Tetraploid Induction by Colchicine Treatment and Crossing with a Diploid Reveals Less-Seeded Fruit Production in Pointed Gourd (Trichosanthes dioica Roxb.)

 ,
,
Abstract
1. Introduction
2. Results
2.1. Effect of Colchicine Treatments on the Germination Rate and Surviving Seedling Rate
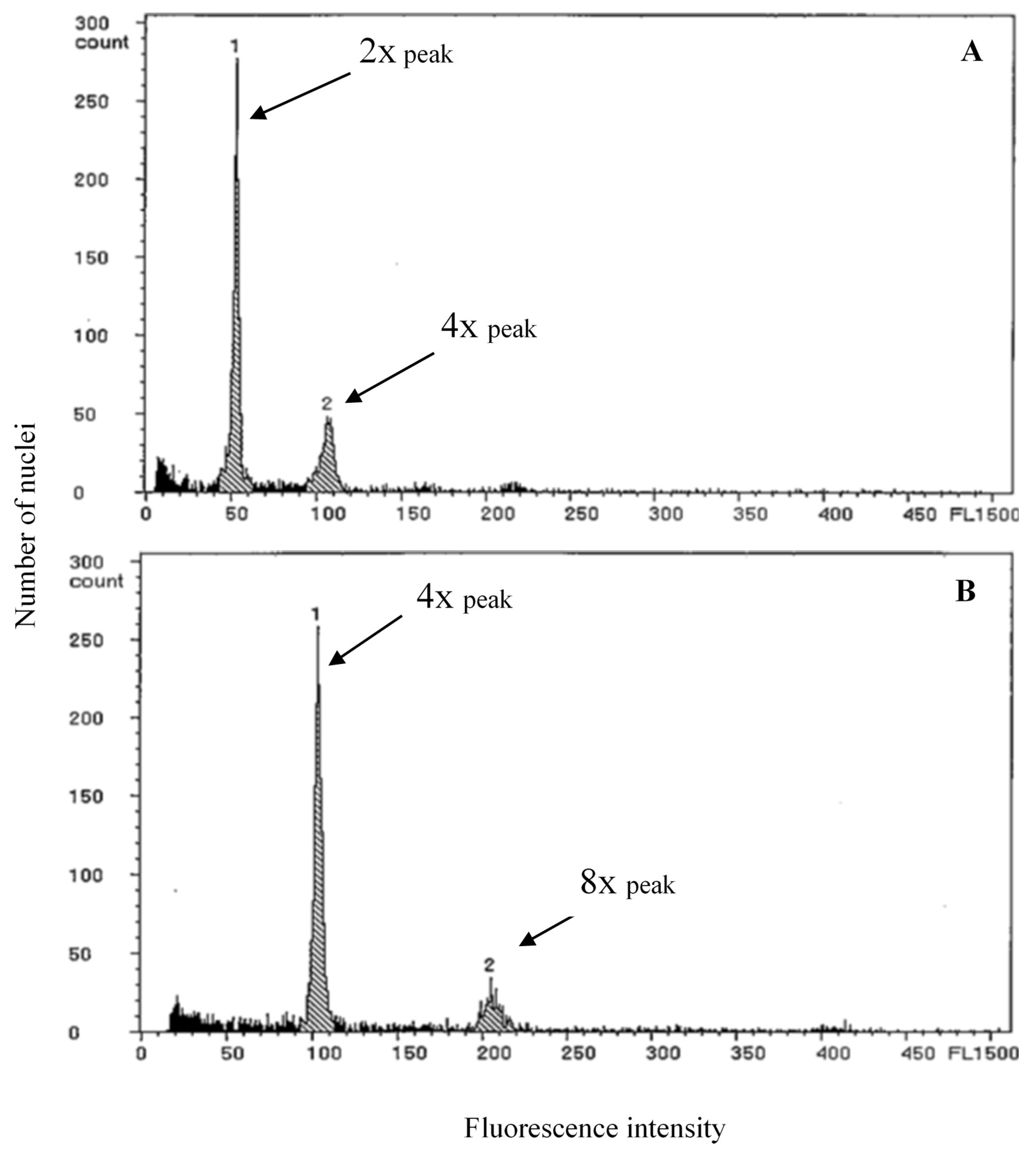
2.2. Confirmation of Ploidy Level
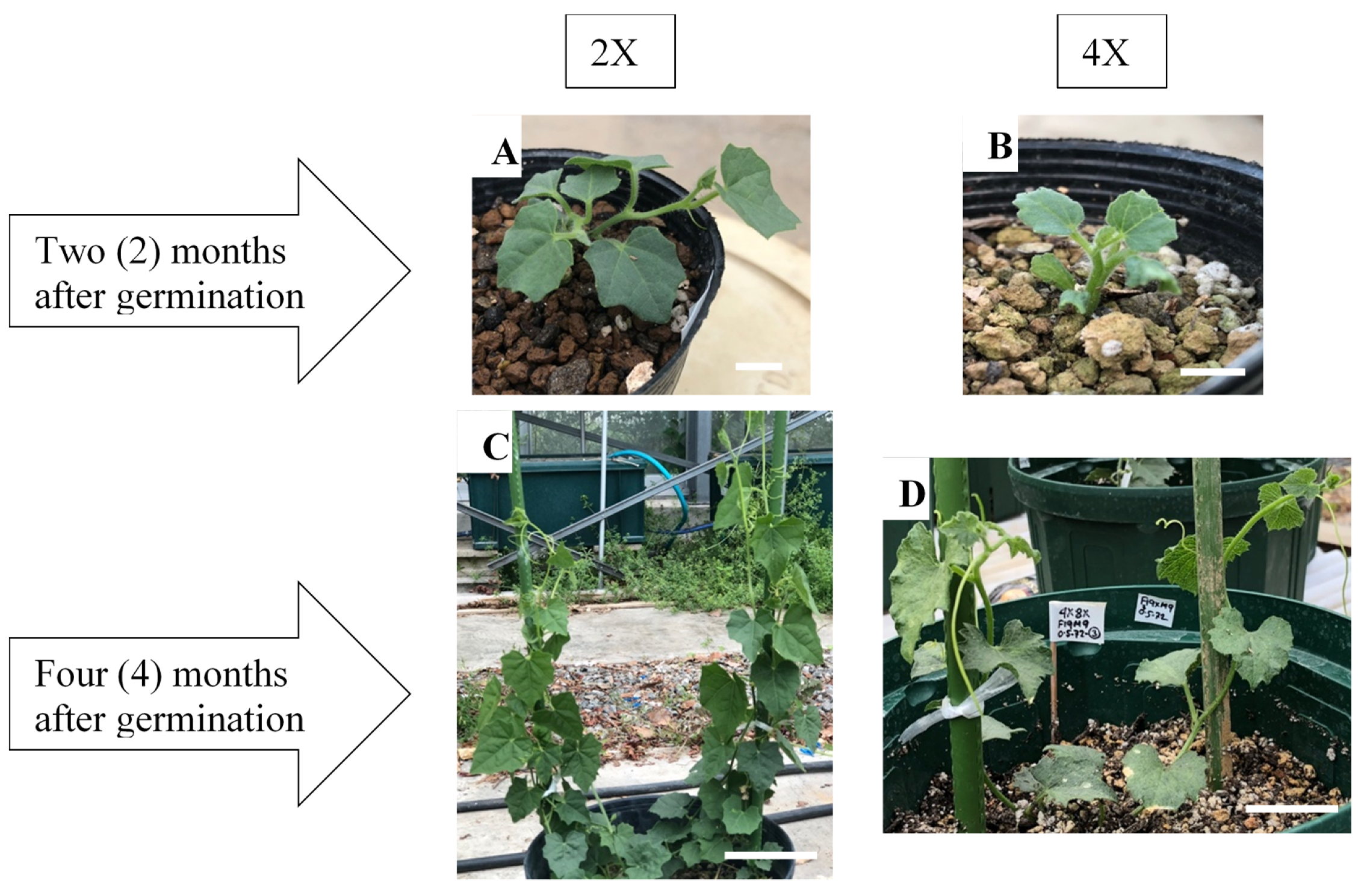
2.3. Morphological Characteristics of Diploid and Colchicine-Induced Tetraploid Plants
3. Discussion
4. Materials and Methods
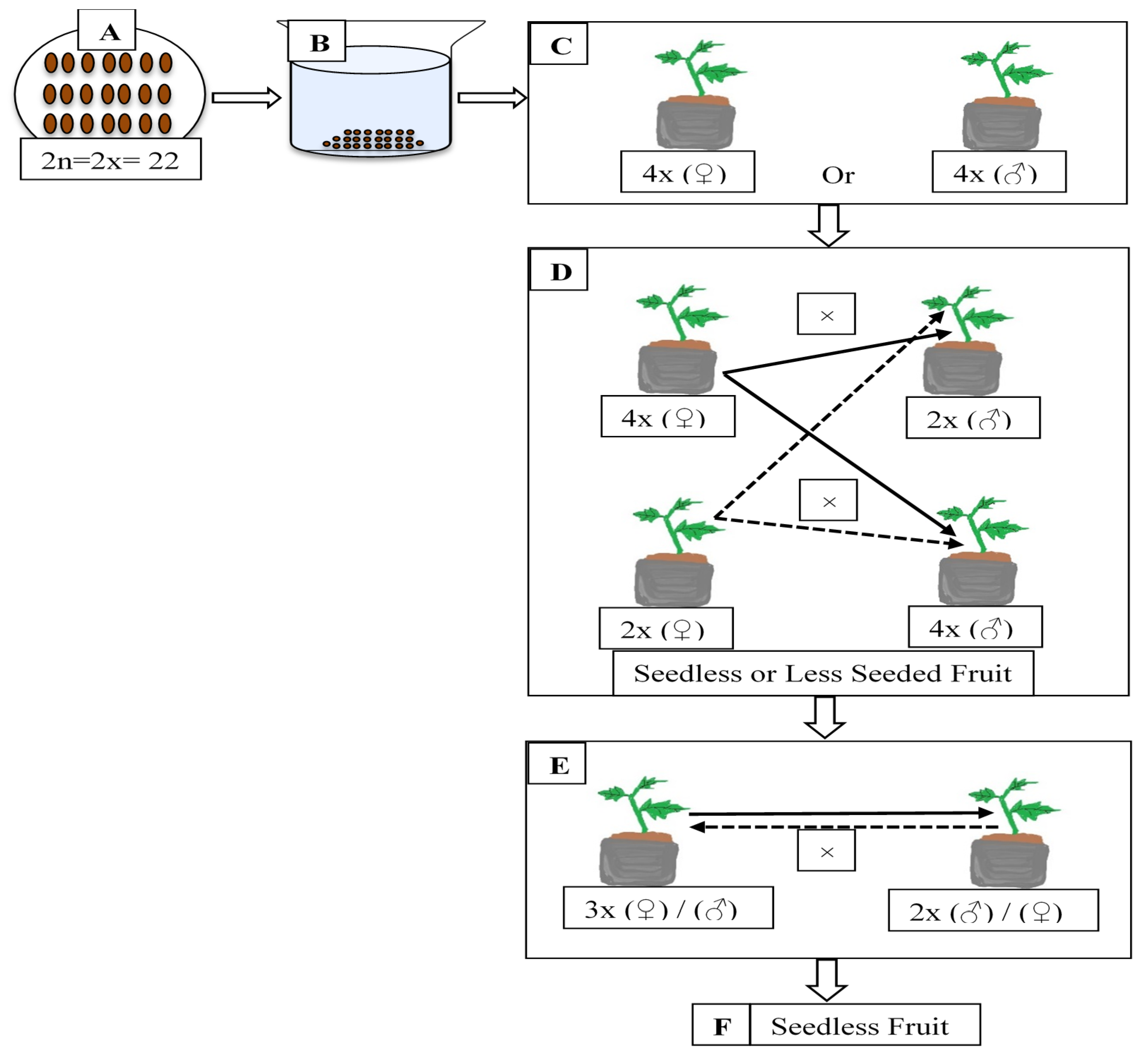
4.1. Colchicine Treatment
4.2. Confirmation of Ploidy Level
4.3. Vegetative and Reproductive Traits
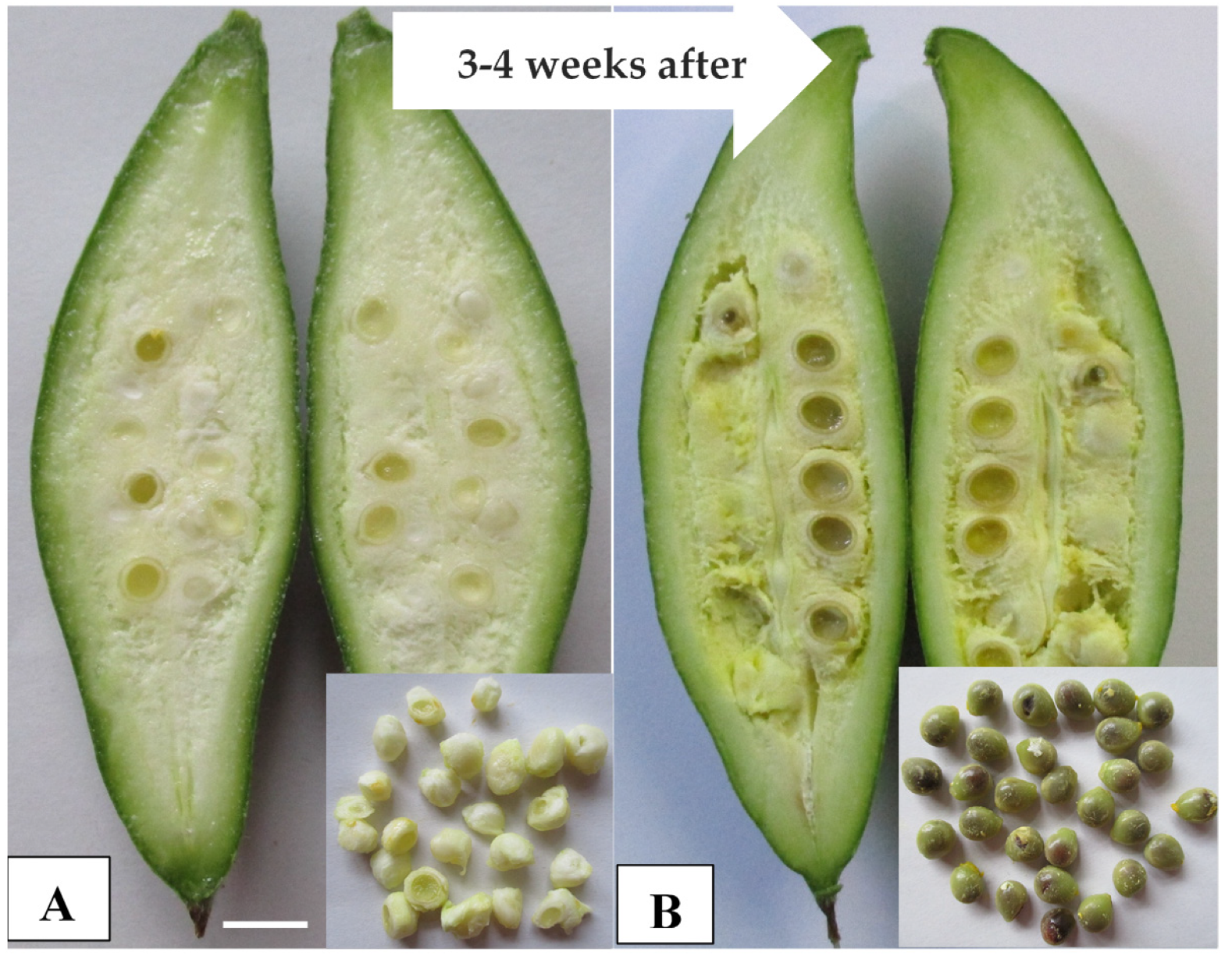
4.4. Crosspollination of Diploids with Colchicine-Induced Tetraploids
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sinha, S.; Guha, A.; Sinha, R.K. Karyotype and sex expression in Trichosanthes dioica. Cytologia 2003, 68, 357–361. [Google Scholar] [CrossRef]
- Hassan, J.; Miyajima, I. Induction of Parthenocarpy in Pointed Gourd (Trichosanthes dioica Roxb.) by Application of Plant Growth Regulators. J. Hortic. Plant Res. 2019, 8, 12–21. [Google Scholar] [CrossRef]
- Dubey, K.C.; Nair, P.K.R. Induced parthenocarpic fruit-set in pointed gourd (Trichosanthes dioica Roxb.). Indian J. Agric. Sci. 1972, 42, 765–768. [Google Scholar]
