Effects of Rhizome Integration on the Water Physiology of Phyllostachys edulis Clones Under Heterogeneous Water Stress

Abstract
1. Introduction
2. Results and Discussion
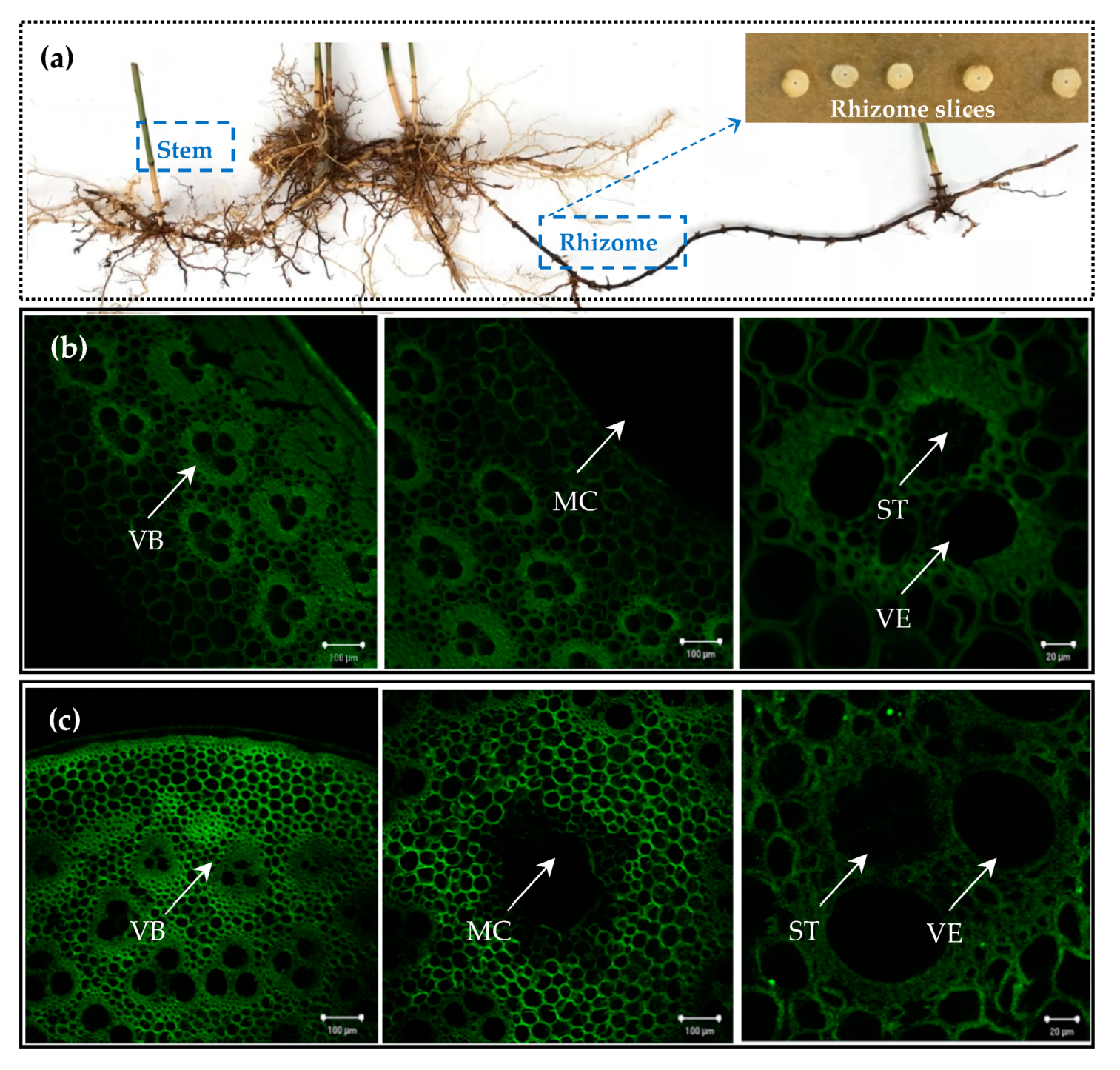
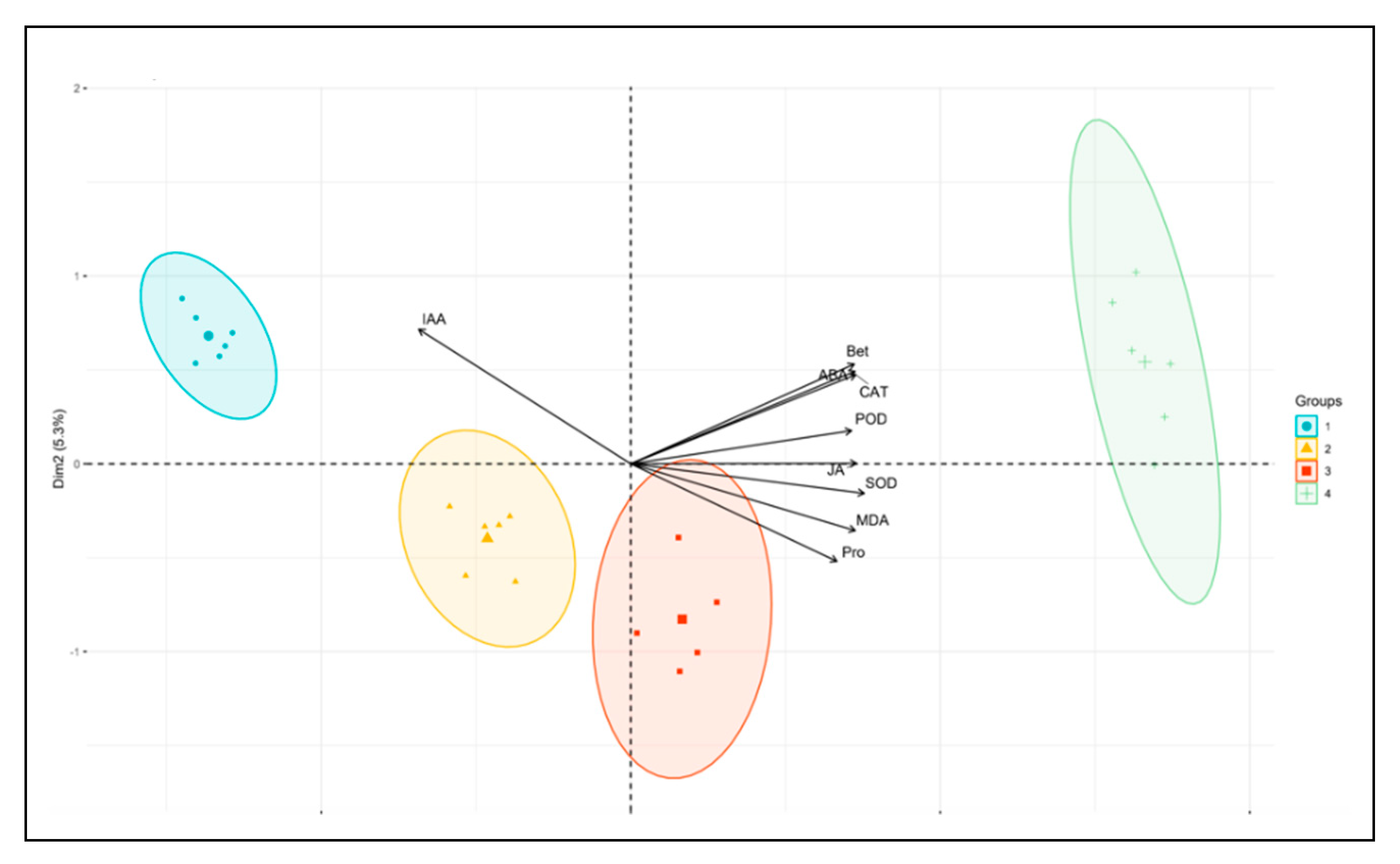
2.1. Morphological Basis of P. edulis Physiology Integration and Analysis of Its Influencing Factor
2.2. Effects of Rhizome Integration on the Related Physiological Indexes of P. edulis Clones under Heterogeneous Water Stress
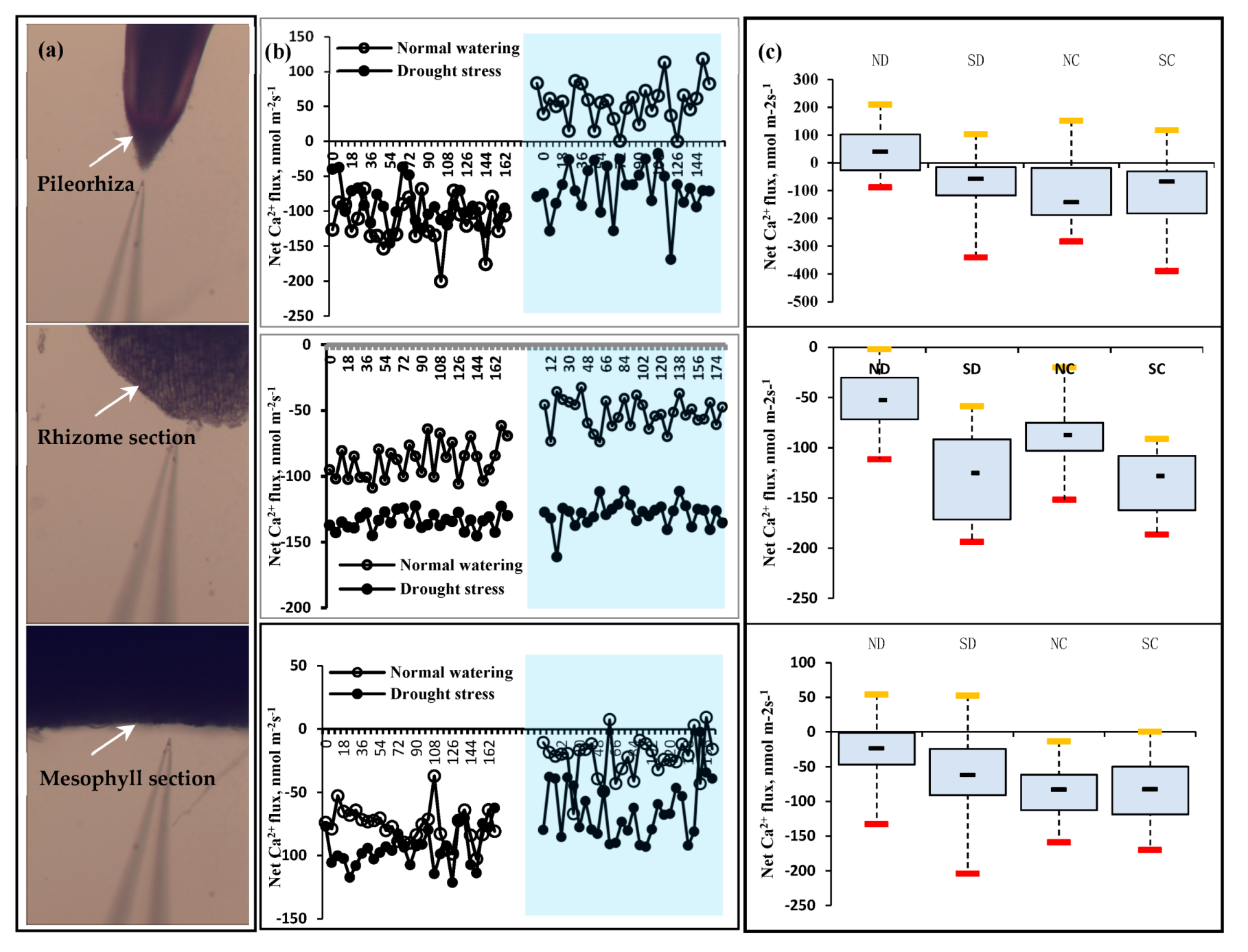
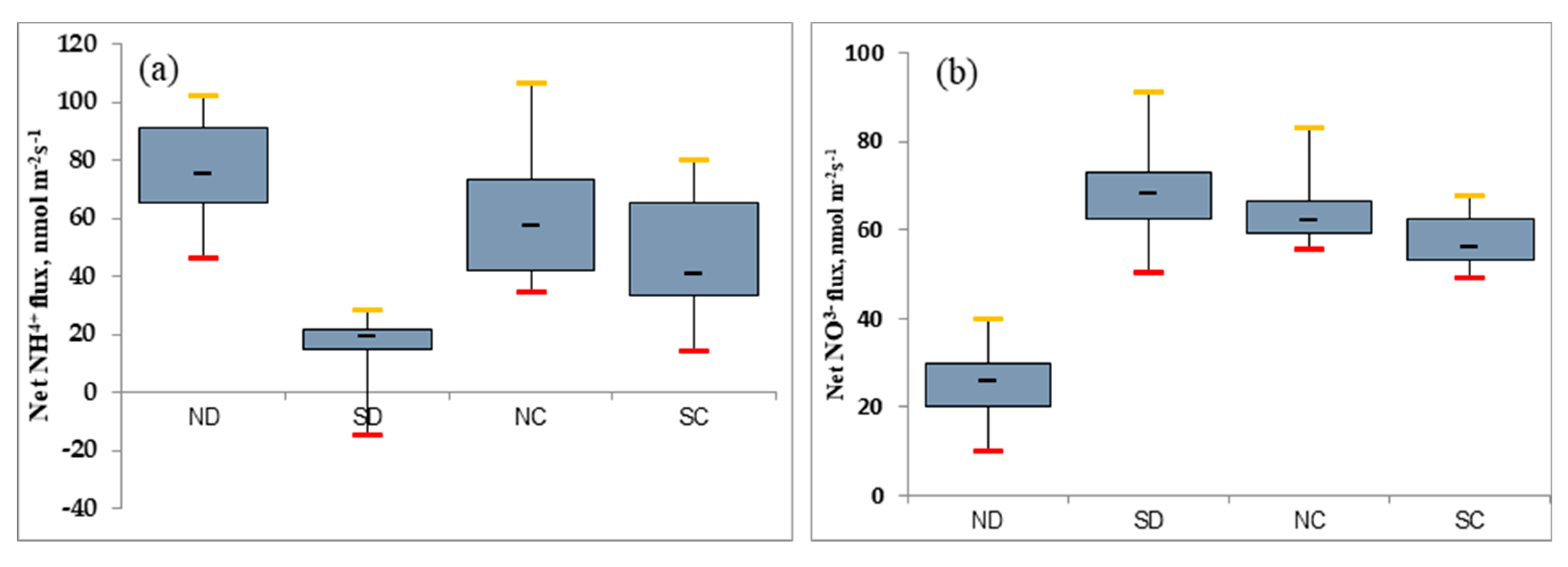
2.3. Effect of the Rhizome on the Ca2+ Flux as the Second Messenger of P. edulis under Heterogeneous Water Stress
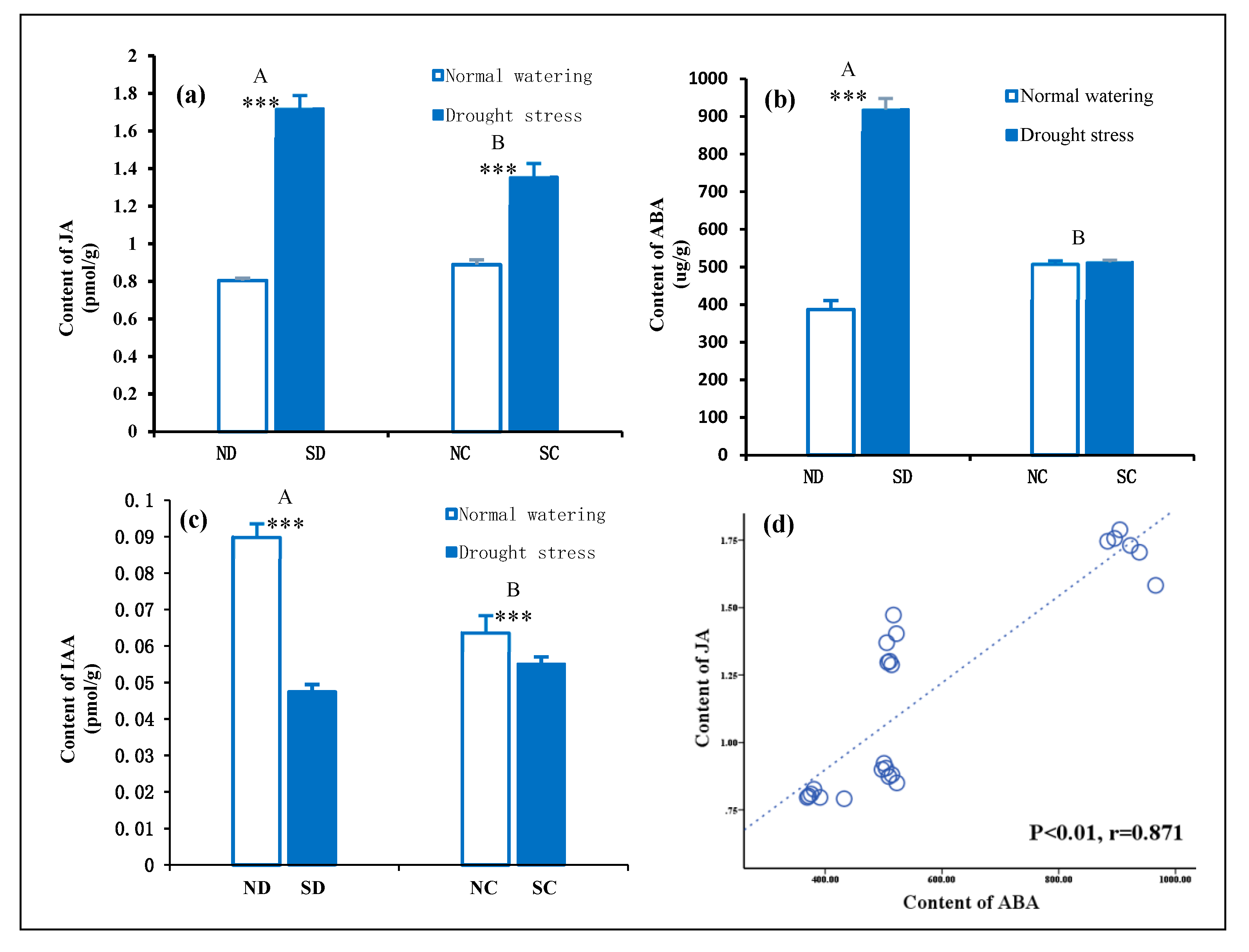
2.4. Effect of Rhizome Integration on Endogenous Hormone Concentration in the Leaves of P. edulis under Heterogeneous Water Stress
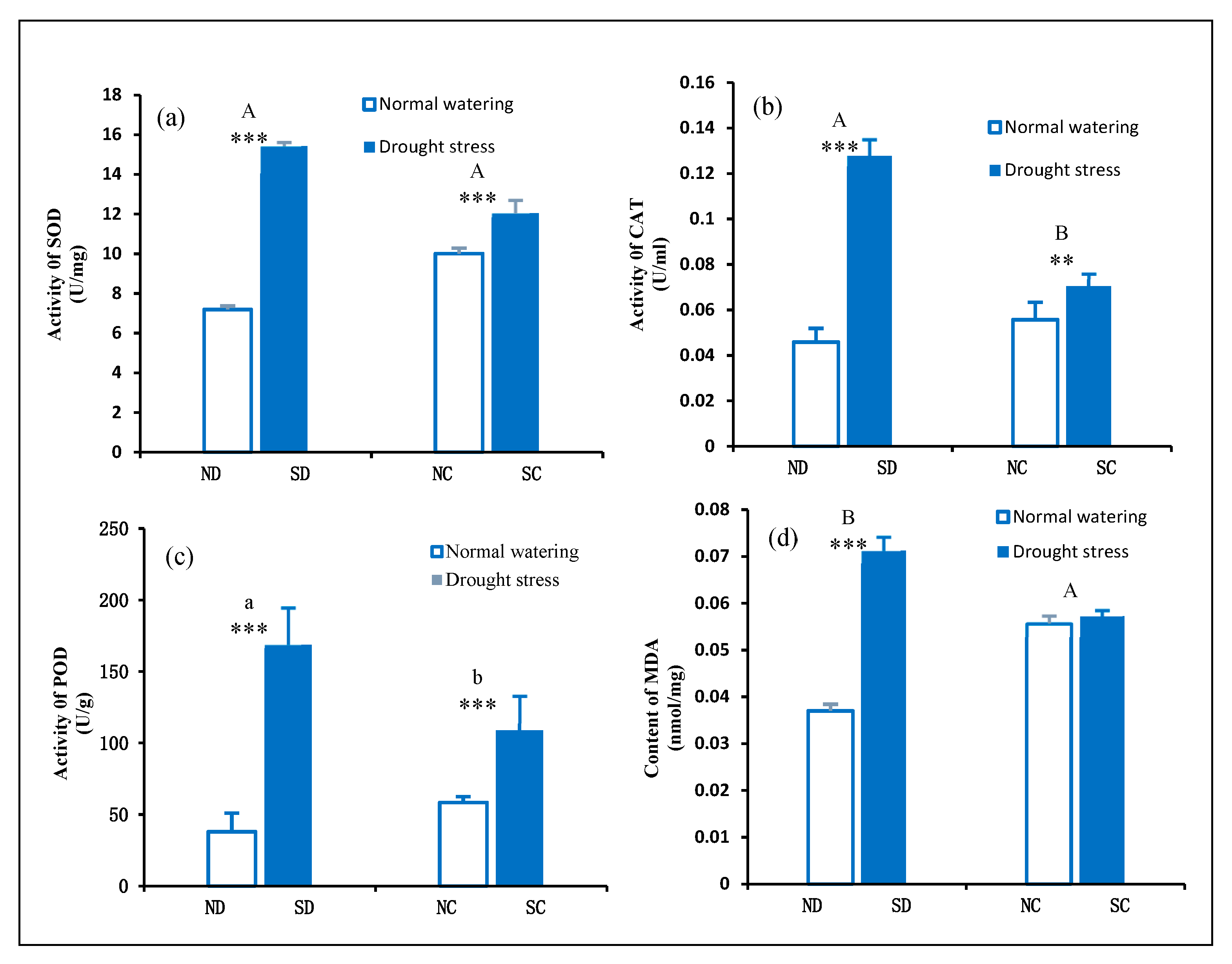
2.5. Effect of Rhizome Severance on the Antioxidant Enzyme System in the Leaves of P. edulis under Heterogeneous Water Stress
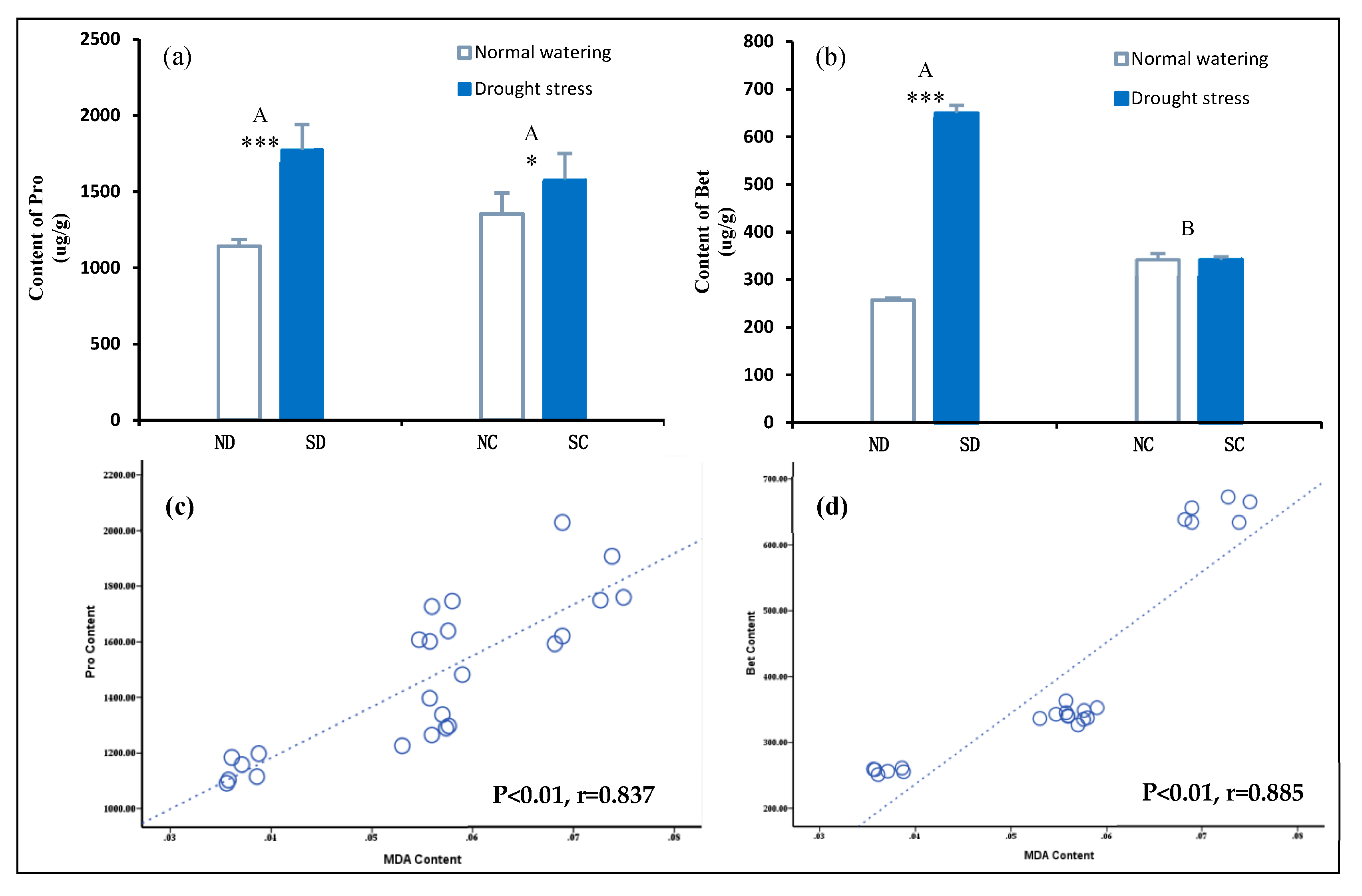
2.6. Effects of the Rhizome on Osmotic Adjustment Substances in P. edulis Leaves under Heterogeneous Water Stress
2.7. Effects of the Rhizome on the Nitrogen Metabolism in the Leaves of P. edulis under Heterogeneous Water Stress
3. Materials and Methods
3.1. Materials
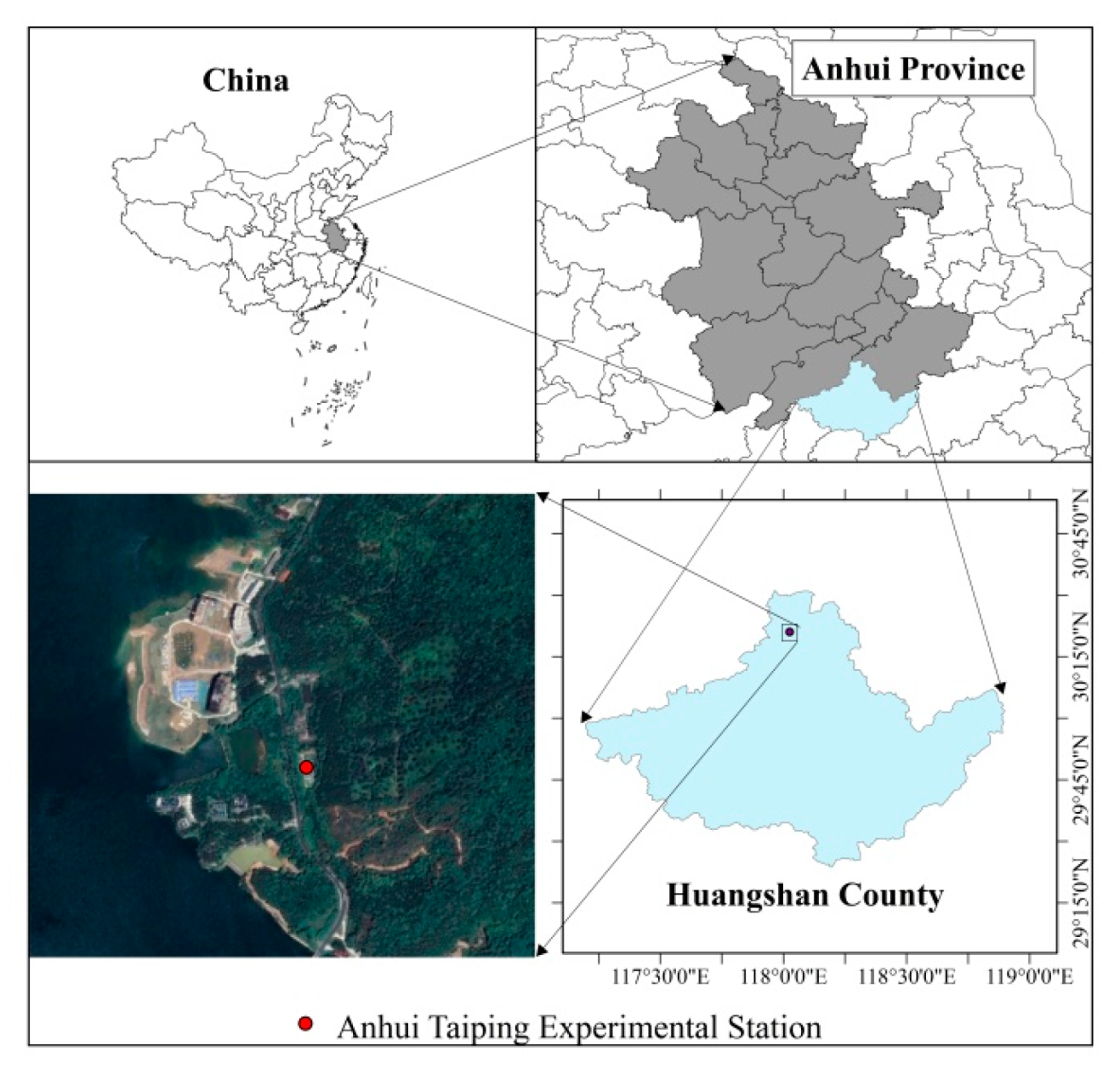
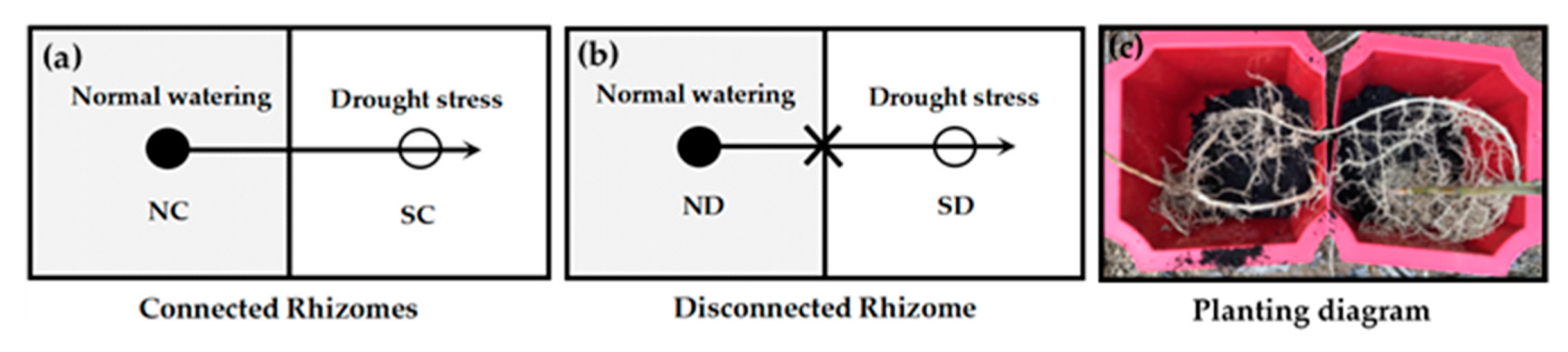
3.2. Experiment Design
3.3. Methods
3.3.1. Laser Confocal Microscopy Luminescence Imaging to Observe the Structural Characteristics of Vascular Bundles of the Stem and Rhizome
3.3.2. Measurement of Ca2+, NH4+, and NO3− Flux
