Foliar Application of Calcium and Growth Regulators Modulate Sweet Cherry (Prunus avium L.) Tree Performance




Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Site and Plant Material
2.2. Experimental Design and Treatments
2.3. Leaf Gas Exchange
2.4. Leaf Water Status
2.5. Metabolite Composition Determination
2.5.1. Photosynthetic Pigments
2.5.2. Total Soluble Sugars and Starch
2.5.3. Soluble Proteins
2.6. Yield Determination
2.7. Fruit Cracking Index Determination
2.8. Statistical Analysis
3. Results
3.1. Leaf Gas Exchange Parameters
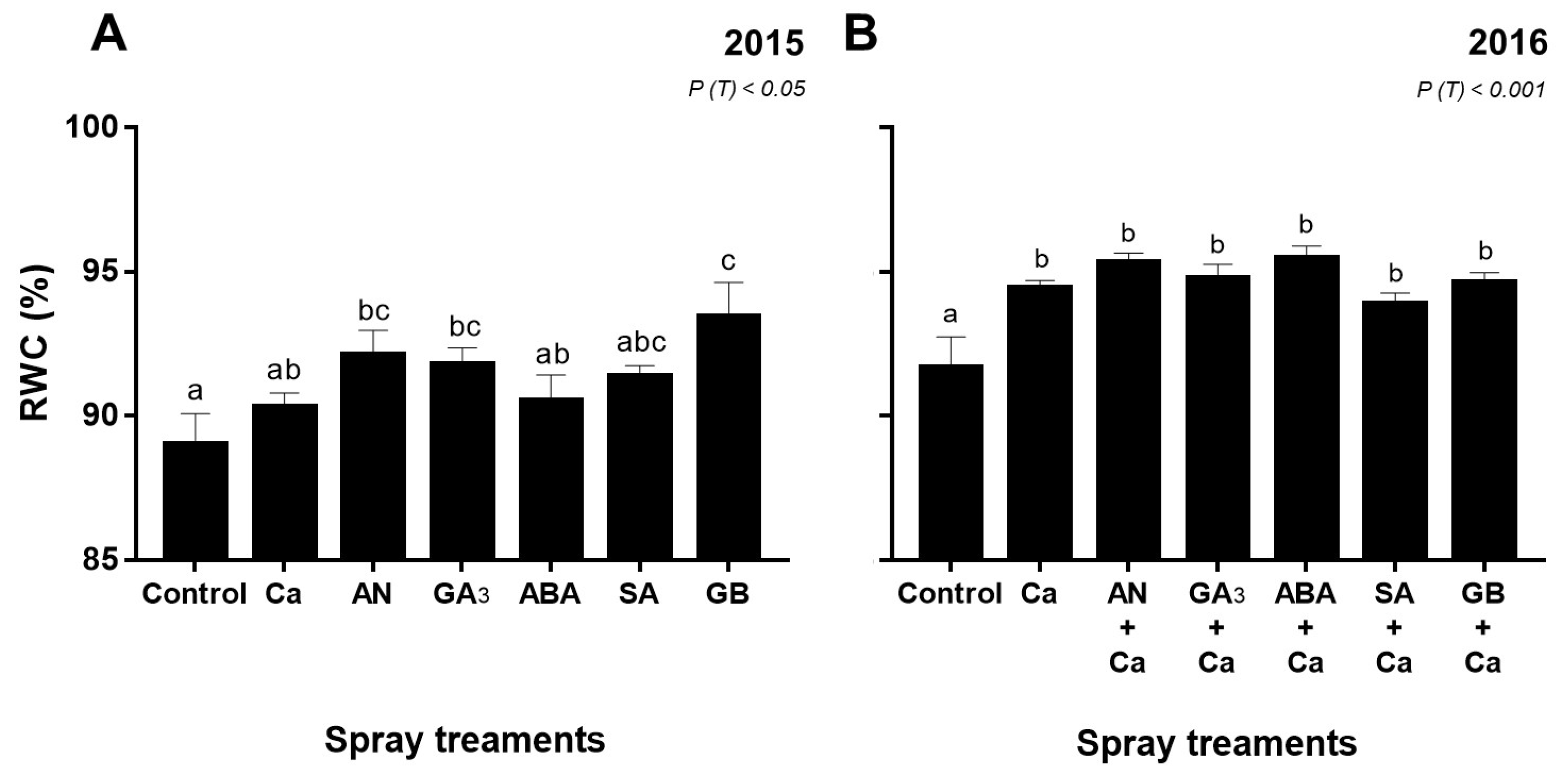
3.2. Leaf Water Status
3.3. Leaf Photosynthetic Pigments and Metabolites
3.4. Yield of Sweet Cherry Trees
3.5. Cracking Incidence
4. Discussion
4.1. Spray Treatments Modulate Leaf Gas Exchange and Water Status of Sweet Cherry Trees
4.2. Photosynthetic Pigments’ and Metabolites’ Behavior in Response to Spray Treatments
4.3. Effect of Spray Treatments on Yield of Sweet Cherry Trees and Fruit Cracking Incidence
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Global Warming of 1.5 °C. An. IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas. Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- Hanson, J.B. The function of calcium in plant nutrition. In Advances in Plant Nutrition; Tinker, P.B., Lauchli, A., Eds.; Praeger: New York, NY, USA, 1984; pp. 149–208. [Google Scholar]
- Demirsoy, L.; Bilgener, S. The effects of preharvest calcium hydroxide applications on cracking in ‘0900 Ziraat’, ‘Lambert’ and ‘Van’ sweet cherries. Acta Hortic. 1998, 468, 657–662. [Google Scholar] [CrossRef]
- Vangdal, E.; Hovland, K.L.; Børve, J.; Sekse, L.; Slimestad, R. Foliar application of calcium reduces postharvest decay in sweet cherry by various mechanisms. Acta Hortic. 2008, 768, 143–148. [Google Scholar] [CrossRef]
- Wójcik, P.; Akgül, H.; Demirtaş, I.; Sarısu, C.; Aksu, M.; Gubbuk, H. Effect of preharvest sprays of calcium chloride and sucrose on cracking and quality of Burlat sweet cherry fruit. J. Plant. Nutr. 2013, 36, 1453–1465. [Google Scholar] [CrossRef]
- Erogul, D. Effect of preharvest calcium treatments on sweet cherry fruit quality. Not. Bot. Hort. Agrobot. 2014, 42, 150–153. [Google Scholar] [CrossRef]
- Xu, C.; Li, X.; Zhang, L. The effect of calcium chloride on growth, photosynthesis, and antioxidant responses of Zoysia japonica under drought conditions. PLoS ONE 2013, 8, e68214. [Google Scholar] [CrossRef]
- Naeem, M.; Naeem, M.S.; Ahmad, R.; Ihsan, M.Z.; Ashraf, M.Y.; Hussain, Y.; Fahad, S. Foliar calcium spray confers drought stress tolerance in maize via modulation of plant growth, water relations, proline content and hydrogen peroxide activity. Arch. Agron. Soil Sci. 2017, 64, 116–131. [Google Scholar] [CrossRef]
- Razem, F.A.; Baron, K.; Hill, R.D. Turning on gibberellin and abscisic acid signaling. Curr. Opin. Plant. Biol. 2006, 9, 454–459. [Google Scholar] [CrossRef]
- Erogul, D.; Sen, F. Effects of gibberellic acid treatments on fruit thinning and fruit quality in Japanese plum (Prunus salicina Lindl.). Sci. Hortic. 2015, 186, 137–142. [Google Scholar] [CrossRef]
