Tissue Culture and Refreshment Techniques for Improvement of Transformation in Local Tetraploid and Diploid Potato with Late Blight Resistance as an Example


Abstract
1. Introduction
2. Results and Discussion
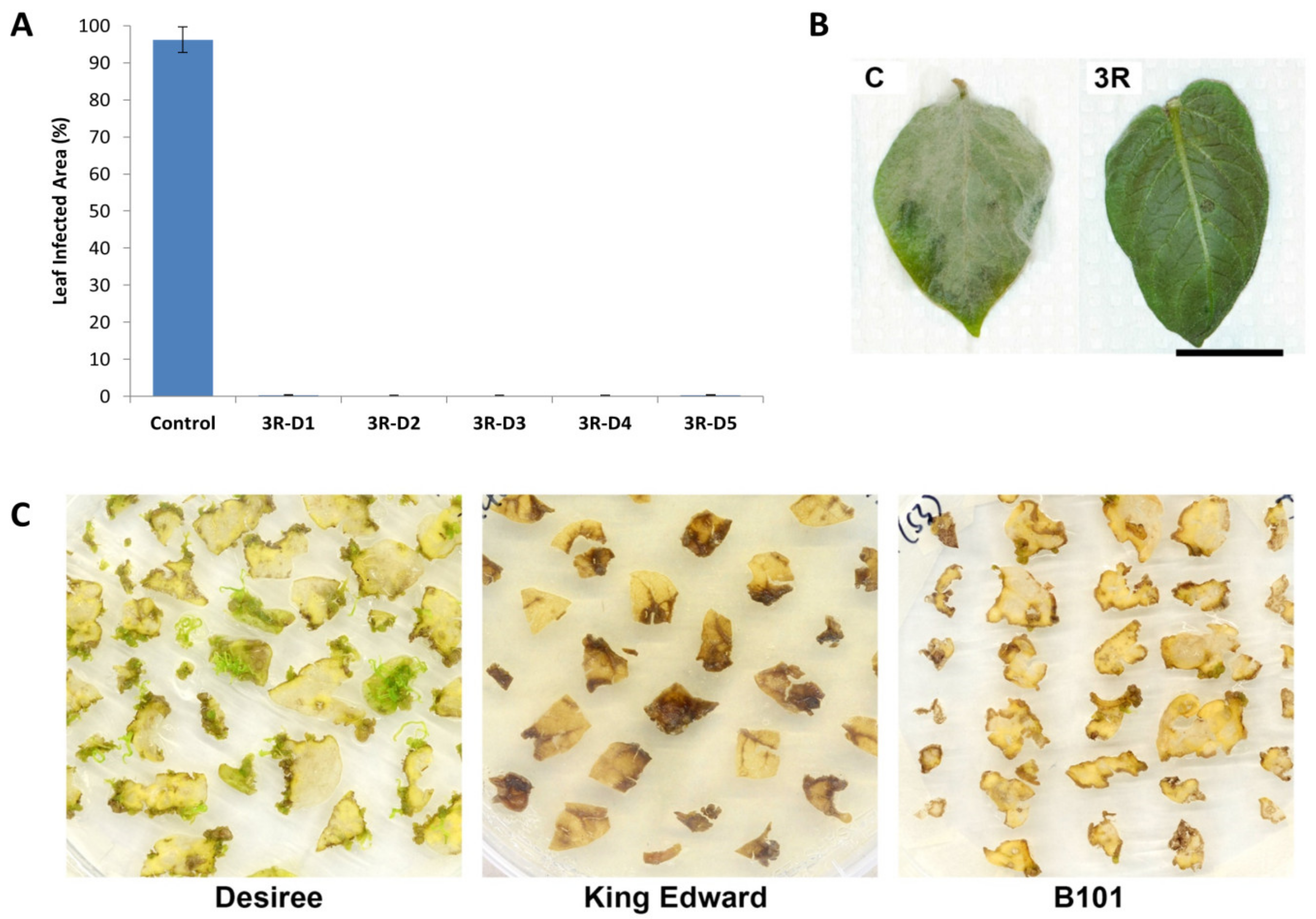
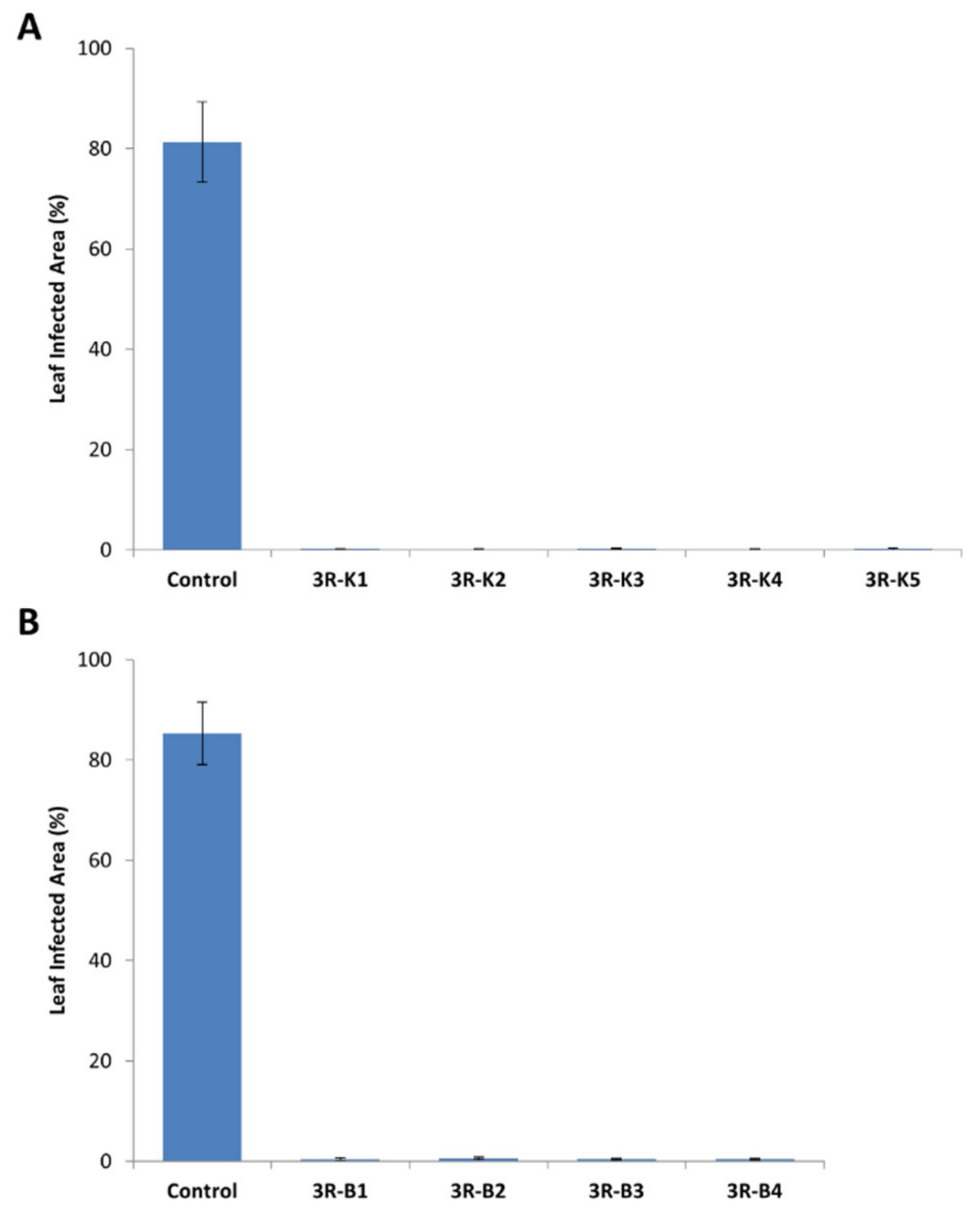
2.1. Introduction of a Triple Resistance Gene Stack in Désirée
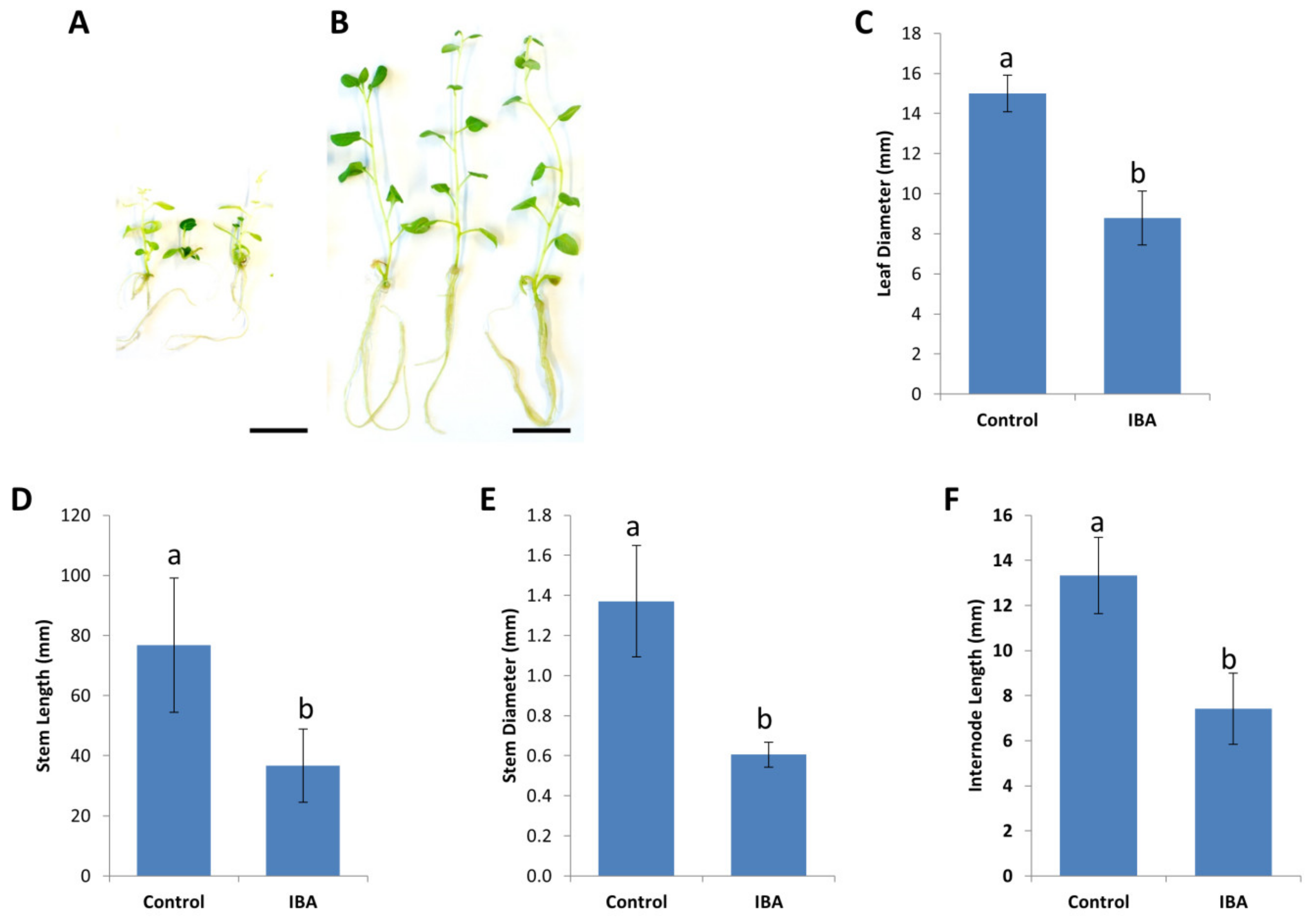
2.2. Long-Term Effects of Plant Growth Regulators on In Vitro Plant Material
2.3. Optimization of Plant Growth Regulators for Shoot Induction from King Edward and B101
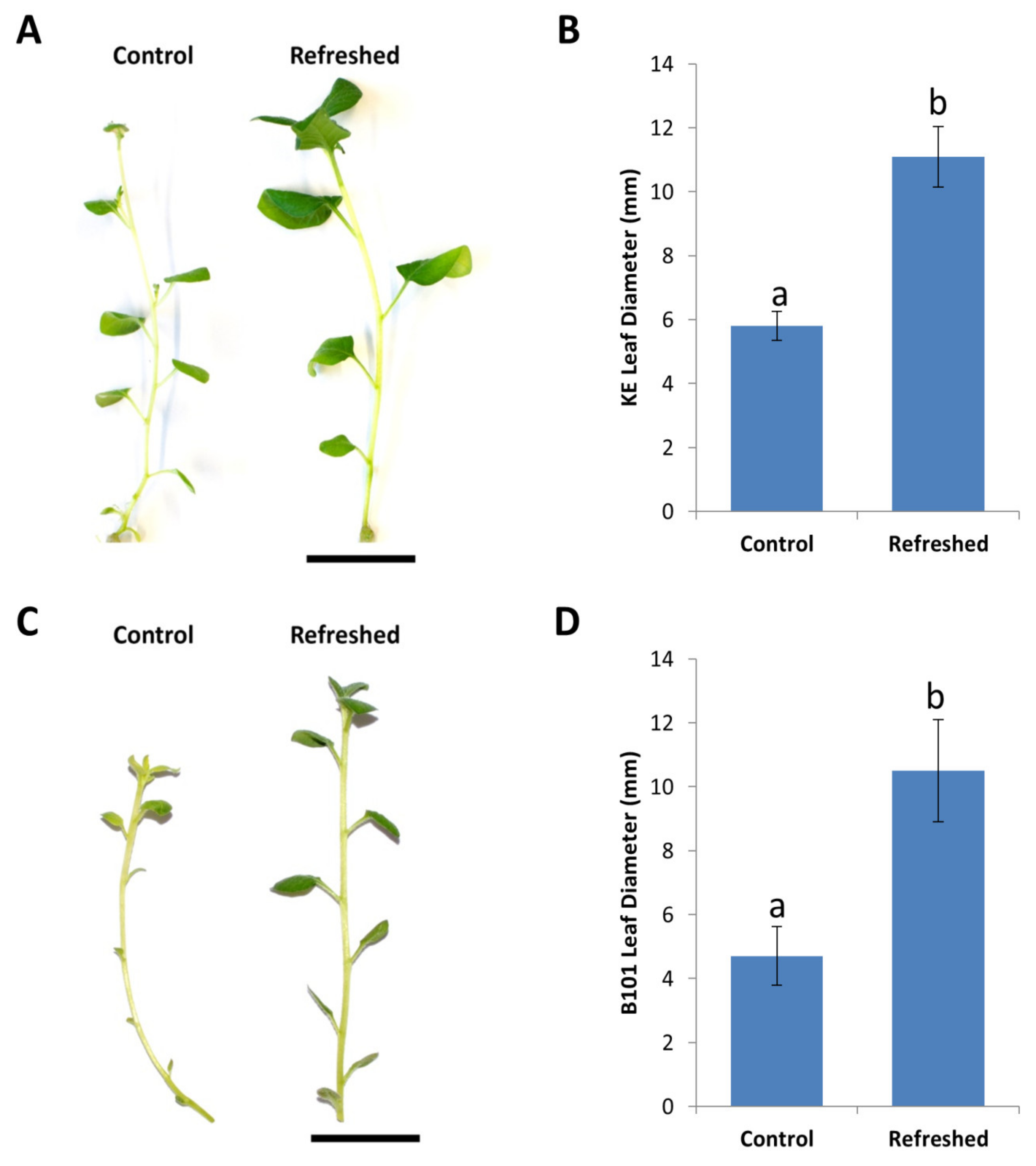
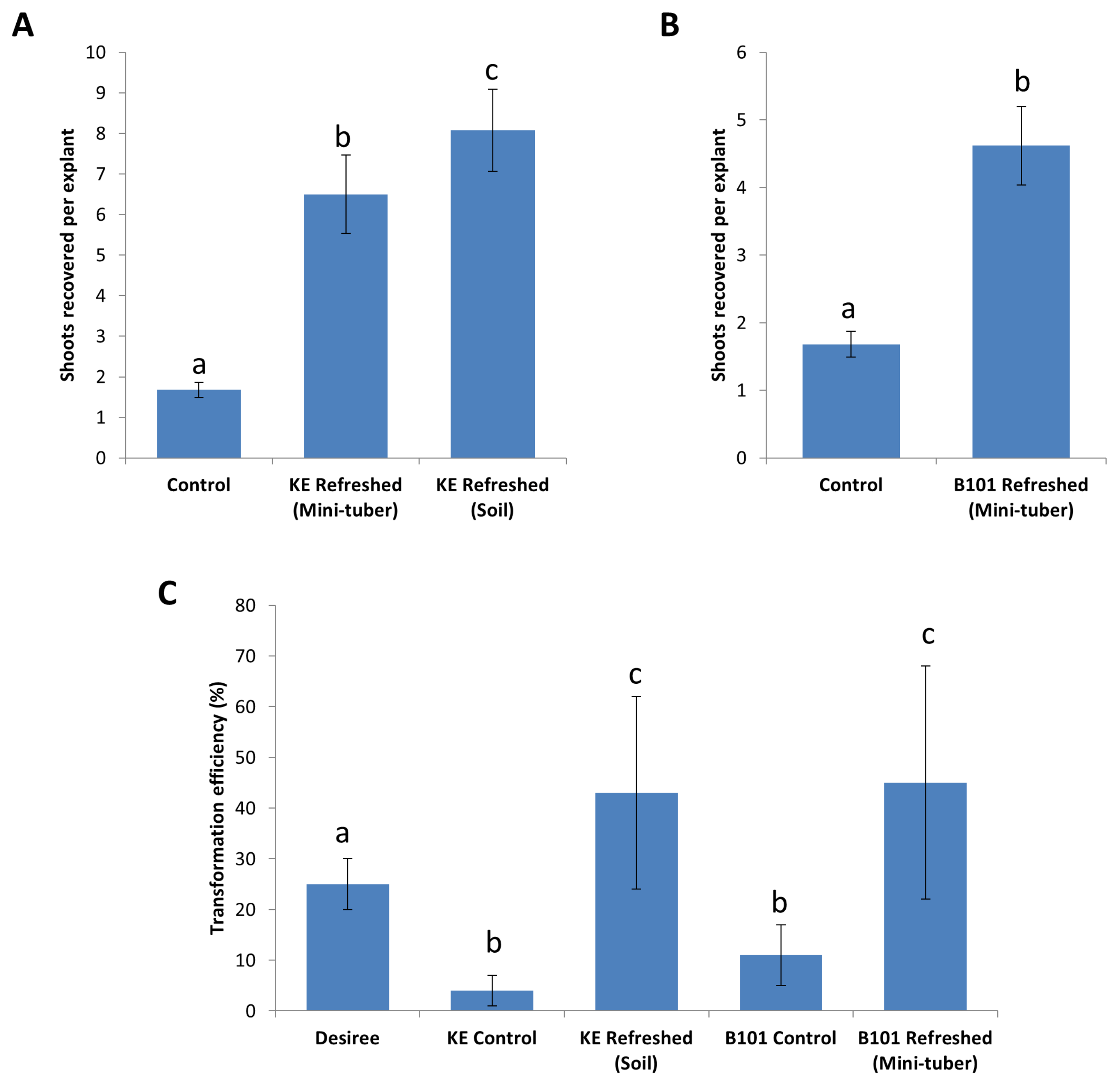
2.4. Improving Regeneration with Refreshed Stock Lines
2.5. Stacking of Resistance Genes in King Edward and B101
3. Materials and Methods
3.1. Materials
3.2. Potato Transformation Protocol
3.3. Molecular Screening
3.4. Maintenance and Refreshment of In Vitro Material
3.5. Detached-Leaf Assay
3.6. Data Collection and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eriksson, D.; Carlson-Nilsson, U.; Ortíz, R.; Andreasson, E. Overview and Breeding Strategies of Table Potato Production in Sweden and the Fennoscandian Region. Potato Res. 2016, 59, 279–294. [Google Scholar] [CrossRef]