- Kim, I.S.; Okubo, H.; Fujieda, K. Endogenous levels of IAA in relation to parthenocarpy in cucumber (Cucumis sativus L.). Sci. Hortic. 1992, 52, 1–8. [Google Scholar] [CrossRef]
- Rasul, M.G.; Mian, M.A.K.; Cho, Y.; Ozaki, Y.; Okubo, H. Application of plant growth regulators on the parthenocarpic fruit development in teasle gourd (kakrol, Momordica dioica Roxb.). J. Fac. Agric. Kyushu Univ. 2008, 53, 39–42. [Google Scholar]
- Chen, L.P.; Wang, Y.J.; Zhao, M. In vitro induction and characterization of tetraploid Lychnis senno Siebold et Zucc. HortScience 2006, 41, 759–761. [Google Scholar] [CrossRef]
- Liu, Z.; Min, Z.; Sun, X.; Cheng, J.; Hu, Y. Study on induction and characterization of tetraploid plants in pumpkin. J. North China Agric. Univ. 2015, 30, 125–129. [Google Scholar]
- Maynard, D.N. An introduction to the watermelon. In Watermelons: Characteristics, Production and Marketing; ASHS Press: Alexandria, Egypt, 2001; pp. 9–20. [Google Scholar]
- Kazi, N.A. Polyploidy in vegetables. J. Glob. Biosci. 2015, 4, 1774–1779. [Google Scholar]
- Hazra, P. Induced polyploidy as a breeding approach in pointed gourd. J. Breed. Genet. 2001, 33, 47–48. [Google Scholar]
- Allum, J.F.; Bringloe, D.H.; Roberts, A.V. Chromosome doubling in a Rosa rugosa Thunb. Hybrid by exposure of in vitro nodes to oryzalin: The effects of node length, oryzalin concentration and exposure time. Plant Cell Rep. 2007, 26, 1977–1984. [Google Scholar] [CrossRef]