- Ozkan, Y.; Ucar, M.; Yildiz, K.; Ozturkc, B. Pre-harvest gibberellic acid (GA3) treatments play an important role on bioactive compounds and fruit quality of sweet cherry cultivars. Sci. Hortic. 2016, 211, 358–362. [Google Scholar] [CrossRef]
- Cline, J.A.; Trought, M. Effect of gibberellic acid on fruit cracking and quality of ‘Bing’ and ‘Sam’ sweet cherries. Can. J. Plant. Sci. 2007, 87, 545–550. [Google Scholar] [CrossRef]
- Horvitz, S.; López Camelo, A.F.; Yommi, A.; Godoy, C. Application of gibberellic acid to ‘sweetheart’ sweet cherries: Effects on fruit quality at harvest and during cold storage. Acta Hortic. 2003, 628, 311–316. [Google Scholar] [CrossRef]
- Hoppe, F.; Huyskens-Keil, S.; Ulrichs, C.; Hanrahan, I. Assessment of susceptibility and prevention of cracking of ‘Skeena’ sweet cherry. In Proceedings of the II International Symposium on Horticulture in Europe 1099, Angers, France, 1–5 July 2012; pp. 819–826. [Google Scholar]
- Usenik, V.; Kastelec, D.; Stampar, F. Physicochemical changes of sweet cherry fruits related to application of gibberellic acid. Food Chem. 2005, 90, 663–671. [Google Scholar] [CrossRef]
- Suran, P.; Vavra, R.; Zeleny, L. Effectiveness of potential products to reduce rain cracking of cherry fruit. Acta Hortic. 2016, 1137, 183–186. [Google Scholar] [CrossRef]
- Leung, J.; Giraudat, J. Abcisic acid signal transduction. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1998, 49, 199–222. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant. Sci. 2015, 6, 462. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Silva, E.; Gonçalves, A.; Matos, C.; Rodrigues, M.A.; Moutinho-Pereira, J.; Barros, A.; Correia, C. Kaolin and salicylic acid foliar application modulate yield, quality and phytochemical composition of olive pulp and oil from rainfed trees. Sci. Hortic. 2018, 237, 176–183. [Google Scholar] [CrossRef]
- El-Shazly, S.M.; Eisa, A.M.; Moảtamed, A.M.H.; Kotb, H.R.M. Effect of some agrochemicals preharvest foliar application on yield and fruit quality of ‘Swelling’ peach trees. Alex. J. Agric. Res. 2013, 58, 219–229. [Google Scholar]
- Mohamed, R.A.; Abdelbaset, A.K.; Abd-Elkader, D.Y. Salicylic acid effects on growth, yield, and fruit quality of strawberry cultivars. J. Med. Act. Plants 2017, 6, 1–11. [Google Scholar]
- Giménez, M.J.; Valverde, J.M.; Valero, D.; Guillén, F.; Martínez-Romero, D.; Serrano, M.; Castillo, S. Quality and antioxidant properties on sweet cherries as affected by preharvest salicylic and acetylsalicylic acids treatments. Food Chem. 2014, 160, 226–232. [Google Scholar] [CrossRef]
- Correia, S.; Queirós, F.; Ribeiro, C.; Vilela, A.; Aires, A.; Barros, A.I.; Schouten, R.; Silva, A.P.; Gonçalves, B. Effects of calcium and growth regulators on sweet cherry (Prunus avium L.) quality and sensory attributes at harvest. Sci. Hortic. 2019, 248, 231–240. [Google Scholar] [CrossRef]
- Jiang, F.; Hartung, W. Long-distance signalling of abscisic acid (ABA): The factors regulating the intensity of the ABA signal. J. Exp. Bot. 2008, 59, 37–43. [Google Scholar] [CrossRef]
