Taxonomic Implications of Leaf Micromorphology Using Microscopic Analysis: A Tool for Identification and Authentication of Korean Piperales



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
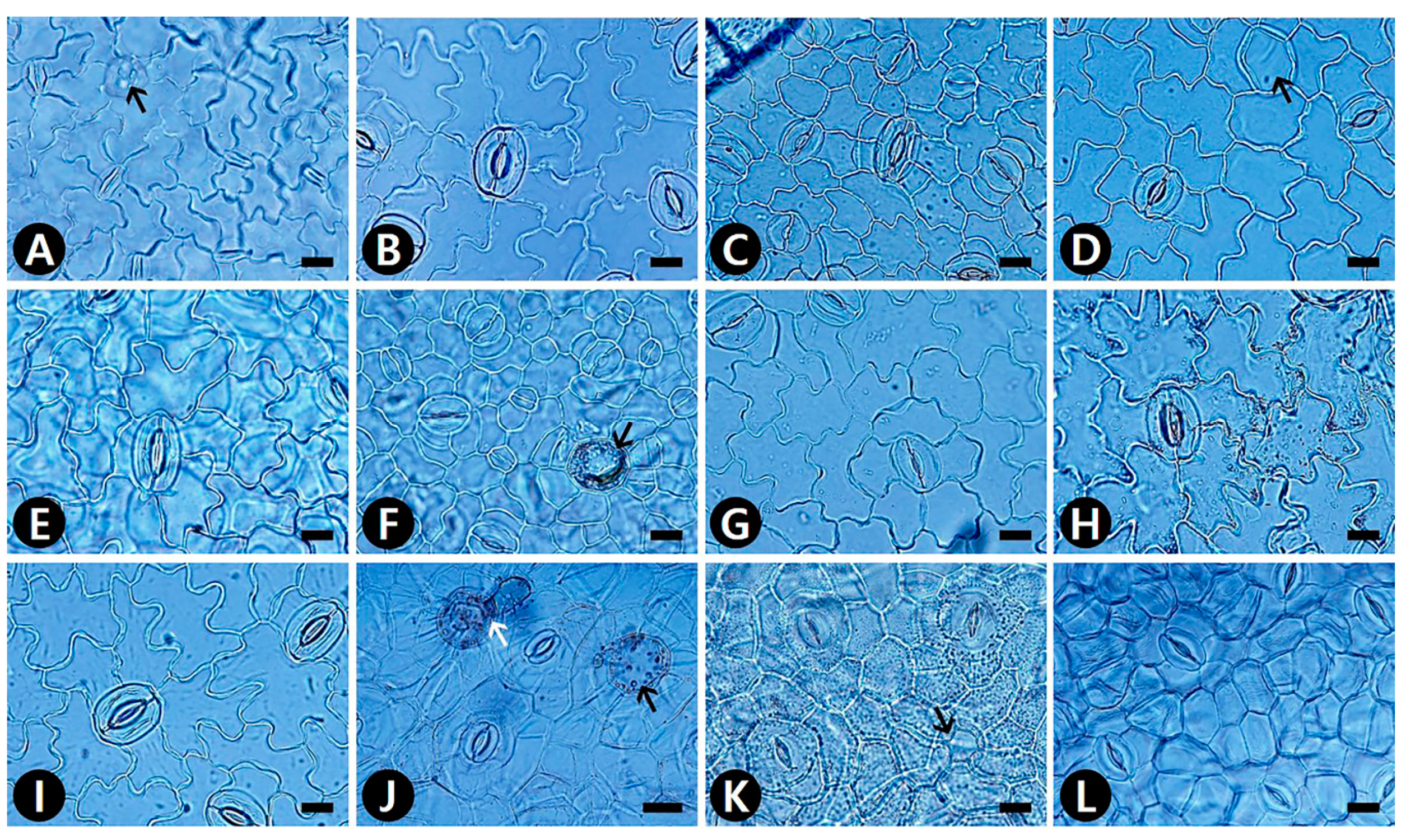
2.1. Epidermal Cells
2.2. Idioblast Cells
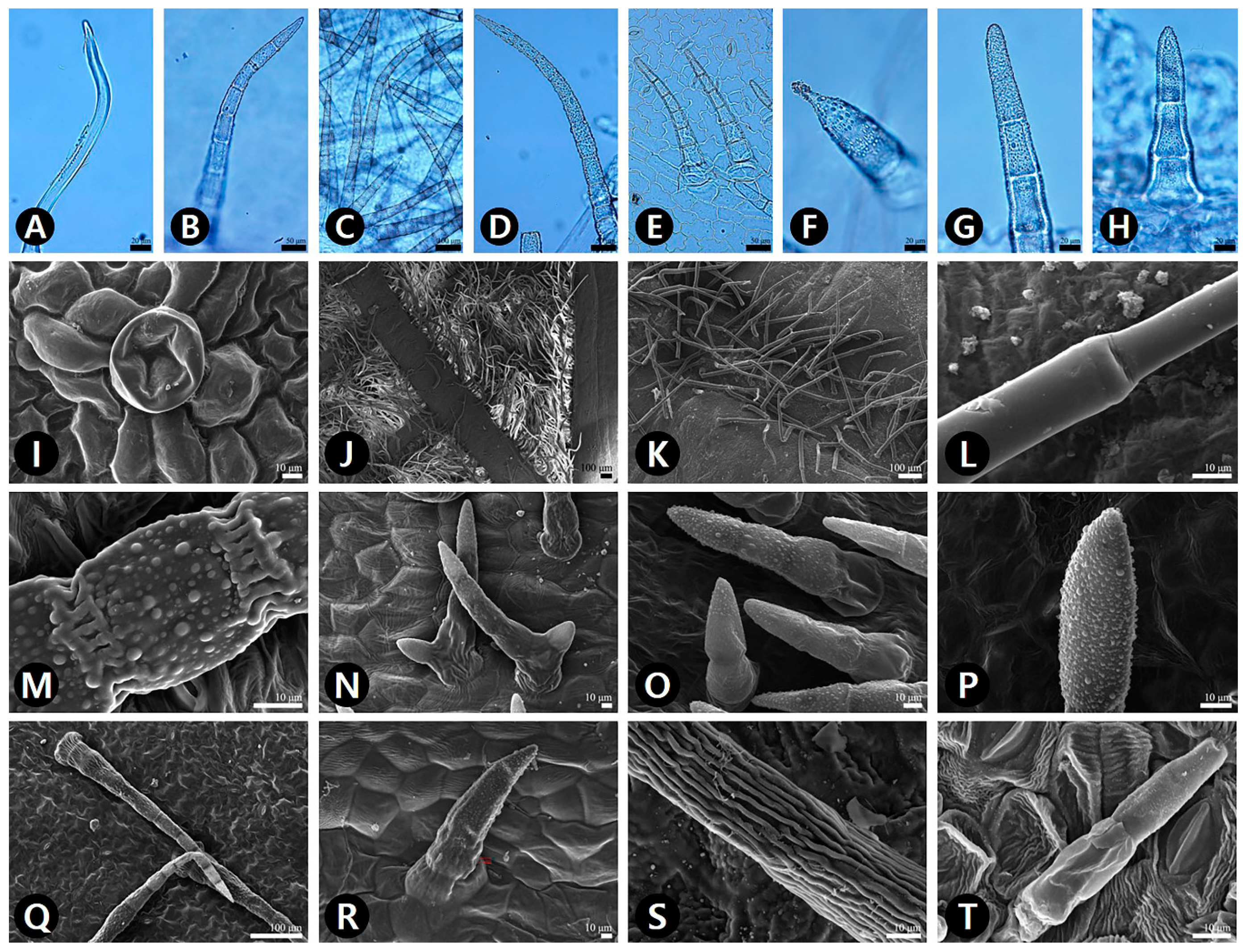
2.3. Trichomes
2.4. Stomatal Complex
2.5. Identification Key Based on Leaf Micromorphology
| 1. Stomatal position amphistomatic ........................................................................................................... 2 1’. Stomatal position hypostomatic ............................................................................................................ 3 2. Stomatal ledge aperture polar rods to the guard cells; stomatal pore fusiform slit ..................................................................................................................................... Houttuynia cordata 2’. Stomatal ledge narrowly elliptical; stomatal pore elliptic slit ........................................................................................................................................................................ 4 3. Stomatal ledge aperture polar rods to the guard cells; stomatal pore fusiform slit; stomatal surface striae extended as lateral wing or radiating striae ..................................................................... 5 3’. Stomatal ledge aperture narrowly elliptical; stomatal pore elliptic slit; stomatal surface not striated ........................................................................................................................................................... 6 4. Presence of both glandular trichomes and verrucate multicellular non-glandular trichomes on the abaxial surface .......................................... Asarum heterotropoides vars. mandshuricum and seoulense 4’. Presence of verrucate multicellular non-glandular trichomes only on the abaxial surface ............................................................................................................................................................... 7 5. Stomatal surface striae extended as lateral wing; stomatal types anisocytic and tetracytic; guard cells concentric rings; secretory idioblasts protruding ........................................................ Piper kadsura 5’. Stomatal surface radiating striae; stomatal type actinocytic; guard cells smooth; secretory idioblasts flat to central tubercle .................................................................................... Saururus chinensis 6. Fine relief of the epidermal cells smooth; sparsely pubescent with glandular trichomes on the abaxial surface; stomatal ledge aperture thick and wide outer stomatal ledge with linear slit pore ................................................................................................................................. Aristolochia contorta 6’. Fine relief of the epidermal cells striate and wrinkled; densely pubescent with verrucate multicellular non-glandular trichomes on the abaxial surface; stomatal ledge aperture narrowly elliptical with elliptic slit pore ........................................................................... Aristolochia manshuriensis 7. Stomatal types anomocytic, staurocytic, and tetracytic ...................................................................... 