Expression of Genes Involved in Heavy Metal Trafficking in Plants Exposed to Salinity Stress and Elevated Cd Concentrations

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Cadmium Treatment up to 10 mM Does Not Affect the Growth and Development of Soil-Grown Common Ice Plants
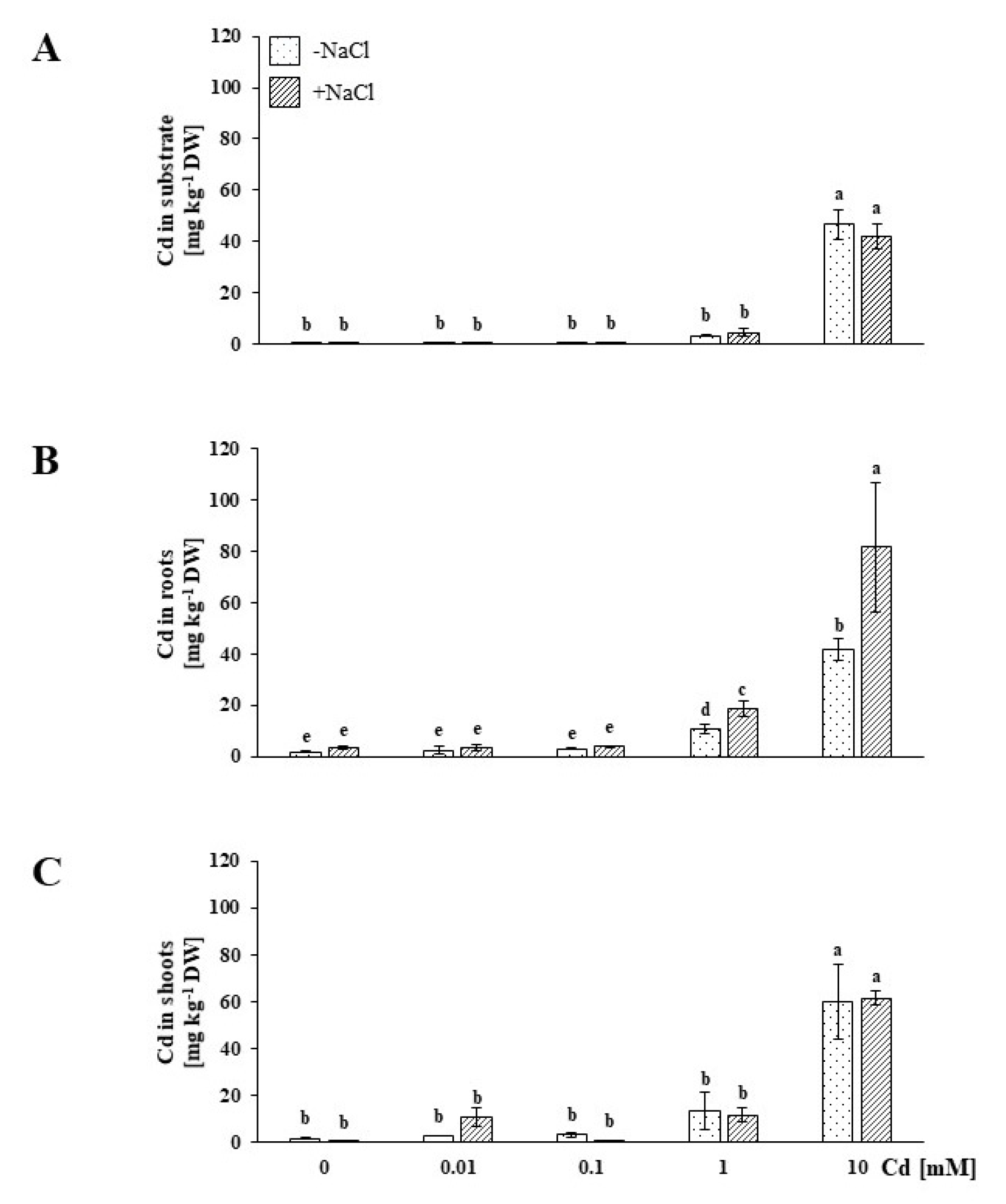
2.2. NaCl-Stressed Plants Accumulated More Cd than NaCl-Untreated Plants; Cd Was Deposited mostly in the Roots
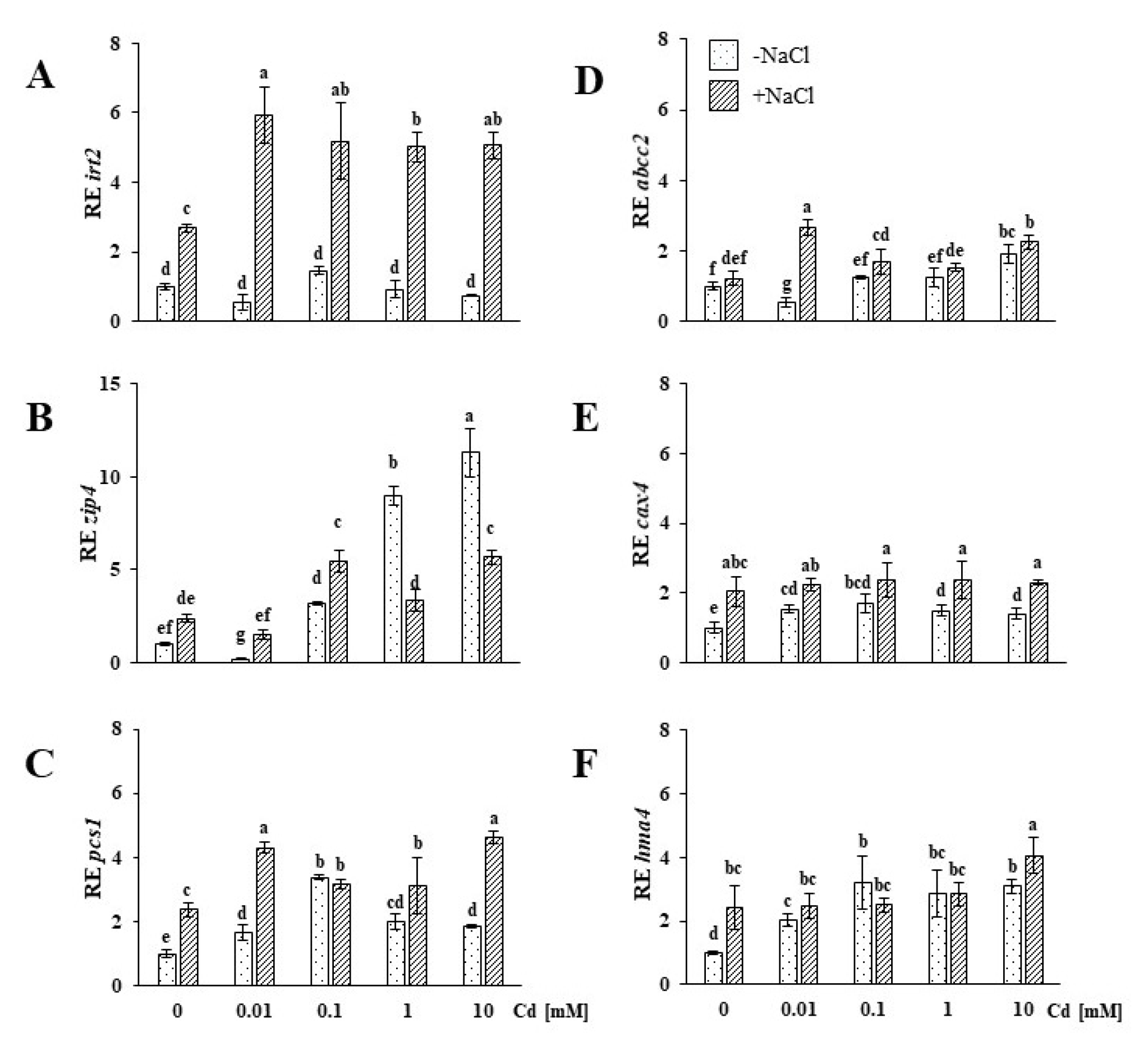
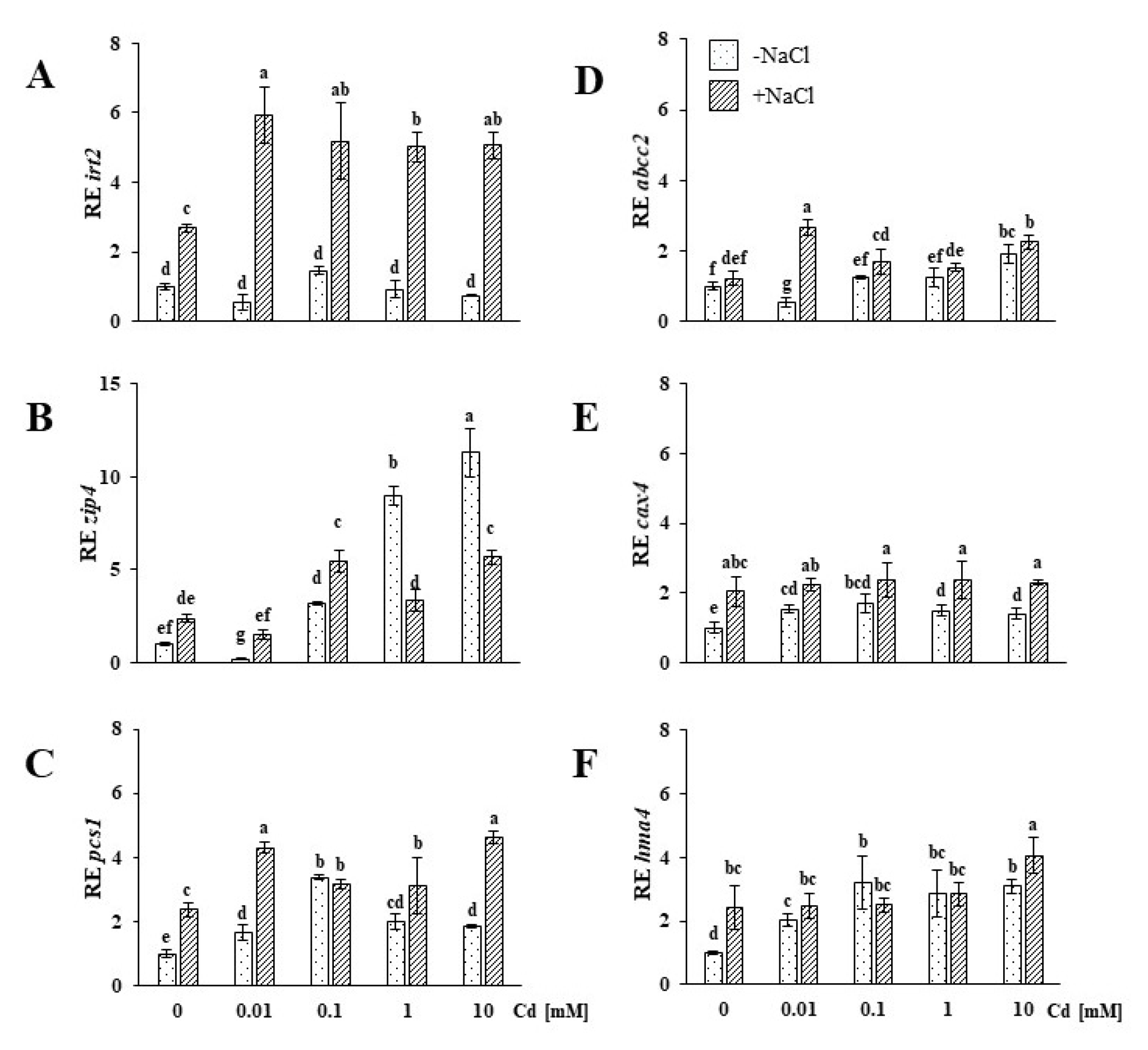
2.3. Genes Involved in Divalent Cation Uptake, Vacuolar Sequestration and Translocation Are Upregulated in the Roots of Plants Exposed to Salinity Stress
3. Discussion
4. Materials and Methods
4.1. Plant Cultivation and Cd Treatment
4.2. Biometric Analysis
4.3. Cadmium Concentration Analysis
4.4. RNA Preparation
4.5. qPCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Machado, R.; Serralheiro, R. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanetti, F.; Zegada-Lizarazu, W.; Lambertini, C.; Monti, A. Salinity effects on germination, seedlings and full-grown plants of upland and lowland switchgrass cultivars. Biomass Bioenerg. 2019, 120, 273–280. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hakeem, K.; Nahar, K.; Alharby, H. Plant Abiotic Stress Tolerance Agronomic, Molecular and Biotechnological Approaches, Plant Abiotic Stress Tolerance; Springer Nature: Cham, Switzerland, 2019. [Google Scholar]
- Asrari, E. Heavy Metal Contamination of Water and Soil: Analysis, Assessment, and Remediation Strategies; Apple Academic Press: New York, NY, USA, 2014. [Google Scholar]
- Lutts, S.; Lefèvre, I. How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann. Bot. 2015, 115, 509–528. [Google Scholar] [CrossRef]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Amari, T.; Ghnaya, T.; Debez, A.; Taamali, M.; Youssef, N.B.; Lucchini, G.; Sacchi, G.A.; Abdelly, C. Comparative Ni tolerance and accumulation potentials between Mesembryanthemum crystallinum (halophyte) and Brassica juncea: Metal accumulation, nutrient status and photosynthetic activity. J. Plant Physiol. 2014, 171, 1634–1644. [Google Scholar] [CrossRef]