- Balbontín, C.; Gutiérrez, C.; Wolff, M.; Figueroa, C.R. Effect of abscisic acid and methyl jasmonate preharvest applications on fruit quality and cracking tolerance of sweet cherry. Chil. J. Agric. Res. 2018, 78, 438–446. [Google Scholar] [CrossRef]
- Ma, X.L.; Wang, Y.J.; Xie, S.L.; Wang, C.; Wang, W. Glycine betaine application ameliorates negative effects of drought stress in tobacco. Russ. J. Plant. Physiol. 2007, 54, 472–479. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant. Sci. 2017, 7, 2049. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant. Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Etienne, P.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F. Brassica napus growth is promoted by Ascophyllum nodosum (L.) seaweed extract: Microarray analysis and physiological characterization of N, C, and S metabolisms. J. Plant. Growth Regul. 2013, 32, 31–52. [Google Scholar] [CrossRef]
- Correia, S.; Oliveira, I.; Queirós, F.; Ribeiro, C.; Ferreira, L.; Luzio, A.; Silva, A.P.; Gonçalbes, B. Preharvest application of seaweed based biostimulant reduced cherry (Prunus avium L.) cracking. In Proceedings of the Environmental Sciences-Agriculture and Climate Change—Adapting Crops to Increased Uncertainty (AGRI 2015), Amsterdam, The Netherlands, 15–17 February 2015; Volume 29, pp. 251–252. [Google Scholar]
- Ayala, M.; Lang, G.A. Chapter 12: Morphology, cropping physiology and canopy training. In Cherries: Botany, Production and Uses; Quero-García, J., Iezzoni, A., Puławska, J., Lang, G., Eds.; CABI: Wallingford, UK, 2017; pp. 269–304. [Google Scholar]
- Von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Düring, H. Photosynthesis of ungrafted and grafted grapevines: Effects of rootstock genotype and plant age. Am. J. Enol. Vitic. 1994, 45, 297–299. [Google Scholar]
- Šesták, Z.; Castky, J.; Jarvis., P.G. Plant. Photosynthetic Production. Manual of Methods; Dr. W. Junk Publishers: Haia, The Netherlands, 1971; p. 818. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method Enzymol. 1987, 148, 350–382. [Google Scholar]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Osaki, M.; Shinano, T.; Tadano, T. Redistribution of carbon and nitrogen compounds from the shoot to the harvesting organs during maturation in field crops. Soil Sci. Plant. Nutr. 1991, 37, 117–128. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein using the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Christensen, J.V. Cracking in cherries III: Determination of cracking susceptibility. Acta Agric. Scand. 1972, 22, 128–136. [Google Scholar] [CrossRef]
- Gonçalves, B.; Moutinho-Pereira, J.; Santos, A.; Silva, A.P.; Bacelar, E.; Correia, C.; Rosa, E. Scion-rootstock interaction affects the physiology and fruit quality of sweet cherry. Tree Physiol. 2005, 26, 93–104. [Google Scholar] [CrossRef]
- Yuan, L.; Xu, D.Q. Stimulation effect of gibberellic acid short-term treatment on leaf photosynthesis related to the increase in Rubisco content in broad bean and soybean. Photosyn. Res. 2001, 68, 39–47. [Google Scholar] [CrossRef]