3.3.3. Changes of Hormone, Malondialdehyde (MDA), and Osmotic Adjustment Substance Content of P. edulis under Heterogeneous Drought Stress
3.3.4. The Capacity of Antioxidant Defense Systems Activity of P. edulis under Heterogeneous Drought Stress
3.3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carrão, H.; Naumann, G.; Barbosa, P. Global projections of drought hazard in a warming climate: A prime for disaster risk management. Clim. Dyn. 2018, 50, 2137–2155. [Google Scholar] [CrossRef]
- Wu, S.; Hu, C.; Tan, Q.; Li, L.; Shi, K.; Zheng, Y. Drought stress tolerance mediated by zinc-induced antioxidative defense and osmotic adjustment in cotton (Gossypium Hirsutum ). Acta Physiol. Plant. 2015, 37, 167. [Google Scholar] [CrossRef]
- Haq, T.U.; Ali, A.; Nadeem, S.; Maqbool, M.M.; Ibrahim, M. Performance of canola cultivars under drought stress induced by withholding irrigation at different growth stages. Soil Environ. 2014, 33, 43–50. [Google Scholar]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant. 2016, 2, 201–209. [Google Scholar] [CrossRef]
- Jiang, Z.H.; Fei, B.H.; Fan, S.H. Actively develop bamboo industry and vigorously promote the ecological civilization construction. State Acad. For. Adm. J. 2014, 2, 12–16. [Google Scholar]
- Zhou, G.M.; Meng, C.F.; Jiang, P.K.; Xu, Q.F. Review of Carbon Fixation in Bamboo Forests in China. Bot. Rev. 2011, 77, 262–270. [Google Scholar] [CrossRef]
- Bai, S.; Wang, Y.; Conant, R.T.; Zhou, G.; Xu, Y.; Wang, N.; Fang, F.; Chen, J. Can native clonal moso bamboo encroach on adjacent natural forest without human intervention? Sci. Rep. 2016, 6, 31504. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.L.; Xing, Y.P.; Wei, G.W.; Li, C.Y. Clonal integration facilitates spread of Paspalum paspaloides from terrestrial to cadmium contaminated aquatic habitats. Plant Biol. 2017, 19, 859–867. [Google Scholar] [CrossRef]