- Kihara, H. Tetraploid watermelons. Proc. Am. Soc. Hort. Sci. 1951, 58, 217–230. [Google Scholar]
- Zlesak, D.C.; Thill, C.A.; Anderson, N.O. Trifluralin-mediated polyploidization of Rosa chinensis minima (Sims) Voss seedlings. Euphytica 2005, 141, 281–290. [Google Scholar] [CrossRef]
- Dermen, H.; Emsweller, S.L. The use of colchicine in plant breeding. In Crops Research; Agricultural Research Service, U.S. Department of Agriculture, FEDLINK, USDA-arsseries: Washington, DC, USA, 1961; pp. 1–10. [Google Scholar]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Addison-Wesley: London, UK, 1971. [Google Scholar]
- Jaskani, M.J.; Kwon, S.W.; Kim, D.H. Comparative study on vegetative, reproductive and qualitative traits of seven diploid and tetraploid watermelon lines. Euphytica 2005, 145, 259–268. [Google Scholar] [CrossRef]
- Jaskani, M.J.; Khan, I.A.; Husnain, S. Morphological description of citrus colchiploids. Proc. Int. Citrus Congr. 1996, 1, 130–132. [Google Scholar]
- Thao, N.T.P.; Ureshino, K.; Miyajima, I.; Ozaki, Y.; Okubo, H. Induction of tetraploids in ornamental Alocasia through colchicine and oryzaling treatments. Plant Cell Tissue Organ Cult. 2003, 72, 19–25. [Google Scholar] [CrossRef]
- Rose, J.B.; Kubba, J.; Tobutt, K.R. Chromosome doubling in sterile Syringa vulgaris × S. pinnatifolia hybrids by in vitro culture of nodal explants. Plant Cell Tissue Organ Cult. 2000, 63, 127–132. [Google Scholar] [CrossRef]