- Ristaino, J. Tracking historic migrations of the Irish potato famine pathogen, Phytophthora infestans. Microbes Infect. 2002, 4, 1369–1377. [Google Scholar] [CrossRef]
- Rakotonindraina, T.; Chauvin, J.-É.; Pellé, R.; Faivre, R.; Chatot, C.; Savary, S.; Aubertot, J.-N. Modeling of Yield Losses Caused by Potato Late Blight on Eight Cultivars with Different Levels of Resistance to Phytophthora infestans. Plant Dis. 2012, 96, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Helgeson, J.P.; Pohlman, J.D.; Austin, S.; Haberlach, G.T.; Wielgus, S.M.; Ronis, D.; Zambolim, L.; Tooley, P.; McGrath, J.M.; James, R.V.; et al. Somatic hybrids between Solanum bulbocastanum and potato: a new source of resistance to late blight. Theor. Appl. Genet. 1998, 96, 738–742. [Google Scholar] [CrossRef]
- Jansky, S.H. Overcoming hybridization barriers in potato. Plant Breed. 2006, 125, 1–12. [Google Scholar] [CrossRef]
- Lindhout, P.; De Vries, M.; Ter Maat, M.; Ying, S.; Viquez-Zamora, M.; Van Heusden, S.; Solynta, T.N. Hybrid potato breeding for improved varieties. In Biopesticides for Sustainable Agriculture; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2018; Volume 1, pp. 99–122. [Google Scholar]