- Czarnecka, B. Spatiotemporal Patterns of Genets and Ramets in a Population of Clonal Perennial Senecio rivularis: Plant features and habitat effects. Ann. Bot. Fenn. 2008, 45, 19–32. [Google Scholar] [CrossRef]
- Dong, M. Clonal growth in plants in relation to resource heterogeneity: Foraging behavior. Acta Bot. Sin. 1996, 38, 828–835. [Google Scholar]
- Poor, A.; Hershock, C.; Rosella, K.; Goldberg, D.E. Do Physiological Integration and Soil Heterogeneity Influence the Clonal Growth and Foraging of Schoenoplectus pungens? Plant Ecol. 2005, 181, 45–56. [Google Scholar] [CrossRef][Green Version]
- Sergior, R.; Susana, R.; Eduardo, D.L.P.; Helena, F. Physiological integration increases the survival and growth of the clonal invader Carpobrotus edulis. Biol. Invasions 2010, 12, 1815–1823. [Google Scholar]
- Zhang, Y.; Zhang, Q.; Marek, S. Physiological Integration Ameliorates Negative Effects of Drought Stress in the Clonal Herb Fragaria orientalis. PLoS ONE 2012, 7, e44221. [Google Scholar] [CrossRef]
- Herben, T. Physiological integration affects growth form and competitive ability in clonal plants. Evol. Ecol. 2004, 18, 493–520. [Google Scholar] [CrossRef]
- Ito, R.; Miyafuji, H.; Kasuya, N. Rhizome and root anatomy of moso bamboo (Phyllostachys pubescens) observed with scanning electron microscopy. J. Wood Sci. 2015, 61, 431–437. [Google Scholar] [CrossRef]
- Zhuang, M.H.; Li, Y.C.; Chen, S.L. Advances in the Researches of Bamboo Physiological Integration and Its Ecological Significance. J. Bamboo Res. 2011, 30, 1–9. [Google Scholar]