- Wang, H.L.; Tian, C.Y.; Jiang, L.; Wang, L. Remediation of heavy metals contaminated saline soils: A halophyte choice? Environ. Sci. Technol. 2014, 48, 21–22. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Aslam, R.; Bostan, N.; e-Amen, N.; Maria, M.; Safdar, W. A critical review on halophytes: Salt tolerant plants. J. Med. Plant Res. 2011, 5, 7108–7118. [Google Scholar]
- Sruthi, P.; Shackira, A.M.; Puthur, J.T. Heavy metal detoxification mechanisms in halophytes: An overview. Wetl. Ecol. Manag. 2017, 25, 129–148. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.K.; Cheng, Y.; Kanwar, M.K.; Chu, X.Y.; Ahammed, G.J.; Qi, Z.Y. Responses of plant proteins to heavy metal stress—A review. Front. Plant Sci. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siemianowski, O.; Barabasz, A.; Kendziorek, M.; Ruszczyńska, A.; Bulska, E.; Williams, L.E.; Antosiewicz, D.M. HMA4 expression in tobacco reduces Cd accumulation due to the induction of the apoplastic barrier. J. Exp. Bot. 2014, 65, 1125–1139. [Google Scholar] [CrossRef] [Green Version]
- Clemente, R.; Walker, D.J.; Pardo, T.; Martínez-Fernández, D.; Bernal, M.P. The use of a halophytic plant species and organic amendments for the remediation of a trace elements-contaminated soil under semi-arid conditions. J. Hazard. Mater. 2012, 223, 63–71. [Google Scholar] [CrossRef]
- Manousaki, E.; Kalogerakis, N. Phytoextraction of Pb and Cd by the Mediterranean saltbush (Atriplex halimus L.): Metal uptake in relation to salinity. Environ. Sci. Pollut. Res. 2019, 16, 844–854. [Google Scholar] [CrossRef]
- Sharma, A.; Gontia, I.; Agarwal, P.K.; Jha, B. Accumulation of heavy metals and its biochemical responses in Salicornia brachiata, an extreme halophyte. Mar. Biol. Res. 2010, 6, 511–518. [Google Scholar] [CrossRef]
- Amari, T.; Ghnaya, T.; Sghaier, S.; Porrini, M.; Lucchini, G.; Attilio, G.; Abdelly, C. Evaluation of the Ni2+ phytoextraction potential in Mesembryanthemum crystallinum (halophyte) and Brassica juncea. J. Bioremediat. Biodegrad. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Ghnaya, T.; Nouairi, I.; Slama, I.; Messedi, D.; Grignon, C.; Abdelly, C.; Ghorbel, M.H. Cadmium effects on growth and mineral nutrition of two halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum. J. Plant Physiol. 2005, 162, 1133–1140. [Google Scholar] [CrossRef]