8 7’. Stomatal type anomocytic only ............................................................................................................. 9 8. Secretory idioblasts flat to convex; presence of verrucate Y-shaped multicellular non-glandular trichomes ........................................................................................................................... Asarum koreanum 8’. Secretory idioblasts flat to central tubercle; absence of verrucate Y-shaped multicellular non- glandular trichomes ................................................................................................................................... 10 9. Undulous anticlinal cell walls on the adaxial surface; fine relief of the epidermal cells on the abaxial surface smooth .................................................................................................. Asarum maculatum 9’. Straight to curved anticlinal cell walls on the adaxial surface; fine relief of the epidermal cells on the abaxial surface striate and wrinkled ............................................................................................ 11 10. Straight to curved anticlinal cell walls on the abaxial surface; sparsely pubescent with verrucate multicellular non-glandular trichomes on the abaxial surface ................................ Asarum misandrum 10’. Sinuous anticlinal cell walls on the abaxial surface; densely pubescent with verrucate multicellular non-glandular trichomes on the abaxial surface ........................................ Asarum patens 11. Irregular epidermal cell arrangement and presence of pentagonal secretory idioblasts on the adaxial surface; sparsely pubescent with verrucate multicellular non-glandular trichomes and presence of double semi-circular stomatal ledge on the abaxial surface ................................................................................................................................ Asarum versicolor 11’. Isodiametric epidermal cell arrangement and absence of pentagonal secretory idioblasts on the adaxial surface; moderate to densely pubescent with verrucate multicellular non-glandular trichomes and only presence of lip-shaped stomatal ledge on the abaxial surface ................................................................................................................................... Asarum sieboldii |
3. Discussion
3.1. Leaf Micromorphology and Its Taxonomic Implication in Korean Piperales
3.2. Leaf Micromorphology for Taxonomy of Korean Asarum
3.3. Application for Authentication of Herbal Medicine
4. Materials and Methods
4.1. Taxon Sampling and Identification
4.2. Light Microscopic Analysis
4.3. Scanning Electron Microscopic Analysis
4.4. Quantitative Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Stace, C.A. The taxonomic importance of the leaf surface. In Current Concepts in Plant Taxonomy; Academic Press: London, UK, 1984; Volume 25, pp. 67–93. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons, 2nd ed.; Clarendon Press: Oxford, UK, 1979; pp. 456–473. [Google Scholar]