- Jing, X.; Cai, C.J.; Fan, S.H.; Liu, G.L. Research Advances in Ecological Adaptability of Bamboo under Environmental Stresses. World For. Res. 2018, 31, 36–41. [Google Scholar]
- Saitoh, T.; Nishiwaki, S.A. Importance of Physiological Integration of Dwarf Bamboo to Persistence in Forest Understorey: A Field Experiment. J. Ecol. 2002, 90, 78–85. [Google Scholar] [CrossRef]
- Xui, S.N.; Liu, Y.C.; Liu, Y.H.; Chen, Z.L.; Li, Y.; Zhang, L.H. Physiological integration of growth and photosynthesis of zoysia japonica clonal ramets under nutrient heterogeneity. J. Appl. Ecol. 2018, 29, 811–817. [Google Scholar]
- Qian, Y.Q.; Sun, Z.Y.; Han, L.; Ju, G.S. Photosynthate integration and regulation within clones of buffalograss under heterogeneous water supply. Acta Ecol. Sin. 2010, 30, 3966–3973. [Google Scholar]
- Hu, J.J.; Chen, S.L.; Guo, Z.W.; Yang, Q.P.; Li, Y.C. Effects of Spacer Length on Water Physiological Integration of Indocalamus decorus Ramets under Heterogeneous Water Supply. Acta Bot. Boreali Occident. Sin. 2015, 35, 2532–2541. [Google Scholar]
- Satchivi, N.M.; Stoller, E.W.; Wax, L.M.; Briskin, D.P. A Nonlinear Dynamic Simulation Model for Xenobiotic Transport and Whole Plant Allocation Following Foliar Application. I. Conceptual Foundation for Model Development. Pestic. Biochem. Physiol. 2000, 68, 67–84. [Google Scholar] [CrossRef]
- Liu, Y.H.; Jia, H.S.; Gao, J. Review on researches of photoassimilates partitioning and its models. Acta Ecol. Sin. 2006, 6, 1981–1992. [Google Scholar]
- Mh, C.D.C. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar]