- Jo, K.-R.; Kim, C.-J.; Kim, S.-J.; Kim, T.-Y.; Bergervoet, M.; Jongsma, M.A.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Development of late blight resistant potatoes by cisgene stacking. BMC Biotechnol. 2014, 14, 50. [Google Scholar] [CrossRef]
- Haesaert, G.; Vossen, J.H.; Custers, R.; De Loose, M.; Haverkort, A.; Heremans, B.; Hutten, R.; Kessel, G.; Landschoot, S.; Van Droogenbroeck, B.; et al. Transformation of the potato variety Desiree with single or multiple resistance genes increases resistance to late blight under field conditions. Crop. Prot. 2015, 77, 163–175. [Google Scholar] [CrossRef]
- Ghislain, M.; Byarugaba, A.A.; Magembe, E.; Njoroge, A.; Rivera, C.; Román, M.L.; Tovar, J.C.; Gamboa, S.; Forbes, G.A.; Kreuze, J.F.; et al. Stacking three late blight resistance genes from wild species directly into African highland potato varieties confers complete field resistance to local blight races. Plant Biotechnol. J. 2018, 17, 1119–1129. [Google Scholar] [CrossRef]
- Visser, R.G.F. Regeneration and transformation of potato by Agrobacterium tumefaciens. In Plant Tissue Culture Manual; Springer: Berlin/Heidelberg, Germany, 1991; Volume B5, pp. 301–309. [Google Scholar]
- Sun, K.; Wolters, A.-M.A.; Vossen, J.H.; Rouwet, M.E.; Loonen, A.E.H.M.; Jacobsen, E.; Visser, R.G.F.; Bai, Y. Silencing of six susceptibility genes results in potato late blight resistance. Transgenic Res. 2016, 25, 731–742. [Google Scholar] [CrossRef]