- Câmara, P.E.; Kellogg, E.A. Morphology and development of leaf papillae in Sematophyllaceae. Bryologist 2010, 113, 22–34. [Google Scholar] [CrossRef]
- Khan, A.S.; Ahmad, M.; Zafar, M.; Athar, M.; Ozdemir, F.A.; Gilani, S.A.A.; Sultana, S.; Ahmad, S.; Butt, M.A.; Majeed, S.; et al. Morphological characterization of Hypnaceae (Bryopsida, Hypnales): Investigating four genera from Western Himalayas by using LM and SEM techniques. Microsc. Res. Tech. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.N.; Ahmad, M.; Zafar, M.; Malik, K.; Rashid, N.; Ullah, F.; Zaman, W.; Ali, M. A light and scanning electron microscopic diagnosis of leaf epidermal morphology and its systematic implications in Dryopteridaceae: Investigating 12 Pakistani taxa. Micron 2018, 111, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.N.; Ahmad, M.; Zafar, M.; Razzaq, A.; Malik, K.; Rashid, N.; Ullah, F.; Iqbal, M.; Zaman, W. Foliar epidermal micromorphology and its taxonomic implications in some selected species of Athyriaceae. Microsc. Res. Tech. 2018, 81, 902–913. [Google Scholar] [CrossRef]
- Shah, S.N.; Celik, A.; Ahmad, M.; Ullah, F.; Zaman, W.; Zafar, M.; Malik, K.; Rashid, N.; Iqbal, M.; Sohail, A.; et al. Leaf epidermal micromorphology and its implications in systematics of certain taxa of the fern family Pteridaceae from northern Pakistan. Microsc. Res. Techniq. 2019, 82, 317–332. [Google Scholar] [CrossRef]
- Clugston, J.A.R.; Jeffree, C.E.; Ahrends, A.; Mill, R.R. Do environmental factors affect the taxonomic reliability of leaf cuticular micromorphological characters? A case study in Podocarpaceae. Edinb. J. Bot. 2017, 74, 299–343. [Google Scholar] [CrossRef]
- Elpe, C.; Knopf, P.; Stützel, T.; Schulz, C. Cuticle micromorphology and the evolution of characters in leaves of Taxaceae sl. Bot. J. Linn. Soc. 2017, 184, 503–517. [Google Scholar] [CrossRef]
- Khan, R.; Abidin, S.Z.U.; Ahmad, M.; Zafar, M.; Liu, J.; Jamshed, S.; Kiliç, Ö. Taxonomic importance of SEM and LM foliar epidermal micro-morphology: A tool for robust identification of gymnosperms. Flora 2019, 255, 42–68. [Google Scholar] [CrossRef]
- Woodenberg, W.; Govender, J.; Murugan, N.; Ramdhani, S. Cycad forensics: Leaflet micromorphology as a taxonomic tool for South African cycads. Plant Syst. Evol. 2019, 305, 445–457. [Google Scholar] [CrossRef]
- Behnke, H.D.; Barthlott, W. New evidence from the ultrastructural and micromorphological fields in angiosperm classification. Nord. J. Bot. 1983, 3, 43–66. [Google Scholar] [CrossRef]
- Carpenter, K.J. Stomatal architecture and evolution in basal angiosperms. Am. J. Bot. 2005, 92, 1595–1615. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.K.; Hong, S.P.; Smets, E.; Huysmans, S. Phylogenetic significance of leaf micromorphology and anatomy in the tribe Mentheae (Nepetoideae: Lamiaceae). Bot. J. Linn. Soc. 2009, 160, 211–231. [Google Scholar] [CrossRef]
- Lima, J.F.; Leite, K.R.B.; Clark, L.G.; de Oliveira, R.P. Leaf micromorphology in Poaceae subtribe Olyrinae (Bambusoideae) and its systematic implications. Bot. J. Linn. Soc. 2020, 192, 184–207. [Google Scholar] [CrossRef]