- Sadiqov, S.T.; Akbulut, M.; Ehmedov, V. Role of Ca2+ in drought stress signaling in wheat seedlings. Biochemistry 2002, 67, 491–497. [Google Scholar] [PubMed]
- Ahmad, P.; Bhardwaj, R.; Tuteja, N. Plant Signaling Under Abiotic Stress Environment. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Springer: New York, NY, USA, 2012. [Google Scholar]
- Roberts, A.W.; Haigler, C.H. Rise in chlorotetracycline fluorescence accompanies tracheary element differentiation in suspension cultures of Zinnia. Protoplasm 1998, 152, 37–45. [Google Scholar] [CrossRef]
- Brauer, M.; Zhong, W.J.; Jelitto, T.; Schobert, C.; Sanders, D.; Komor, E. Free calcium ion concentration in the sieve-tube sap of Ricinus communis L. Planta 1998, 206, 103–107. [Google Scholar] [CrossRef]
- Yu, F. Studies on Developmental Biology of Phloem Ganglion in Phyllostachys Edulis; Nanjing Forestry University: Nanjing, China, 2005. [Google Scholar]
- Zhang, Q.X.; Lan, Y.P. Signal Transduction within Plants under Water Stress. J. Shanxi Teach. Univ. 2004, 18, 86–91. [Google Scholar]
- Jacobsen, S.E.; Liu, F.; Jensen, C.R. Does root-sourced ABA play a role for regulation of stomata under drought in quinoa (Chenopodium quinoa Willd). Sci. Hortic. 2009, 122, 281–287. [Google Scholar] [CrossRef]
- Kuromori, T.; Seo, M.; Shinozaki, K. ABA Transport and Plant Water Stress Responses. Trends Plant Sci. 2018, 23, 513–522. [Google Scholar] [CrossRef]
- Ma, H.P.; Liu, Z.M. Transmission and distribution of ABA in seedlings and shoots of Malus micromalus Makino and their relationship with water. Plant Physiol. J. 1998, 24, 253–258. [Google Scholar]
- Chen, J.S.; Lei, N.F.; Liu, Q. Defense signaling among interconnected ramets of a rhizomatous clonal plant, induced by jasmonic-acid application. Acta Oecologica 2011, 37, 355–360. [Google Scholar] [CrossRef]