- Nosek, M.; Kaczmarczyk, A.; Śliwa, M.; Jędrzejczyk, R.; Kornaś, A.; Supel, P.; Kaszycki, P.; Miszalski, Z. The response of a model C3/CAM intermediate semi-halophyte Mesembryanthemum crystallinum L. to elevated cadmium concentrations. J. Plant Physiol. 2019, 240, 153005. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.-F.; Curie, C. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002, 133, 1223–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vert, G.; Briat, J.F.; Curie, C. Arabidopsis IRT2 gene encodes a root-periphery iron transporter. Plant J. 2001, 26, 181–189. [Google Scholar] [CrossRef] [PubMed]
- He, X.L.; Fan, S.K.; Zhu, J.; Guan, M.Y.; Liu, X.X.; Zhang, Y.S.; Jin, C.W. Iron supply prevents Cd uptake in Arabidopsis by inhibiting IRT1 expression and favoring competition between Fe and Cd uptake. Plant Soil 2017, 416, 453–462. [Google Scholar] [CrossRef]
- Qureshi, M.I.; D’Amici, G.M.; Fagioni, M.; Rinalducci, S.; Zolla, L. Iron stabilizes thylakoid protein-pigment complexes in Indian mustard during Cd-phytoremediation as revealed by BN-SDS-PAGE and ESI-MS/MS. J. Plant Physiol. 2010, 167, 761–770. [Google Scholar] [CrossRef]
- Muneer, S.; Jeong, B.R.; Kim, T.H.; Lee, J.H.; Soundararajan, P. Transcriptional and physiological changes in relation to Fe uptake under conditions of Fe-deficiency and Cd-toxicity in roots of Vigna radiata L. J. Plant Res. 2014, 127, 731–742. [Google Scholar] [CrossRef]
- Siedlecka, A.; Krupa, Z. Cd/Fe interaction in higher plants—Its consequences for the photosynthetic apparatus. Photosynthetica 1999, 36, 321–331. [Google Scholar] [CrossRef]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Yang, Q.; Ma, X.; Luo, S.; Gao, J.; Yang, X.; Feng, Y. SaZIP4, an uptake transporter of Zn/Cd hyperaccumulator Sedum alfredii Hance. Environ. Exp. Bot. 2018, 155, 107–117. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2000, 53, 1–11. [Google Scholar] [CrossRef]