- Da Silva, G.; Gomes, E.; Serrano, R.; Silva, O. Authentication of Euclea natalensis leaf by botanical identification. Planta Med. 2010, 76, P017. [Google Scholar] [CrossRef]
- Da Silva, G.; Serrano, R.; Gomes, E.T.; Silva, O. Botanical features for identification of Gymnosporia arenicola dried leaf. Microsc. Res. Tech. 2015, 78, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Kahkeshani, N.; Khanavi, M.; Hadjiakhoondi, A. quality control of Nepeta menthoides Boiss & Buhse using micromorphological analysis and phytochemical screening. J. Med. Plant Res. 2014, 13, 34–38. [Google Scholar]
- Otang, W.M.; Grierson, D.S.; Ndip, R. Foliar micro-morphology of Gasteria bicolour haw. (Asphodelaceae) from South Africa. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 71–76. [Google Scholar] [CrossRef][Green Version]
- Zhao, Y.; Han, B.; Peng, H.; Wang, X.; Chu, S.; Dai, J.; Peng, D. Identification of “Huoshan shihu” Fengdou: Comparative authentication of the Daodi herb Dendrobium huoshanense and its related species by macroscopic and microscopic features. Microsc. Res. Tech. 2017, 80, 712–721. [Google Scholar] [CrossRef]
- Rashid, S.; Zafar, M.; Ahmad, M.; Shinwari, M.I.; Yaseen, G.; Sultana, S.; Memon, R.A.; Zehra, S.A.; Wibawa, P.A.H. Authentication of herbal drug Tukhm-e-balango (Lallemantia royleana Benth.) using microscopic, pharmacognostic, and phytochemical characterization. Microsc. Res. Tech. 2019, 82, 731–740. [Google Scholar] [CrossRef]
- APG [=Angiosperm Phylogeny Group] IV. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Isnard, S.; Prosperi, J.; Wanke, S.; Wagner, S.T.; Samain, M.S.; Trueba, S.; Frenzke, L.; Neinhuis, C.; Rowe, N.P. Growth form evolution in Piperales and its relevance for understanding angiosperm diversification: An integrative approach combining plant architecture, anatomy, and biomechanics. Int. J. Plant Sci. 2012, 173, 610–639. [Google Scholar] [CrossRef]
- Wiart, C. Medicinal Plants of Asia and the Pacific; CRC Press: Boca Raton, FL, USA, 2006; ISBN 978-0-8493-7245-2. [Google Scholar]
- Wiart, C. Medicinal Plants of China, Korea, and Japan: Bioresources for Tomorrow’s Drugs and Cosmetics; CRC Press: Boca Raton, FL, USA, 2012; ISBN 9781439899113. [Google Scholar]
- Simpson, M.G. Plant. Systematics, 3rd ed.; Academic Press: San Diego, CA, USA, 2019; ISBN 9780128126288. [Google Scholar]
- Zhang, H.M.; Zhao, X.H.; Sun, Z.H.; Li, G.C.; Liu, G.C.; Sun, L.R.; Hou, J.Q.; Zhou, W. Recognition of the toxicity of aristolochic acid. J. Clin. Pharm. Ther. 2019, 44, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Pei, J.; Poon, J.; Lau, A.Y.; Zhang, L.; Wang, Y.; Liu, C.; Huang, L. Worldwide research trends on aristolochic acids (1957–2017): Suggestions for researchers. PLoS ONE 2019, 14, e0216135. [Google Scholar] [CrossRef] [PubMed]
- Flora of Korea Editorial Committee. The Genera of Vascular Plants of Flora; Hongreung Publishing Co.: Seoul, Korea, 2018; ISBN 979-11-5600-612-1. [Google Scholar]
- Korea Institute of Oriental Medicine. Defining Dictionary for Medicinal Herbs. 2020. Available online: http://https://oasis.kiom.re.kr/herblib/hminfo/hbmcod/hbmcodList.do (accessed on 21 February 2020).