- Wehner, T.C.; Shetty, N.V.; Elmstrom, G.W. Breeding and seed production. In Watermelons: Characteristics, Production and Marketing; Maynard, D.N., Ed.; ASHS Press: Alexandria, Egypt, 2001; pp. 27–73. [Google Scholar]
- Tiwari, S.; Spielman, M.; Schulz, R.; Oakey, R.J.; Kelsey, G.; Salazar, A.; Zhang, K.; Pennell, R.; Scott, R.J. Transcriptional profiles underlying parent-of-origin effects in seeds of Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 70. [Google Scholar] [CrossRef]
- Stebbins, G.L., Jr.; Polyploidy, I. Occurrences and nature of polyploid types. In Variation and Evolution in Plants; Dunn, L.C., Ed.; Columbia Univ. Press: New York, NY, USA, 1950; pp. 298–339. [Google Scholar]
- Burton, T.L.; Husband, B.C. Fitness differences among diploids, tetraploids, and their triploid progeny in Chamerion angustifolium: Mechanism of inviability and implications for polyploid evolution. Evolution 2000, 54, 1182–1191. [Google Scholar] [CrossRef]
- Walters, S.A.; Wehner, T.C. Incompatibility in diploid and tetraploid crosses of Cucumis sativus and Cucumis metuliferus. Euphytica 2002, 128, 371–374. [Google Scholar] [CrossRef]
- Wall, J.R. Use of marker genes in producing triploid watermelon. Proc. Am. Soc. Hort. Sci. 1960, 76, 577–581. [Google Scholar]
- Earhart, D.R.; Dainello, F.J.; Baker, M.L. Seedless (Triploid) watermelon evaluations for East Texas. Prog. Rep. 1994, 9, 5148. [Google Scholar]