- Teszlák, P.; Kocsis, M.; Gaal, K.; Nikfardjam, M.P. Regulatory effects of exogenous gibberellic acid (GA3) on water relations and CO2 assimilation among grapevine (Vitis vinifera L.) cultivars. Sci. Hortic. 2013, 159, 41–51. [Google Scholar] [CrossRef]
- Kumar, P.; Lakshmi, N.J.; Mani, V.P. Interactive effects of salicylic acid and phytohormones on photosynthesis and grain yield of soybean (Glycine max L. Merrill). Physiol. Mol. Biol. Plants 2000, 6, 179–186. [Google Scholar]
- Fariduddin, Q.; Hayat, S.; Ahmad, A. Salicylic acid influences net photosynthetic rate, carboxylation efficiency, nitrate reductase activity and seed yield in Brassica juncea. Photosynthetica 2003, 41, 281–284. [Google Scholar] [CrossRef]
- Khan, W.; Prithviraj, B.; Smith, D.L. Photosynthetic responses of corn and soybean to foliar application of salicylates. J. Plant. Physiol. 2003, 160, 485–492. [Google Scholar] [CrossRef]
- Suzy, Y.R.; Dennis, H.G.; Hutton, R.J.; Clarke, S.J. Transpiration efficiency of the grapevine cv. Semillon is tied to VPD in warm climates. Ann. Appl. Biol. 2011, 158, 106–114. [Google Scholar]
- Giri, J. Glycinebetaine and abiotic stress tolerance in plants. Plant. Signal. Behav. 2011, 11, 1746–1751. [Google Scholar] [CrossRef] [PubMed]
- Denaxa, N.K.; Roussos, P.A.; Damvakaris, T.; Stournaras, V. Comparative effects of exogenous glycine betaine, kaolin clay particles and ambiol on photosynthesis, leaf sclerophylly indexes and heat load of olive cv. Chondrolia Chalkidikis under drought. Sci. Hortic. 2012, 137, 87–94. [Google Scholar] [CrossRef]
- Mikiciuk, G.; Mikiciuk, M.; Ptak, P. The effects of anitranspirant di-1-p-menthene on some physiological traits of strawberry. Ecol. Eng. 2015, 16, 161–167. [Google Scholar] [CrossRef]
- Franks, P.J.; Farquhar, G.D. The effect of exogenous abscisic acid on stomatal development, stomatal mechanics, and leaf gas exchange in Tradescantia virginiana. Plant. Physiol. 2001, 125, 935–942. [Google Scholar] [CrossRef]
- Xu, C.; Leskovar, D.I. Effects of A. nodosum seaweed extracts on spinach growth, physiology and nutrition value under drought stress. Sci. Hortic. 2015, 183, 39–47. [Google Scholar] [CrossRef]
- El-Mageed, T.A.; Semida, W.M.; Radyc, M.M. Moringa leaf extract as biostimulant improves water use efficiency, physio-biochemical attributes of squash plants under deficit irrigation. Agr. Water Manag. 2017, 193, 46–54. [Google Scholar] [CrossRef]
- Cabo, S.; Morais, M.C.; Aires, A.; Carvalho, R.; Pascual-Seva, N.; Silva, A.P.; Gonçalves, B. Kaolin and seaweed-based extracts can be used as middle and long-term strategy to mitigate negative effects of climate change in physiological performance of hazelnut tree. J. Agro. Crop. Sci. 2019, 206, 28–42. [Google Scholar] [CrossRef]
- Ali, H.M.; Siddiqui, M.H.; Basalah, M.O.; Al-Whaibi, M.H.; Sakran, A.M.; Al-Amri, A. Effects of gibberellic acid on growth and photosynthetic pigments of Hibiscus sabdariffa L. under salt stress. Afr. J. Biotechnol. 2012, 11, 800–804. [Google Scholar]