- Rea, P.A. Phytochelatin synthase: Of a protease a peptide polymerase made. Physiol. Plant. 2012, 145, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.-B.; Smith, A.P.; Howden, R.; Dietrich, W.M.; Bugg, S.; O’Connell, M.J.; Goldsbrough, P.B.; Cobbett, C.S. Phytochelatin synthase genes from Arabidopsis and the yeast Schizosaccharomyces pombe. Plant Cell 1999, 11, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Howden, R.; Goldsbrough, P.B.; Andersen, C.R.; Cobbett, C.S. Cadmium-sensitive, cad1 mutants of Arabidopsis thaliana are phytochelatin deficient. Plant Physiol. 1995, 107, 1059–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zitka, O.; Krystofova, O.; Sobrova, P.; Adam, V.; Zehnalek, J.; Beklova, M.; Kizek, R. Phytochelatin synthase activity as a marker of metal pollution. J. Hazard. Mater. 2011, 192, 794–800. [Google Scholar] [CrossRef]
- Szarka, A.; Tomasskovics, B.; Bánhegyi, G. The ascorbate-glutathione-α-tocopherol triad in abiotic stress response. Int. J. Mol. Sci. 2012, 13, 4458–4483. [Google Scholar] [CrossRef] [Green Version]
- Kuźniak, E.; Kaźmierczak, A.; Wielanek, M.; Głowacki, R.; Kornas, A. Involvement of salicylic acid, glutathione and protein S-thiolation in plant cell death-mediated defence response of Mesembryanthemum crystallinum against Botrytis cinerea. Plant Physiol. Biochem. 2013, 63, 30–38. [Google Scholar] [CrossRef]
- Pomponi, M.; Censi, V.; Di Girolamo, V.; De Paolis, A.; Di Toppi, L.S.; Aromolo, R.; Costantino, P.; Cardarelli, M. Overexpression of Arabidopsis phytochelatin synthase in tobacco plants enhances Cd2+ tolerance and accumulation but not translocation to the shoot. Planta 2006, 223, 180–190. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.G.; Butko, E.; Springer, F.; Torpey, J.W.; Komives, E.A.; Kehr, J.; Schroeder, J.I. Identification of high levels of phytochelatins, glutathione and cadmium in the phloem sap of Brassica napus. A role for thiol-peptides in the long-distance transport of cadmium and the effect of cadmium on iron translocation. Plant J. 2008, 54, 249–259. [Google Scholar]
