stresses in Crop Plants: Implications, Cross Talk, and Potential Management Opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Anjum, S.A.; Hussain, S. Seed Priming Toward Enhanced Chilling Tolerance in Field Crops: An Overview. In Priming and Pretreatment of Seeds and Seedlings; Springer Nature: Singapore, 2019; pp. 265–286. [Google Scholar]
- Janowiak, F.; Markowski, A. Changes in Leaf Water Relations and Injuries in Maize Seedlings Induced by Different Chilling Conditions. J. Agron. Crop. Sci. 1994, 172, 19–28. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Juan, J.P.; Sánchez-Díaz, M. Drought enhances chilling tolerance in a chilling-sensitive maize (Zea mays) variety. New Phytol. 1996, 134, 53–59. [Google Scholar] [CrossRef]
- Cutforth, H.W.; Shaykewich, C.F.; Cho, C.M. Effect of soil water and temperature on corn (Zea mays L.) root growth during emergence. Can. J. Soil Sci. 1986, 66, 51–58. [Google Scholar] [CrossRef]
- Manivannan, P.; Jaleel, C.A.; Sankar, B.; Kishorekumar, A.; Somasundaram, R.; Lakshmanan, G.; Panneerselvam, R. Growth, biochemical modifications and proline metabolism in Helianthus annuus L. as induced by drought stress. Coll. Surf. B Biointerfaces 2007, 59, 141–149. [Google Scholar] [CrossRef]
- Hussain, S.; Hussain, S.; Qadir, T.; Khaliq, A.; Ashraf, U.; Parveen, A.; Saqib, M.; Rafiq, M. Drought stress in plants: An overview on implications, tolerance mechanisms and agronomic mitigation strategies. Plant Sci. Today 2019, 6, 389–402. [Google Scholar] [CrossRef]
- Deng, B.; Du, W.; Liu, C.; Sun, W.; Tian, S.; Dong, H. Antioxidant response to drought, cold and nutrient stress in two ploidy levels of tobacco plants: Low resource requirement confers polytolerance in polyploids? Plant Growth Regul. 2011, 66, 37–47. [Google Scholar] [CrossRef]
- Gill, S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Baier, M.; Kandlbinder, A.; Golldack, D.; Dietz, K.-J. Oxidative stress and ozone: Perception, signalling and response. Plant Cell Environ. 2005, 28, 1012–1020. [Google Scholar] [CrossRef]
- Srivalli, B.; Sharma, G.; Khanna-Chopra, R. Antioxidative defense system in an upland rice cultivar subjected to increasing intensity of water stress followed by recovery. Physiol. Plant. 2003, 119, 503–512. [Google Scholar] [CrossRef]
- Krasensky-Wrzaczek, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- Khaliq, A.; Zia-Ul-Haq, M.; Ali, F.; Aslam, F.; Matloob, A.; Navab, A.; Hussain, S. Salinity Tolerance in Wheat Cultivars Is Related to Enhanced Activities of Enzymatic Antioxidants and Reduced Lipid Peroxidation. CLEAN—Soil Air Water 2015, 43, 1248–1258. [Google Scholar] [CrossRef]
- Hussain, S.; Khaliq, A.; Tanveer, M.; Matloob, A.; Hussain, H.A. Aspirin priming circumvents the salinity-induced effects on wheat emergence and seedling growth by regulating starch metabolism and antioxidant enzyme activities. Acta Physiol. Plant. 2018, 40, 68. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought Induced Changes in Growth, Osmolyte Accumulation and Antioxidant Metabolism of Three Maize Hybrids. Front. Plant Sci. 2017, 8, 19. [Google Scholar] [CrossRef]