- Gray, D.T.; Elmstrom, G.W. Process for Accelerated Production of Triploid Seed, for Seedless Watermelon Cultivar. U.S. Patent US5007198, 16 April 1991. [Google Scholar]
- Hassan, J.; Miyajima, I. Morphological and Ecological Characteristics of Pointed Gourd (Trichosanthes dioica Roxb.). J. Fac. Agric. Kyushu Univ. 2019, 64, 183–189. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2010. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Colchicine Treatments | PGF02 × PGM03 | PGF17 × PGM08 | PGF18 × PGM08 | PGF19 × PGM09 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Conc. (%) | Duration (h) | Germinated Seeds (%) | Surviving Seedling (%) z | Germinated Seeds (%) | Surviving Seedling (%) | Germinated Seeds (%) | Surviving Seedling (%) | Germinated Seeds (%) | Surviving Seedling (%) |
| 0.05 | 24 | 1 (4.0) | 1 (4.0) | 3 (12.0) | 2 (8.0) | 2 (8.0) | 2 (8.0) | 1 (4.0) | 1 (4.0) |
| 48 | 5 (20.0) | 5 (20.0) | 1 (4.0) | 1 (4.0) | 3 (12.0) | 1 (4.0) | 2 (8.0) | 2 (8.0) | |
| 72 | 12 (48.0) | 9 (36.0) | 4 (16.0) | 4 (16.0) | 0 (0.0) | 0 (0.0) | 9 (36.0) | 9 (36.0) | |
| 0.1 | 24 | 1 (4.0) | 1 (4.0) | 2 (8.0) | 1 (4.0) | 5 (20.0) | 5 (20.0) | 7 (28.0) | 7 (28.0) |
| 48 | 5 (20.0) | 5 (20.0) | 0 (0.0) | 0 (0.0) | 4 (16.0) | 4 (16.0) | 7 (28.0) | 6 (24.0) | |
| 72 | 8 (32.0) | 6 (24.0) | 3 (12.0) | 3 (12.0) | 2 (8.0) | 2 (8.0) | 5 (20.0) | 4 (16.0) | |