- Liu, Z.H.; Guo, X.; Gang, W.; Li, G.M. Drought stress and signal transduction of ABA. Chin. Bull. Bot. 2004, 21, 228–234. [Google Scholar]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M. Areb1 is a transcription activator of novel abre-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell 2005, 17, 3470. [Google Scholar] [CrossRef]
- Li, C.J. Autoinhibition in polar transport of indoleacetic acid. Acta Bot. Sin. 1997, 39, 737–741. [Google Scholar]
- Xu, S.C. The Roles of Ca2+-Dependent and Calmodulin Stimulated Protein Kinase in Abscisic Acid-Induced Antioxidant Defense in Leaves of Maize Plants. Ph.D. Thesis, Nanjing Agricultural University, Najing, China, 2009. [Google Scholar]
- Li, X.; Yue, H.; Huang, L.Q.; Guo, P.L. Study on the Reaction Regular of Plant Antioxidant Enzymes under Environmental Stress; Chinese Pharmaceutical Congress: Beijing, China, 2010. [Google Scholar]
- Schuller, D.J.; Ban, N.; Huystee, R.B.; Mcpherson, A.; Poulos, T.L. The crystal structure of peanut peroxidase. Structure 1996, 4, 311. [Google Scholar] [CrossRef]
- Xu, S.N.; Li, Y.; Chen, Z.L.; Zhong, L.H. Physiological integration of antioxidant enzymes and malondialdehyde in connected and disconnected clonal ramet under nutrient heterogeneity. Pratacult. Sci. 2018, 35, 13. [Google Scholar]
- Qin, W.U.; Zhang, G.; Pei, B. Physiological and biochemical responses to different soil drought stress in three tree species. Acta Ecol. Sin. 2013, 33, 3648–3656. [Google Scholar] [CrossRef]
- Wang, H.C. Effect of Light on Proline Synthesis under Drought Stress and Its Relationship with ABA. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2013. [Google Scholar]