- Birch, R.G. PLANT TRANSFORMATION: Problems and Strategies for Practical Application. Annu. Rev. Plant Boil. 1997, 48, 297–326. [Google Scholar] [CrossRef]
- Smýkal, P.; Valledor, L.; Rodriguez, R.; Griga, M. Assessment of genetic and epigenetic stability in long-term in vitro shoot culture of pea (Pisum sativum L.). Plant Cell Rep. 2007, 26, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Us-Camas, R.; Rivera-Solis, G.; Duarte-Aké, F.; De-La-Peña, C. In vitro culture: an epigenetic challenge for plants. Plant Cell Tissue Organ Cult. (PCTOC) 2014, 118, 187–201. [Google Scholar] [CrossRef]
- Baránek, M.; Čechová, J.; Raddová, J.; Holleinová, V.; Ondrušíková, E.; Pidra, M. Dynamics and Reversibility of the DNA Methylation Landscape of Grapevine Plants (Vitis vinifera) Stressed by In Vitro Cultivation and Thermotherapy. PLoS ONE 2015, 10, e0126638. [Google Scholar] [CrossRef] [PubMed]
- Han, E.-H.; Goo, Y.-M.; Lee, M.-K.; Lee, S.-W. An efficient transformation method for a potato (Solanum tuberosum L. var. Atlantic). J. Plant Biotechnol. 2015, 42, 77–82. [Google Scholar] [CrossRef]