- Mendoza-Cózatl, D.G.; Jobe, T.O.; Hauser, F.; Schroeder, J.I. Long-distance transport, vacuolar sequestration, tolerance, and transcriptional responses induced by cadmium and arsenic. Curr. Opin. Plant Biol. 2011, 14, 554–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, P.; Zanella, L.; De Paolis, A.; Di Litta, D.; Cecchetti, V.; Falasca, G.; Barbieri, M.; Altamura, M.M.; Costantino, P.; Cardarelli, M. Cadmium-inducible expression of the ABC-type transporter AtABCC3 increases phytochelatin-mediated cadmium tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 3815–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhati, K.K.; Sharma, S.; Aggarwal, S.; Kaur, M.; Shukla, V.; Kaur, J.; Mantri, S.; Pandey, A.K. Genome-wide identification and expression characterization of ABCC-MRP transporters in hexaploid wheat. Front. Plant Sci. 2015, 6, 488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.D.; Zhao, K.X.; Yang, Z.M. Identification of genomic ATP binding cassette (ABC) transporter genes and Cd-responsive ABCs in Brassica napus. Gene 2018, 664, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Manohar, M.; Shigaki, T.; Hirschi, K.D. Plant cation/H+ exchangers (CAXs): Biological functions and genetic manipulations. Plant Biol. 2011, 13, 561–569. [Google Scholar] [CrossRef]
- Koren’kov, V.; Park, S.; Cheng, N.H.; Sreevidya, C.; Lachmansingh, J.; Morris, J.; Hirschi, K.; Wagner, G.J. Enhanced Cd2+-selective root-tonoplast-transport in tobaccos expressing Arabidopsis cation exchangers. Planta 2007, 225, 403–411. [Google Scholar] [CrossRef]
- Mei, H.; Hirschi, K.D.; Cheng, N.H.; Zhao, J.; Park, S.; Escareno, R.A.; Pittman, J.K. Root development under metal stress in Arabidopsis thaliana requires the H+/cation antiporter CAX4. New Phytol. 2009, 183, 95–105. [Google Scholar] [CrossRef]
- Bassil, E.; Blumwald, E. The ins and outs of intracellular ion homeostasis: NHX-type cation/H+ transporters. Curr. Opin. Plant Biol. 2014, 22, 1–6. [Google Scholar] [CrossRef]
- Jiang, X.; Leidi, E.O.; Pardo, J.M. How do vacuolar NHX exchangers function in plant salt tolerance? Plant Signal. Behav. 2010, 5, 792–795. [Google Scholar] [CrossRef] [Green Version]
- Barkla, B.J.; Zingarelli, L.; Blumwald, E.; Smith, J. Tonoplast Na+/H+ antiport activity and its energization by the vacuolar H+-ATPase in the halophytic plant Mesembryanthemum crystallinum L. Plant Physiol. 1995, 109, 549–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratajczak, R.; Richter, J.; Luttge, U. Adaptation of the tonoplast V-type H+-ATPase of Mesembryanthemum crystallinum to salt stress, C3–CAM transition and plant age. Plant. Cell Environ. 1994, 17, 1101–1112. [Google Scholar] [CrossRef]