- Korea Food and Drug Administration. The Korean Pharmacophoeia 2, 11th ed.; Korea Food and Drug Administration: Seoul, Korea, 2014; pp. 57–58, 205–207, 274–276, 444–445.
- Lee, T.B. Illustrated Flora of Korea; Hyangmunsa: Seoul, Korea, 1980. [Google Scholar]
- Lee, T.B. Colored Flora of Korea; Hyangmunsa: Seoul, Korea, 2003; Volume I. [Google Scholar]
- Lee, W.T. Standard Illustrations of Korean Plants; Academy Publishing Co.: Seoul, Korea, 1996. [Google Scholar]
- So, S.; Kim, M. A taxonomic study of Asarum (Aristolochiaceae) in Korea. Korean J. Plant Taxon. 2008, 38, 121–149. [Google Scholar] [CrossRef]
- Oh, B.U. Asarum. In Flora of Korea Volume 2a Magnoliidae; Flora of Korea Editorial Committee and the National Institute of Biological Resources: Incheon, Korea, 2017; pp. 20–22. ISBN 978-89-6811-275-1. [Google Scholar]
- Oh, B.U. A taxonomic review of Korean Asarum (Aristolochiaceae). Korean J. Plant Taxon. 2008, 38, 251–270. [Google Scholar] [CrossRef]
- Kim, Y.S.; Park, H.D.; Kim, E.C. Taxonomic study of the Genus Asarum in Korea. Korean J. Plant Taxon. 1978, 8, 19–31. [Google Scholar] [CrossRef]
- Lee, B.R.; Kim, S.H.; Huh, M.K. Phylogenic study of genus Asarum (Aristolochiaceae) in Korea by trnL-trnT region. J. Life Sci. 2010, 20, 1697–1703. [Google Scholar] [CrossRef]
- So, S.; Kim, M. Genetic variation and population structure of Asarum misandrum (Aristolochiaceae) in Korea. Korean J. Plant Taxon. 2013, 43, 181–187. [Google Scholar] [CrossRef][Green Version]
- Leem, H.; Oh, B.U. Taxonomy of Korean Asarum (Aristolochiaceae) by the morphological characters. Korean J. Plant Res. 2019, 32, 344–354. [Google Scholar] [CrossRef]
- Garcia Alvarez, S.; Garcia-Amorena, I.; Rubiales, J.M.; Morla, C. The value of leaf cuticle characteristics in the identification and classification of Iberian Mediterranean members of the genus Pinus. Bot. J. Linn. Soc. 2009, 161, 436–448. [Google Scholar] [CrossRef]
- Grace, O.M.; Simmonds, M.S.; Smith, G.F.; Van Wyk, A.E. Taxonomic significance of leaf surface morphology in Aloe section Pictae (Xanthorrhoeaceae). Bot. J. Linn. Soc. 2009, 160, 418–428. [Google Scholar] [CrossRef][Green Version]
- Yang, Y.; Zhang, L.Y.; Liu, B.; van der Werff, H. Leaf cuticular anatomy and taxonomy of Syndiclis (Lauraceae) and its allies. Syst. Bot. 2012, 37, 861–878. [Google Scholar] [CrossRef]
- Nisa, S.U.; Shah, S.A.; Mumtaz, A.S.; Sultan, A. Stomatal novelties in Vincetoxicum arnottianum (Asclepiadeae: Asclepiadoideae: Apocynaceae). Flora 2019, 260, 151464. [Google Scholar] [CrossRef]
- Webster, G.L.; Del-Arco-Aguilar, M.J.; Smith, B.A. Systematic distribution of foliar trichome types in Croton (Euphorbiaceae). Bot. J. Linn. Soc. 1996, 121, 41–57. [Google Scholar] [CrossRef]
- Hu, G.X.; Balangcod, T.; Xiang, C.L. Trichome micromorphology of the Chinese-Himalayan genus Colquhounia (Lamiaceae), with emphasis on taxonomic implications. Biologia 2012, 67, 867–874. [Google Scholar] [CrossRef]
- Eiji, S.; Salmaki, Y. Evolution of trichomes and its systematic significance in Salvia (Mentheae; Nepetoideae; Lamiaceae). Bot. J. Linn. Soc. 2016, 180, 241–257. [Google Scholar] [CrossRef]