- Woodward, A.; Bartel, B. Auxin: Regulation, Action, and Interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, V.; Mallubhotla, S.; MacDonnell, J.; Sanyal-Bagchi, M.; Chakravarty, B.; Wang-Pruski, G.; Rothwell, C.; Audy, P.; De Koeyer, D.; Siahbazi, M.; et al. Transformation and Plant Regeneration from Leaf Explants of Solanum tuberosum L. cv. ‘Shepody’. Plant Cell Tissue Organ Cult. (PCTOC) 2006, 85, 361–366. [Google Scholar] [CrossRef]
- Yee, S.; Stevens, B.; Coleman, S.; Seabrook, J.E.A.; Li, X. High-efficiency regnerationin vitro from potato petioles with intact leaflets. Am. J. Potato Res. 2001, 78, 151–157. [Google Scholar] [CrossRef]
- Gupta, R.; Chakrabarty, S.K. Gibberellic acid in plant. Plant Signal. Behav. 2013, 8, e25504. [Google Scholar] [CrossRef]
- Hedden, P.; Sponsel, V. A Century of Gibberellin Research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, P.; Tang, D.; Yang, Z.; Lu, F.; Qi, J.; Tawari, N.R.; Shang, Y.; Li, C.; Huang, S. The genetic basis of inbreeding depression in potato. Nat. Genet. 2019, 51, 374–378. [Google Scholar] [CrossRef]
- Hodgson, W.A.; Grainger, P.N. Culture Of Phytophthora Infestans On Artificial Media Prepared From Rye Seeds. Can. J. Plant Sci. 1964, 44, 583. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef] [PubMed]