| 0.5 | 24 | 1 (4.0) | 1 (4.0) | 5 (20.0) | 5 (20.0) | 2 (8.0) | 2 (8.0) | 3 (12.0) | 2 (8.0) |
| 48 | 4 (16.0) | 3 (12.0) | 1 (4.0) | 1 (4.0) | 5 (20.0) | 2 (8.0) | 5 (20.0) | 5 (20.0) | |
| 72 | 5 (20.0) | 4 (16.0) | 2 (8.0) | 2 (8.0) | 2 (8.0) | 1 (4.0) | 6 (24.0) | 5 (20.0) | |
| Total | 42 | 35 | 21 | 19 | 25 | 19 | 45 | 41 | |
| Colchicine Treatments | PGF02 × PGM03 | PGF17 × PGM08 | PGF18 × PGM08 | PGF19 × PGM09 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Conc. (%) | Duration (h) | Ploidy Levels z | Ploidy Levels | Ploidy Levels | Ploidy Levels | ||||||||
| 2x | 4x | 2x + 4x | 2x | 4x | 2x + 4x | 2x | 4x | 2x + 4x | 2x | 4x | 2x + 4x | ||
| 0.05 | 24 | 1 | 0 | 0 | 2 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 |
| 48 | 4 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | |
| 72 | 8 | 0 | 1 | 4 | 0 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | |
| 0.1 | 24 | 1 | 0 | 0 | 1 | 0 | 0 | 5 | 0 | 0 | 7 | 0 | 0 |
| 48 | 4 | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 1 | 6 | 0 | 0 | |
| 72 | 3 | 0 | 3 | 2 | 0 | 1 | 1 | 0 | 1 | 4 | 0 | 0 | |
| 0.5 | 24 | 1 | 0 | 0 | 5 | 0 | 0 | 2 | 0 | 0 | 2 | 0 | 0 |
| 48 | 3 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 2 | 3 | 1 | 1 | |
| 72 | 3 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 2 | 3 | |
| Total | 28 | 0 | 7 | 16 | 0 | 3 | 14 | 0 | 5 | 34 | 3 | 4 | |
| Ploidy Level | No. of Evaluated Plants | Leaf Length (cm) | Leaf Diameter (cm) | Internode Length (cm) | Internode Diameter (mm) |