- Mannethody, S.; Purayidathkandy, S. Trichome micromorphology and its systematic significance in Asian Leucas (Lamiaceae). Flora 2018, 242, 70–78. [Google Scholar] [CrossRef]
- Ashfaq, S.; Ahmad, M.; Zafar, M.; Sultana, S.; Bahadur, S.; Ullah, F.; Zaman, W.; Ahmed, S.N.; Nazish, M. Foliar micromorphology of Convolvulaceous species with special emphasis on trichome diversity from the arid zone of Pakistan. Flora 2019, 255, 110–124. [Google Scholar] [CrossRef]
- Barthlott, W.; Neinhuis, C.; Cutler, D.; Ditsch, F.; Meusel, I.; Theisen, I.; Wilhelmi, H. Classification and terminology of plant epicuticular waxes. Bot. J. Linn. Soc. 1998, 126, 237–260. [Google Scholar] [CrossRef]
- Wissemann, V. Epicuticular wax morphology and the taxonomy of Rosa (section Caninae, subsection Rubiginosae). Plant Syst. Evol. 2000, 221, 107–112. [Google Scholar] [CrossRef]
- Tomaszewski, D.; Zieliński, J. Epicuticular wax structures on stems and comparison between stems and leaves–A survey. Flora 2014, 209, 215–232. [Google Scholar] [CrossRef]
- Tomaszewski, D.; Byalt, A.; Gawlak, M. Leaf and stem epicuticular wax structures in Lonicera species (Caprifoliaceae). Nord. J. Bot. 2019, 37, e02210. [Google Scholar] [CrossRef]
- Marinho, C.R.; Zacaro, A.A.; Ventrella, M.C. Secretory cells in Piper umbellatum (Piperaceae) leaves: A new example for the development of idioblasts. Flora 2011, 206, 1052–1062. [Google Scholar] [CrossRef]
- Kakoli, D.; Pratik, R. Studies on micromorphology of leaves in some members of genus piper linn. Int. J. Life Sci. 2015, 3, 379–386. [Google Scholar]
- Mahfoud, H.; Wanke, S.; Neinhuis, N. A survey of leaf epicuticular waxes and trichomes in the genus Aristolochia (Aristolochiaceae) using scanning electron microscopy (SEM). SSRG Int. J. Agric. Environ. Sci. 2018, 5, 120–131. [Google Scholar]
- Peng, Y.D.; Gadagkar, S.R.; Li, J.; Xie, Y.Y.; Huang, X.Y.; Lu, H.Z.; Huang, B.Y.; Yu, L.Y. Aristolochia kechangensis sp. nov. (Aristolochiaceae) from Guangxi, China. Nord. J. Bot. 2019, 37, e02456. [Google Scholar] [CrossRef]
- Stace, C.A. Cuticular studies as an aid to plant taxonomy. Bull. Br. Mus. (Natl. Hist. ) Bot. 1965, 4, 3–78. [Google Scholar]
- Cronquist, A. An Integrated System of Classification of Flowering Plants, 2nd ed.; Columbia Univ. Press: New York, NY, USA, 1981. [Google Scholar]
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F. Plants Systematics: A Phylogenetic Approach, 2nd ed.; Sinauer Ass.: Sunderland, UK, 2002. [Google Scholar]
- Ni, X.L.; Peng, L.; Liu, W.Z. Structures, components and functions of secretory tissues in Houttuynia cordata. J. Integr. 2007, 49, 1734–1745. [Google Scholar] [CrossRef]
- Bickford, C.P. Ecophysiology of leaf trichomes. Funct. Plant Biol. 2016, 43, 807–814. [Google Scholar] [CrossRef]
- Görts-Van Rijn, A.R.A. Peperomia gracieana (Piperaceae), a new species from French Guiana. Brittonia 1998, 50, 56–58. [Google Scholar] [CrossRef]
- Huang, S.M.; Kelly, L.M.; Gilbert, M. Aristolochiaceae. In Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MA, USA, 2003; Volume 5, pp. 246–269. [Google Scholar]