- Burra, D.D.; Berkowitz, O.; E Hedley, P.; Morris, J.; Resjö, S.; Levander, F.; Liljeroth, E.; Andréasson, E.; Alexandersson, E. Phosphite-induced changes of the transcriptome and secretome in Solanum tuberosum leading to resistance against Phytophthora infestans. BMC Plant Boil. 2014, 14, 254. [Google Scholar] [CrossRef] [PubMed]
- Kieu, N.P.; Lenman, M.; Andreasson, E. Potato as a Model for Field Trials with Modified Gene Functions in Research and Translational Experiments in “Solanum Tuberosum: Methods and Protocols”. In Methods in Molecular Biology; Dobnik, D., Ramšak, Ž., Coll, A., Gruden, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Bengtsson, T.; Weighill, D.; Proux-Wéra, E.; Levander, F.; Resjö, S.; Burra, D.D.; Moushib, L.I.; E Hedley, P.; Liljeroth, E.; Jacobson, D.; et al. Proteomics and transcriptomics of the BABA-induced resistance response in potato using a novel functional annotation approach. BMC Genom. 2014, 15, 315. [Google Scholar] [CrossRef] [PubMed]
- Goth, R.W.; Keane, J. A detached-leaf method to evaluate late blight resistance in potato and tomato. Am. J. Potato Res. 1997, 74, 347–352. [Google Scholar] [CrossRef]
- Moushib, L.I.; Witzell, J.; Lenman, M.; Liljeroth, E.; Andreasson, E. Sugar beet extract induces defence against Phytophthora infestans in potato plants. Eur. J. Plant Pathol. 2013, 136, 261–271. [Google Scholar] [CrossRef]