|---|---|---|---|---|---|
| Diploid (2x) | 3 | 5.6 ± 0.2 b z | 6.5 ± 0.1 b | 9.9 ± 1.0 ns | 2.6 ± 0.1 ns |
| Tetraploid (4x) | 3 | 8.7 ± 0.1 a | 9.7 ± 0.1 a | 8.6 ± 0.5 | 3.6 ± 0.5 |
| Ploidy Levels z | No. of Days to First Flowering | Node No. of First Flowering | Flowers No./ Plant y | Ovary Length (mm) | Ovary Diameter (mm) | Stalk Length (mm) | Petal Length (mm) | Petal Diameter (mm) |
|---|---|---|---|---|---|---|---|---|
| 2x | 74.3 ± 1.5 b x | 12.7 ± 1.1 b | 51.0 ± 2.0 a | 15.5 ± 1.1 b | 5.4 ± 0.6 b | 28.5 ± 0.9 b | 19.5 ± 0.8 b | 6.3 ± 0.6 b |
| 4x | 129.7 ± 3.2 a | 30.3 ± 2.3 a | 31.7 ± 2.5 b | 29.0 ± 1.2 a | 10.4 ± 1.0 a | 31.1 ± 0.8 a | 33.0 ± 1.2 a | 17.6 ± 1.0 a |
| Ploidy Levels z | Stalk Length (mm) | Stalk Diameter (mm) | Anther Length (mm) | Anther Diameter (mm) | No. of Anthers | Petal Length (mm) | Petal Diameter (mm) |
|---|---|---|---|---|---|---|---|
| 2x | 17.5 ± 0.9 b y | 3.6 ± 0.6 b | 3.1 ± 0.1 b | 5.1 ± 0.2 b | 3.0 ± 0.0 ns | 16.1 ± 0.6 b | 5.6 ± 0.1 b |
| 4x | 45.8 ± 1.2 a | 6.0 ± 0.9 a | 4.5 ± 0.2 a | 9.4 ± 0.3 a | 3.0 ± 0.0 | 24.5 ± 0.5 a | 14.1 ± 0.3 a |
| Cross Combination w (Seed Parent × Pollen Parent) | No. of Flowers Pollinated/Repetition z | No. of Flowers that Set Fruits | Fruit Set Rate (%) | Fruit Length (cm) | Fruit Diameter (cm) | Fruit Weight (g) |
|---|---|---|---|---|---|---|