- Cheng, N.; Pittman, J.K.; Shigaki, T.; Hirschi, K.D. Characterization of CAX4, an Arabidopsis H+/cation antiporter. Plant Physiol. 2002, 128, 1245–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, R.F.; Francini, A.; Ferreira Da Rocha, P.S.C.; Baccarini, P.J.; Aylett, M.; Krijger, G.C.; Williams, L.E. The plant P1B-type ATPase AtHMA4 transports Zn and Cd and plays a role in detoxification of transition metals supplied at elevated levels. FEBS Lett. 2005, 579, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.; Cobbett, E. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yin, H.; Liu, X.; Li, X. Salt affects plant Cd-stress responses by modulating growth and Cd accumulation. Planta 2010, 231, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.F.; Krijgert, G.C.; Baccarini, P.J.; Hall, J.L.; Williams, L.E. Functional expression of AtHMA4, a P1B-type ATPase of the Zn/Co/Cd/Pb subclass. Plant J. 2003, 35, 164–176. [Google Scholar] [CrossRef]
- Bernard, C.; Roosens, N.; Czernic, P.; Lebrun, M.; Verbruggen, N. A novel CPx-ATPase from the cadmium hyperaccumulator Thlaspi caerulescens. FEBS Lett. 2004, 569, 140–148. [Google Scholar] [CrossRef]
- Papoyan, A. Identification of Thlaspi caerulescens genes that may be involved in heavy metal hyperaccumulation and tolerance. Characterization of a novel heavy metal transporting ATPase. Plant Physiol. 2004, 136, 3814–3823. [Google Scholar] [CrossRef] [Green Version]
- Houba, V.J.G.; Temminghoff, E.J.M.; Gaikhorst, G.A.; van Vark, W. Soil analysis procedures using 0.01 M calcium chloride as extraction reagent. Commun. Soil Sci. Plant Anal. 2000, 31, 1299–1396. [Google Scholar] [CrossRef]
- Rozpądek, P.; Domka, A.; Ważny, R.; Nosek, M.; Jędrzejczyk, R.; Tokarz, K.; Turnau, K. How does the endophytic fungus Mucor sp. improve Arabidopsis arenosa vegetation in the degraded environment of a mine dump? Environ. Exp. Bot. 2018, 147, 31–42. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acid Res. 2001, 29, 16–21. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Cadmium Concentration [mM] | Root DW [g/Plant] | Shoot DW [g/Plant] | ||