- Guimarães, E.F.; Monteiro, D. Piper giordanoi (Piperaceae): A new species from southeastern Brazil. Novon 2008, 18, 175–177. [Google Scholar] [CrossRef]
- Van Do, T.; Wanke, S.; Neinhuis, C. Aristolochia bidoupensis sp. nov. from southern Vietnam. Nord. J. Bot. 2016, 34, 513–516. [Google Scholar] [CrossRef]
- Yunker, T.G.; Gray, W.D. Anatomy of Hawaiian Peperomias; Bernice, P., Ed.; Bishop Museum: Honolulu, HI, USA, 1934. [Google Scholar]
- Gogosz, A.M.; Boeger, M.R.T.; Negrelle, R.R.B.; Bergo, C. Anatomia foliar comparativa de nove espécies do gênero Piper (Piperaceae). Rodriguésia 2012, 63, 405–417. [Google Scholar] [CrossRef]
- Raman, V.; Galal, A.M.; Khan, I.A. An investigation of the vegetative anatomy of Piper sarmentosum, and a comparison with the anatomy of Piper betle (Piperaceae). Am. J. Plant Sci. 2012, 3, 1135–1144. [Google Scholar] [CrossRef]
- dos Santos, V.L.P.; Raman, V.; Bobek, V.B.; Migacz, I.P.; Franco, C.R.C.; Khan, I.A.; Budel, J.M. Anatomy and microscopy of Piper caldense, a folk medicinal plant from Brazil. Rev. Bras. Farmacogn. 2018, 28, 9–15. [Google Scholar] [CrossRef]
- Parkhurst, D.F. The adaptive significance of stomatal occurrence on one or both surfaces of leaves. J. Ecol. 1978, 66, 367–383. [Google Scholar] [CrossRef]
- Tseng, Y.C.; Xia, N.H.; Gilbert, M.G. Piperaceae. In Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MA, USA, 1999; Volume 4, pp. 110–129. [Google Scholar]
- Royer, D.L. Stomatal density and stomatal index as indicators of paleoatmospheric CO2 concentration. Rev. Palaeobot. Palynol. 2001, 114, 1–28. [Google Scholar] [CrossRef]
- Rossatto, D.R.; Kolb, R.M. Gochnatia polymorpha (Less.) Cabrera (Asteraceae) changes in leaf structure due to differences in light and edaphic conditions. Acta Bot. Bras. 2010, 24, 605–612. [Google Scholar] [CrossRef]
- Galmes, J.; Flexas, J.; Save, H.; Medrano, H. Water relation and stomatal characteristics of Mediterranean plants with different growth forms and leaf habits: Responses to water stress and recovery. Plant Soil 2007, 290, 139–155. [Google Scholar] [CrossRef]
- Driscoll, S.P.; Prins, A.; Olmos, E.; Kunert, K.J.; Foyer, C.H. Specification of adaxial and abaxial stomata, epidermal structure and photosynthesis to CO2 enrichment in maize leaves. J. Exp. Bot. 2006, 57, 381–390. [Google Scholar] [CrossRef]
- Kondorosi, E.; Roudier, F.; Gendreau, E. Plant cell-size control: Growing by ploidy? Curr. Opin. Plant Biol. 2000, 3, 488–492. [Google Scholar] [CrossRef]
- Mizukami, Y. A matter of size: Developmental control of organ size in plants. Curr. Opin. Plant Biol. 2001, 4, 533–539. [Google Scholar] [CrossRef]
- Yasmin, G.; Khan, M.A.; Shaheen, N.; Hayat, M.Q. Taxonomic significance of leaf epidermal anatomy of selected Persicaria Mill. species of family Polygonaceae from Pakistan. Afr. J. Biotechnol. 2010, 9, 3759–3768. [Google Scholar]
- Yamaki, K.; Terabayashi, S.; Okada, M.; Pak, J.H. A new species and a new variety of Asiasarum (Aristolochiaceae) from Korea. J. Jpn. Bot. 1996, 71, 1–10. [Google Scholar]
- Yook, C.S.; Kim, J.G. A new species and two new forma of Asiasarum. Kor. J. Pharmacogn. 1996, 27, 342–346. [Google Scholar]
- Oh, B.U.; Nam, O.H.; Kim, J.G. A new species of Asarum sect. Asiasarum from Korea: A. misandrum B. Oh et J. Kim. Korean J. Plant Taxon. 1997, 4, 491–499. [Google Scholar]