- Hussain, S.; Saleem, M.F.; Ashraf, M.Y.; Cheema, M.A.; Haq, M.A. Abscisic acid, a stress hormone helps in improving water relations and yield of sunflower (Helianthus annuus L.) hybrids under drought. Pak. J. Bot. 2010, 42, 2177–2189. [Google Scholar]
- Brito, C.; Dinis, L.T.; Luzio, A.; Silva, E.; Gonçalves, A.; Meijón, M.; Escandón, M.; Arrobas, M.; Rodrigues, M.A.; Moutinho-Pereira, J.; et al. Kaolin and salicylic acid alleviate summer stress in rainfed olive orchards by modulation of distinct physiological and biochemical responses. Sci. Hortic. 2019, 246, 201–211. [Google Scholar] [CrossRef]
- Ragab, M.E.; Helal, N.A.S.; Sawan, O.M.; Fawzy, Z.F.; El-Sawy, S.M. Foliar application of glycine betaine for alleviating water stress of tomato plants grown under sandy soil conditions. Int. J. Chem. Tech. Res. 2015, 8, 52–67. [Google Scholar]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Abscisic acid increases carotenoid and chlorophyll concentrations in leaves and fruit of two tomato genotypes. J. Amer. Soc. Hort. Sci. 2014, 139, 261–266. [Google Scholar] [CrossRef]
- Kabiri, R.; Naisibi, F.; Farahbakhsh, H. Effect of exogenous salicylic acid on some physiological parameters and alleviation of drought stress in Nigella sativa plant under hydroponic culture. Plant. Protect. Sci. 2014, 50, 43–51. [Google Scholar] [CrossRef]
- Hayat, S.; Ahmad, A. Salicylic Acid a Plant. Hormone; Springer Publishers: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Tian, F.; Wang, W.; Liang, C.; Wang, X.; Wang, G.; Wang, W. Overaccumulation of glycine betaine makes the function of the thylakoid membrane better in wheat under salt stress. Crop. J. 2017, 5, 73–82. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A. Why and what for the leaves are yellow inautumm? On the interpretation of optical spectra of senescing leaves (Acerplatanoides L.). J. Plant. Physiol. 1995, 145, 315–320. [Google Scholar] [CrossRef]
- Luo, H.; Dai, S.; Ren, J.; Zhang, C.; Ding, Y.; Li, Z.; Sun, Y.; Ji, K.; Wang, Y.; Li, Q.; et al. The role of ABA in the maturation and postharvest life of a nonclimacteric sweet cherry fruit. J. Plant. Growth Regul. 2014, 33, 373–383. [Google Scholar] [CrossRef]
- Murcia, G.; Pontin, M.; Reinoso, H.; Baradi, R.; Bertazza, G.; Gómez-Talquenca, S.; Bottini, R.; Piccolia, P.N. ABA and GA3 increase carbon allocation in different organs of grapevine plants by inducing accumulation of non-structural carbohydrates in leaves, enhancement of phloem area and expression of sugar transporters. Physiol. Plant. 2016, 156, 323–337. [Google Scholar] [CrossRef]
- Correia, S.; Aires, A.; Queirós, F.; Carvalho, R.; Schouten, R.; Silva, A.P.; Gonçalves, B. Climate conditions and spray treatments induce shifts in health promoting compounds in cherry (Prunus avium L.) fruits. Sci. Hortic. 2020, 263, 109147. [Google Scholar] [CrossRef]
- Sasek, T.W.; De Lucía, E.H.; Strain, B.R. Reversibility of photosynthetic inhibition in cotton after long-term exposure to elevated CO2 concentrations. Plant. Physiol. 1985, 78, 612–622. [Google Scholar] [CrossRef]