- An, H.; ShangGuan, Z.P. The nitrogen cycling of plants and it’s physiological mechanism of root-zone environment. Res. Soil Water Conserv. 2006, 13, 83–85. [Google Scholar]
- Xu, Y.; Sun, T.; Yin, L.P. Application of Non-invasive Microsensing System to Simultaneously Measure Both H+, and O2 Fluxes Around the Pollen Tube. J. Integr. Plant Biol. 2006, 48, 823–831. [Google Scholar] [CrossRef]
- Xin, Z.Y.; Zhou, X.; Pilet, P.E. Level changes of jasmonic, abscisic, and indole-3yl-acetic acids in maize under desiccation stress. J. Plant Physiol. 1997, 151, 120–124. [Google Scholar] [CrossRef]
- Engvall, E. Enzyme immunoassay ELISA and EMIT. Methods Enzym. 1980, 70, 419–439. [Google Scholar]
- Shinagawa, A.; Suzuki, T.; Konosu, S. The Role of Free Amino Acids and Betaines in Intracellular Osmoregulation of Marine Sponges. Nippon Suisan Gakkaishi 1992, 58, 1717–1722. [Google Scholar] [CrossRef]
- Lin, S.Y.; Wu, J.X. Discussion on the Determination of SOD by Tetrazolium Blue (NBT) Method. Strait Pharm. J. 2009, 21, 43–45. [Google Scholar]
- Liu, S.Y.; An, W.; Xu, X.W.; Liu, H.Z. Determination of POD, CAT and PAL enzyme activity in leaves of Hippophae rhamnoides. J. Jilin Agric. Univ. 2009, 31, 584–586. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| W | R | W × R | |
|---|---|---|---|
| SOD activity | 2947.21 *** | 4.11 ns | 979.94 *** |
| CAT activity | 322.15 *** | 77.69 *** | 155.23 *** |
| POD activity | 137.26 *** | 6.54 ns | 26.91 *** |
| JA concentration | 999.70 *** | 40.41 *** | 105.17 *** |
| ABA concentration | 1076.62 *** | 303.77 *** | 1040.69 *** |