- Hussain, S.; Khan, F.; Hussain, H.A.; Nie, L. Physiological and Biochemical Mechanisms of Seed Priming-Induced Chilling Tolerance in Rice Cultivars. Front. Plant Sci. 2016, 7, 17022. [Google Scholar] [CrossRef]
- Hussain, S.; Khan, F.; Cao, W.; Wu, L.; Geng, M. Seed Priming Alters the Production and Detoxification of Reactive Oxygen Intermediates in Rice Seedlings Grown under Sub-optimal Temperature and Nutrient Supply. Front. Plant Sci. 2016, 7, 17022. [Google Scholar] [CrossRef]
- Guo, Z.; Ou, W.; Lu, S.; Zhong, Q. Differential responses of antioxidative system to chilling and drought in four rice cultivars differing in sensitivity. Plant Physiol. Biochem. 2006, 44, 828–836. [Google Scholar] [CrossRef]
- Farooq, M.; Aziz, T.; Wahid, A.; Lee, D.-J.; Siddique, K.H.M. Chilling tolerance in maize: Agronomic and physiological approaches. Crop. Pasture Sci. 2009, 60, 501–516. [Google Scholar] [CrossRef]
- Tommasini, L.; Svensson, J.; Rodriguez, E.M.; Wahid, A.; Malatrasi, M.; Kato, K.; Wanamaker, S.; Resnik, J.; Close, T. Dehydrin gene expression provides an indicator of low temperature and drought stress: Transcriptome-based analysis of Barley (Hordeum vulgare L.). Funct. Integr. Genom. 2008, 8, 387–405. [Google Scholar] [CrossRef] [PubMed]
- Alghabari, F.; Ihsan, M.Z.; Hussain, S.; Aishia, G.; Daur, I. Effect of Rht alleles on wheat grain yield and quality under high temperature and drought stress during booting and anthesis. Environ. Sci. Pollut. Res. 2015, 22, 15506–15515. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Jiang, D. Physiological, proteomic and transcriptional responses of wheat to combination of drought or waterlogging with late spring low temperature. Funct. Plant Boil. 2014, 41, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, M.; Liu, K.; Sui, N. Effects of Drought Stress on Seed Germination and Seedling Growth of Different Maize Varieties. J. Agric. Sci. 2015, 7, 5. [Google Scholar] [CrossRef]
- Parveen, A.; Liu, W.; Hussain, S.; Asghar, J.; Perveen, S.; Xiong, H. Silicon Priming Regulates Morpho-Physiological Growth and Oxidative Metabolism in Maize under Drought Stress. Plants 2019, 8, 431. [Google Scholar] [CrossRef]
- Bano, S.; Aslam, M.; Saleem, M.; Basra, S.M.A.; Aziz, K. Evaluation of maize accessions under low temperature stress at early growth stages. J. Anim. Plant Sci. 2015, 25, 392–400. [Google Scholar]
- Cheng, C.; Yun, K.-Y.; Ressom, H.; Mohanty, B.; Bajic, V.B.; Jia, Y.; Yun, S.J.; Reyes, B.G.D.L. An early response regulatory cluster induced by low temperature and hydrogen peroxide in seedlings of chilling-tolerant japonica rice. BMC Genom. 2007, 8, 175. [Google Scholar] [CrossRef]
- Ruelland, E.; Vaultier, M.-N.; Zachowski, A.; Hurry, V. Chapter 2 Cold Signalling and Cold Acclimation in Plants. Adv. Botanical Res. 2009, 49, 35–150. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Gopi, R.; Sankar, B.; Gomathinayagam, M.; Panneerselvam, R. Differential responses in water use efficiency in two varieties of Catharanthus roseus under drought stress. Comptes Rendus Boil. 2008, 331, 42–47. [Google Scholar] [CrossRef]
- Partelli, F.L.; Vieira, H.D.; Viana, A.P.; Batista-Santos, P.; Rodrigues, A.P.; Leitao, A.; Ramalho, J.C. Low temperature impact on photosynthetic parameters of coffee genotypes. Pesqui. Agropecu. Bras. 2009, 44, 1404–1415. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef]