- Abreha, K.B.; Alexandersson, E.; Vossen, J.H.; Anderson, P.; Andreasson, E. Inoculation of Transgenic Resistant Potato by Phytophthora infestans Affects Host Plant Choice of a Generalist Moth. PLOS ONE 2015, 10, e0129815. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Material | ||||
|---|---|---|---|---|
| Components | Désirée | King Edward | B101 | Use |
| MS1 | Maintenance of in vitro lines, rooting media, stock propagation | |||
| MS salts + Vitamins | 4.4 g/L | 4.4 g/L | 4.4 g/L | |
| Sucrose | 10 g/L | 10 g/L | 10 g/L | |
| pH | 5.8 | 5.8 | 5.8 | |
| Phytoagar | 8 g/L | 8 g/L | 8 g/L | |
| CIM (MS1 including compounds below) | Co-cultivation post transformation, callus induction | |||
| BAP | 2.0 mg/L | 2.0 mg/L | 2.0 mg/L | |
| trans-Zeatin-riboside | - | - | - | |
| NAA | 0.2 mg/L | 0.2 mg/L | 0.2 mg/L | |
| GA3 | - | - | - | |
| SIM (MS1 including compounds below) | Selection and regeneration post transformation, shoot induction | |||
| BAP | - | - | - | |
| trans-Zeatin-riboside | 2.0 mg/L | 4.0 mg/L | 2.0 mg/L | |
| NAA | 0.01 mg/L | 0.01 mg/L | - | |
| GA3 | 0.1 mg/L | 0.1 mg/L | 2.5 mg/L | |