- Lee, Y.N.; Jeon, Y.S.; Kim, Y.S. Two new species, two new varieties and two new forms of Asarum. Bull. Korea Plant Res. 2005, 5, 5–10. [Google Scholar]
- Choi, G. List of Fake Korean Medicinal Herbs; Korea Institute of Oriental Medicine: Daejeon, Korea, 2016; p. 75. ISBN 978-89-5970-249-7. [Google Scholar]
- Moon, B.C.; Choi, G.; Yuan, Y. Origins of Herbal Medicines and Adulterants in Korea and China; Korea Institute of Oriental Medicine: Daejeon, Korea, 2017; ISBN 978-89-5970-310-4. [Google Scholar]
- Zhao, Z.; Xiao, P.; Xiao, Y.; Yuen, J.P. Quality assurance of Chinese herbal medicines (CHMs). J. Food Drug Anal. 2007, 15, 337–346. [Google Scholar]
- Song, J.H.; Moon, B.C.; Choi, G.; Yang, S. Morphological identification of Lepidii Seu Descurainiae Semen and adulterant seeds using microscopic analysis. Appl. Sci. 2018, 8, 2134. [Google Scholar] [CrossRef]
- Park, I.; Song, J.H.; Yang, S.; Kim, W.J.; Choi, G.; Moon, B.C. Cuscuta species identification based on the morphology of reproductive organs and complete chloroplast genome sequences. Int. J. Mol. Sci. 2019, 20, 2726. [Google Scholar] [CrossRef]
- Bantho, S.; Naidoo, Y.; Dewir, Y.H. The secretory scales of Combretum erythrophyllum (Combretaceae): Micromorphology, ultrastructure and histochemistry. S. Afr. J. Bot. 2020, 131, 104–117. [Google Scholar] [CrossRef]
- Salisbury, E. On the causes and ecological significance of stomatal frequency, with special reference to the woodland flora. Philos. Trans. R. Soc. Lond. Ser. B Contain. Pap. Biol. Character 1928, 216, 1–65. [Google Scholar]
- Holland, N.; Richardson, A.D. Stomatal length correlates with elevation of growth in four temperate species. J. Sustain. For. 2009, 28, 63–73. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; ISBN 978-0387981406. [Google Scholar]
- Esau, K. Anatomy of Seed Plants, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1977; 767p. [Google Scholar]
Sample Availability: All samples of the studied taxa are available from the authors, the herbarium of KIOM, and the Ministry of Food and Drug Safety. |






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.-H.; Yang, S.; Choi, G. Taxonomic Implications of Leaf Micromorphology Using Microscopic Analysis: A Tool for Identification and Authentication of Korean Piperales. Plants 2020, 9, 566. https://doi.org/10.3390/plants9050566
Song J-H, Yang S, Choi G. Taxonomic Implications of Leaf Micromorphology Using Microscopic Analysis: A Tool for Identification and Authentication of Korean Piperales. Plants. 2020; 9(5):566. https://doi.org/10.3390/plants9050566
Chicago/Turabian StyleSong, Jun-Ho, Sungyu Yang, and Goya Choi. 2020. "Taxonomic Implications of Leaf Micromorphology Using Microscopic Analysis: A Tool for Identification and Authentication of Korean Piperales" Plants 9, no. 5: 566. https://doi.org/10.3390/plants9050566
APA StyleSong, J.-H., Yang, S., & Choi, G. (2020). Taxonomic Implications of Leaf Micromorphology Using Microscopic Analysis: A Tool for Identification and Authentication of Korean Piperales. Plants, 9(5), 566. https://doi.org/10.3390/plants9050566