- Sicher, R.C.; Bunce, J.A. Adjustments of net photosynthesis in Solanum tuberosum in response to reciprocal changes in ambient and elevated growth CO2 partial pressures. Physiol. Plantarum 2001, 112, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars-metabolism, sensing and abiotic stress a complex network in the life of plants. Plant. Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef]
- Thalmann, M.; Pazmino, D.; Seung, D.; Horrer, D.; Nigro, A.; Meier, T.; Kölling, K.; Pfeifhofer, H.W.; Zeeman, S.C.; Santelia, D. Regulation of leaf starch degradation by abscisic acid is important for osmotic stress tolerance in plants. Plant. Cell 2016, 28, 1860–1878. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, C.M.; Díaz, J.B.R. A statistical model to estimate potential yields in peach before bloom. J. Am. Soc. Hortic. Sci. 2003, 128, 297–301. [Google Scholar] [CrossRef]
- Basak, A. Effect of preharvest treatment with seaweed products, KelpakR and Goëmar BM 86R, on fruit quality in apple. Int. J. Fruit Sci. 2008, 8, 1–14. [Google Scholar] [CrossRef]
- Colavita, G.M.; Spera, N.; Blackhall, V.; Sepulveda, G.M. Effects of seaweed extract on pear fruit quality and yield. Acta Hortic. 2011, 909, 601–607. [Google Scholar] [CrossRef]
- Quiroga, A.; Berli, F.; Moreno, D.; Cavagnaro, J.; Bottini, R. Abscisic acid sprays significantly increase yield per plant in vineyard-grown wine grape (Vitis vinifera L.) cv. Cabernet Sauvignon through increased berry set with no negative effects on anthocyanin content and total polyphenol index of both juice and wine. J. Plant. Growth Regul. 2009, 28, 28–35. [Google Scholar]
- Iqbal, N.; Ashraf, M. Glycine betaine, an osmolyte of interest to improve water stress tolerance in sunflower (Helianthus annuus L.): Water relations and yield. S. Afr. J. Bot. 2008, 74, 274–281. [Google Scholar] [CrossRef]
- Anjum, S.A.; Farooq, M.; Wang, L.C.; Xue, L.L.; Wang, S.G.; Wang, L.; Zhang, S.; Chen, M. Gas exchange and chlorophyll synthesis of maize cultivars are enhanced by exogenously-applied glycine betaine under drought conditions. Plant. Soil Environ. 2011, 57, 326–331. [Google Scholar] [CrossRef]
- Khan, M.M.A.; Gautam, A.C.; Mohammad, F.; Siddiqui, M.H.; Naeem, M.; Khan, M.N. Effect of gibberellic acid spray on performance of tomato. Turk. J. Biol. 2006, 30, 11–16. [Google Scholar]
- Uddain, J.; Hossain, A.K.M.; Mostafa, M.G.; Rahman, M.J. Effect of different plant growth regulators on growth and yield of tomato. Int. J. Sustain. Agric. 2009, 1, 58–63. [Google Scholar]
- Pal, P.; Yadav, K.; Kumar, K.; Singh, N. Effect of gibberellic acid and potassium foliar sprays on productivity and physiological and biochemical parameters of parthenocarpic cucumber Cv. ‘Seven Star F1’. J. Hortic. Res. 2016, 24, 93–100. [Google Scholar] [CrossRef]
- Parry, M.A.J.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Ann. Rev. Plant. Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef]
- Getnet, Z.; Husen, A.; Fetene, M.; Yemata, G. Growth, water status, physiological, biochemical and yield response of stay green sorghum (Sorghum bicolor (L.) Moench) varieties—A field trial under drought-prone area in Amhara regional state Ethiopia. J. Agron. 2015, 14, 188–202. [Google Scholar]