| Kanamycin | 100 mg/L | 100 mg/L | 100 mg/L | |
| Claforan (cefotaxime) | 400 mg/L | 400 mg/L | 400 mg/L | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, E.S.; Kieu, N.P.; Lenman, M.; Andreasson, E. Tissue Culture and Refreshment Techniques for Improvement of Transformation in Local Tetraploid and Diploid Potato with Late Blight Resistance as an Example. Plants 2020, 9, 695. https://doi.org/10.3390/plants9060695
Wang ES, Kieu NP, Lenman M, Andreasson E. Tissue Culture and Refreshment Techniques for Improvement of Transformation in Local Tetraploid and Diploid Potato with Late Blight Resistance as an Example. Plants. 2020; 9(6):695. https://doi.org/10.3390/plants9060695
Chicago/Turabian StyleWang, Eu Sheng, Nam Phuong Kieu, Marit Lenman, and Erik Andreasson. 2020. "Tissue Culture and Refreshment Techniques for Improvement of Transformation in Local Tetraploid and Diploid Potato with Late Blight Resistance as an Example" Plants 9, no. 6: 695. https://doi.org/10.3390/plants9060695
APA StyleWang, E. S., Kieu, N. P., Lenman, M., & Andreasson, E. (2020). Tissue Culture and Refreshment Techniques for Improvement of Transformation in Local Tetraploid and Diploid Potato with Late Blight Resistance as an Example. Plants, 9(6), 695. https://doi.org/10.3390/plants9060695