|---|---|---|---|---|
| −NaCl | +NaCl | −NaCl | +NaCl | |
| 0 | 0.29 ± 0.06a | 0.18 ± 0.06abc | 1.93 ± 0.13a | 2.03 ± 0.58a |
| 0.01 | 0.27 ± 0.05a | 0.13 ± 0.01bc | 1.90 ± 0.18a | 2.20 ± 0.41a |
| 0.1 | 0.20 ± 0.05abc | 0.21 ± 0.09abc | 1.97 ± 0.22a | 2.29 ± 0.57a |
| 1.0 | 0.26 ± 0.07a | 0.25 ± 0.04ab | 2.09 ± 0.33a | 2.06 ± 0.33a |
| 10.0 | 0.21 ± 0.03abc | 0.12 ± 0.04c | 1.80 ± 0.35a | 1.97 ± 0.37a |
| Cadmium Concentration [mM] | Root Water Content [cm3 g−1 DW] | Shoot Water Content [cm3 g−1 DW] | ||
|---|---|---|---|---|
| −NaCl | +NaCl | −NaCl | +NaCl | |
| 0 | 11.78 ± 4.27ab | 8.77 ± 2.41abc | 22.27 ± 1.43a | 14.54 ± 1.48b |
| 0.01 | 8.97 ± 0.53abc | 6.47 ± 1.03bc | 23.10 ± 2.22a | 15.10 ± 1.37b |
| 0.1 | 6.79 ± 0.89bc | 5.34 ± 0.28bc | 22.49 ± 2.84a | 15.37 ± 1.48b |
| 1.0 | 6.94 ± 0.32bc | 7.99 ± 0.94abc | 22.70 ± 2.75a | 15.46 ± 1.53b |
| 10.0 | 7.23 ± 1.15abc | 10.04 ± 2.55ab | 23.24 ± 2.61a | 14.98 ± 2.81b |
| Cadmium Concentration [mM] | Shoot-to-Root (DW) Ratio | |
|---|---|---|
| −NaCl | +NaCl | |
| 0 | 7.01 ± 1.54c | 10.22 ± 1.74bc |
| 0.01 | 7.11 ± 0.84c | 15.51 ± 3.75ab |
| 0.1 | 9.57 ± 1.75bc | 11.13 ± 3.19b |
| 1.0 | 7.58 ± 2.33c | 8.57 ± 0.78c |
| 10.0 | 7.44 ± 0.54c | 16.88 ± 3.46ab |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nosek, M.; Kaczmarczyk, A.; Jędrzejczyk, R.J.; Supel, P.; Kaszycki, P.; Miszalski, Z. Expression of Genes Involved in Heavy Metal Trafficking in Plants Exposed to Salinity Stress and Elevated Cd Concentrations. Plants 2020, 9, 475. https://doi.org/10.3390/plants9040475
Nosek M, Kaczmarczyk A, Jędrzejczyk RJ, Supel P, Kaszycki P, Miszalski Z. Expression of Genes Involved in Heavy Metal Trafficking in Plants Exposed to Salinity Stress and Elevated Cd Concentrations. Plants. 2020; 9(4):475. https://doi.org/10.3390/plants9040475
Chicago/Turabian StyleNosek, Michał, Adriana Kaczmarczyk, Roman J. Jędrzejczyk, Paulina Supel, Paweł Kaszycki, and Zbigniew Miszalski. 2020. "Expression of Genes Involved in Heavy Metal Trafficking in Plants Exposed to Salinity Stress and Elevated Cd Concentrations" Plants 9, no. 4: 475. https://doi.org/10.3390/plants9040475