- Surendar, K.K.; Devil, D.D.; Ravi, I.; Jeyakumar, P.; Velayudham, K. Studies on the impact of water deficit on morphological, physiological and yield of banana (Musa spp.) cultivars and hybrids. Int. J. Agr. Sci. 2013, 3, 473–482. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatment (T) | Chltotal (mg g−1 DW) | Carottotal (mg g−1 DW) | SS (mg g−1 DW) | St (mg g−1 DW) | SP (mg g−1 DW) |
|---|---|---|---|---|---|---|
| 2015 | Control | 7.95 ± 0.42 a | 0.93 ± 0.04 a | 70.35 ± 1.16 a | 42.98 ± 0.56 a | 10.34 ± 0.79 a |
| Ca | 8.37 ± 0.86 a | 1.15 ± 0.05 cd | 75.04 ± 3.87 abc | 41.67 ± 1.94 a | 12.30 ± 0.53 ab | |
| AN | 9.34 ± 0.27 ab | 1.23 ± 0.05 de | 85.81 ± 3.00 bc | 47.41 ± 1.89 a | 15.08 ± 0.43 cd | |
| GA3 | 9.10 ± 0.32 ab | 1.35 ± 0.04 e | 75.24 ± 4.85 abc | 45.39 ± 2.59 a | 13.81 ± 0.91 bcd | |
| ABA | 10.56 ± 0.68 b | 1.01 ± 0.02 ab | 87.44 ± 4.65 c | 48.13 ± 3.00 a | 12.90 ± 1.08 abc | |
| SA | 8.56 ± 0.33 a | 1.19 ± 0.05 cd | 78.31 ± 6.03 abc | 49.94 ± 1.39 a | 15.75 ± 1.06 d | |
| GB | 8.35 ± 0.26 a | 1.10 ± 0.03 bc | 72.79 ± 4.63 ab | 48.06 ± 2.91 a | 15.26 ± 1.11 cd | |
| P (T) | * | *** | * | ns | ** | |
| 2016 | Control | 7.51 ± 0.26 ab | 1.19 ± 0.14 a | 75.85 ± 2.12 a | 101.10 ± 2.10 a | 10.79 ± 0.69 a |
| Ca | 7.24 ± 0.40 a | 1.25 ± 0.05 a | 80.84 ± 3.74 ab | 116.49 ± 6.77 ab | 10.87 ± 0.85 a | |
| AN+Ca | 8.76 ± 0.44 c | 1.35 ± 0.04 a | 85.89 ± 5.36 ab | 126.44 ± 3.28 bc | 11.90 ± 0.79 a | |
| GA3+Ca | 7.97 ± 0.78 abc | 1.30 ± 0.07 a | 91.60 ± 1.85 b | 110.58 ± 5.14 ab | 12.90 ± 1.41 a | |
| ABA+Ca | 8.22 ± 0.40 abc | 1.32 ± 0.05 a | 93.92 ± 5.09 b | 121.41 ± 8.12 bc | 13.29 ± 0.54 a | |
| SA+Ca | 8.50 ± 0.43 bc | 1.34 ± 0.05 a | 93.27 ± 6.69 b | 137.61 ± 7.71 c | 11.64 ± 0.92 a | |
| GB+Ca | 8.86 ± 0.39 c | 1.43 ± 0.06 a | 87.67 ± 2.71 ab | 125.77 ± 7.27 bc | 12.32 ± 1.42 a | |
| P (T) | * | ns | * | ** | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, S.; Queirós, F.; Ferreira, H.; Morais, M.C.; Afonso, S.; Silva, A.P.; Gonçalves, B. Foliar Application of Calcium and Growth Regulators Modulate Sweet Cherry (Prunus avium L.) Tree Performance. Plants 2020, 9, 410. https://doi.org/10.3390/plants9040410
Correia S, Queirós F, Ferreira H, Morais MC, Afonso S, Silva AP, Gonçalves B. Foliar Application of Calcium and Growth Regulators Modulate Sweet Cherry (Prunus avium L.) Tree Performance. Plants. 2020; 9(4):410. https://doi.org/10.3390/plants9040410
Chicago/Turabian StyleCorreia, Sofia, Filipa Queirós, Helena Ferreira, Maria Cristina Morais, Sílvia Afonso, Ana Paula Silva, and Berta Gonçalves. 2020. "Foliar Application of Calcium and Growth Regulators Modulate Sweet Cherry (Prunus avium L.) Tree Performance" Plants 9, no. 4: 410. https://doi.org/10.3390/plants9040410
APA StyleCorreia, S., Queirós, F., Ferreira, H., Morais, M. C., Afonso, S., Silva, A. P., & Gonçalves, B. (2020). Foliar Application of Calcium and Growth Regulators Modulate Sweet Cherry (Prunus avium L.) Tree Performance. Plants, 9(4), 410. https://doi.org/10.3390/plants9040410