- Khan, F.; Hussain, S.; Tanveer, M.; Khan, S.; Hussain, H.A.; Iqbal, B.; Geng, M. Coordinated effects of lead toxicity and nutrient deprivation on growth, oxidative status, and elemental composition of primed and non-primed rice seedlings. Environ. Sci. Pollut. Res. 2018, 25, 21185–21194. [Google Scholar] [CrossRef] [PubMed]
- Kocsy, G.; Von, B.P.; Rüegsegger, A.; Szalai, G.; Galiba, G.; Brunold, C. Increasing the glutathione content in a chilling-sensitive maize genotype using safeners increased protection against chilling-induced injury. Plant Physiol. 2001, 127, 1147–1156. [Google Scholar] [CrossRef]
- Chen, W.; Guo, C.; Hussain, S.; Zhu, B.; Deng, F.; Xue, Y.; Geng, M.; Wu, L. Role of xylo-oligosaccharides in protection against salinity-induced adversities in Chinese cabbage. Environ. Sci. Pollut. Res. 2015, 23, 1254–1264. [Google Scholar] [CrossRef]
- Xing, C.H.; Xu, S.C.; Bo, S.H.; Tao, L.X.; Cao, H.-X.; Sun, C.-X.; Shao, H.-B.; Lei, X.-T. Effects of low temperature and drought on the physiological and growth changes in oil palm seedlings. Afr. J. Biotechnol. 2011, 10, 2630–2637. [Google Scholar] [CrossRef]
- Guo, H.; Gao, S.; Zhao, F.; Li, F. Effects of cold acclimation on several enzyme activities in Euonymus radicans ‘emorald & gold’ and its relation to semi-lethal temperature. For. Stud. China 2004, 6, 10–17. [Google Scholar] [CrossRef]
- Roberts, L.A.; Pierson, A.J.; Panaviene, Z.; Walker, E. Yellow Stripe1. Expanded Roles for the Maize Iron-Phytosiderophore Transporter1. Plant Physiol. 2004, 135, 112–120. [Google Scholar] [CrossRef]
- Peng, Y.S.; Liu, E. A comparative study of methods of extracting chlorophyll. Acta Agric. Universitatis Pekin. 1992, 18, 247–250. [Google Scholar]
- Shan, D.-P.; Huang, J.; Yang, Y.-T.; Guo, Y.-H.; Wu, C.; Yang, G.-D.; Gao, Z.; Zheng, C. Cotton GhDREB1 increases plant tolerance to low temperature and is negatively regulated by gibberellic acid. New Phytol. 2007, 176, 70–81. [Google Scholar] [CrossRef]












© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, H.A.; Men, S.; Hussain, S.; Zhang, Q.; Ashraf, U.; Anjum, S.A.; Ali, I.; Wang, L. Maize Tolerance against Drought and Chilling Stresses Varied with Root Morphology and Antioxidative Defense System. Plants 2020, 9, 720. https://doi.org/10.3390/plants9060720
Hussain HA, Men S, Hussain S, Zhang Q, Ashraf U, Anjum SA, Ali I, Wang L. Maize Tolerance against Drought and Chilling Stresses Varied with Root Morphology and Antioxidative Defense System. Plants. 2020; 9(6):720. https://doi.org/10.3390/plants9060720
Chicago/Turabian StyleHussain, Hafiz Athar, Shengnan Men, Saddam Hussain, Qingwen Zhang, Umair Ashraf, Shakeel Ahmad Anjum, Iftikhar Ali, and Longchang Wang. 2020. "Maize Tolerance against Drought and Chilling Stresses Varied with Root Morphology and Antioxidative Defense System" Plants 9, no. 6: 720. https://doi.org/10.3390/plants9060720
APA StyleHussain, H. A., Men, S., Hussain, S., Zhang, Q., Ashraf, U., Anjum, S. A., Ali, I., & Wang, L. (2020). Maize Tolerance against Drought and Chilling Stresses Varied with Root Morphology and Antioxidative Defense System. Plants, 9(6), 720. https://doi.org/10.3390/plants9060720