| IAA concentration | 553.84 *** | 73.67 *** | 243.18 *** |
| Pro concentration | 56.84 *** | 0.02 ns | 12.87 ** |
| Bet concentration | 1860.51 *** | 605.20 *** | 1898.34 *** |
| MDA concentration | 518.35 *** | 8.93 ** | 430.09 *** |
| Ion Species Forflux Testing | Test Position Site (μm) | Measuring Solutions Constitution | |||
|---|---|---|---|---|---|
| Pileorhiza | Rhizome | Mesophyll | Concentration | Constitution | |
| Ca2+ | 0 μm from the root apex | Cross-section | Cross-section | 0.1 mM 0.1 mM | NH4NO3 CaCl2 |
| NH4+ | - | - | Cross-section | 0.1 mM 0.1 mM | NH4NO3 CaCl2 |
| NO3− | - | - | Cross-section | 0.1 mM 0.1 mM | NH4NO3 CaCl2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, X.; Cai, C.; Fan, S.; Liu, G.; Wu, C.; Chen, B. Effects of Rhizome Integration on the Water Physiology of Phyllostachys edulis Clones Under Heterogeneous Water Stress. Plants 2020, 9, 373. https://doi.org/10.3390/plants9030373
Jing X, Cai C, Fan S, Liu G, Wu C, Chen B. Effects of Rhizome Integration on the Water Physiology of Phyllostachys edulis Clones Under Heterogeneous Water Stress. Plants. 2020; 9(3):373. https://doi.org/10.3390/plants9030373
Chicago/Turabian StyleJing, Xiong, Chunju Cai, Shaohui Fan, Guanglu Liu, Changming Wu, and Benxue Chen. 2020. "Effects of Rhizome Integration on the Water Physiology of Phyllostachys edulis Clones Under Heterogeneous Water Stress" Plants 9, no. 3: 373. https://doi.org/10.3390/plants9030373
APA StyleJing, X., Cai, C., Fan, S., Liu, G., Wu, C., & Chen, B. (2020). Effects of Rhizome Integration on the Water Physiology of Phyllostachys edulis Clones Under Heterogeneous Water Stress. Plants, 9(3), 373. https://doi.org/10.3390/plants9030373