| 2x × 2x | 5 | 5.0 ± 0.0 a y | 100.0 ± 00.0 a | 11.0 ± 0.9 a | 3.3 ± 0.0 a | 45.7 ± 2.3 a |
| 2x × 4x | 5 | – x | – | – | – | – |
| 4x × 4x | 5 | 0.7 ± 0.6 b | 13.3 ± 11.5 b | 2.9 ± 0.1 c | 0.9 ± 0.8 b | 4.3 ± 3.7 c |
| 4x × 2x | 5 | 4.7 ± 0.5 a | 93.3 ± 11.5 a | 8.8 ± 0.5 b | 3.3 ± 0.2 a | 33.4 ± 0.9 b |
| Cross Combination z (Seed Parent × Pollen Parent) | No. of Evaluated Fruits y | No. of Developed Seeds | No. of Aborted Seeds | Seed Diameter (mm) | 100 Seeds Weight (g) | Seed Germination (%) |
|---|---|---|---|---|---|---|
| 2x × 2x | 3 | 26.4 ± 0.9 a x | 1.3 ± 0.5 b | 5.5 ± 0.1 b | 6.4 ± 0.3 b | 100.0 |
| 2x × 4x | 3 | – w | – | – | – | – |
| 4x × 4x | 3 | 0.0 ± 0.0 b | 1.0 ± 0.9 b | – | – | – |
| 4x × 2x | 3 | 1.8 ± 0.0 b | 2.7 ± 0.3 a | 7.8 ± 0.2 a | 12.4 ± 0.6 a | – |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, J.; Miyajima, I.; Ozaki, Y.; Mizunoe, Y.; Sakai, K.; Zaland, W. Tetraploid Induction by Colchicine Treatment and Crossing with a Diploid Reveals Less-Seeded Fruit Production in Pointed Gourd (Trichosanthes dioica Roxb.). Plants 2020, 9, 370. https://doi.org/10.3390/plants9030370
Hassan J, Miyajima I, Ozaki Y, Mizunoe Y, Sakai K, Zaland W. Tetraploid Induction by Colchicine Treatment and Crossing with a Diploid Reveals Less-Seeded Fruit Production in Pointed Gourd (Trichosanthes dioica Roxb.). Plants. 2020; 9(3):370. https://doi.org/10.3390/plants9030370
Chicago/Turabian StyleHassan, Jahidul, Ikuo Miyajima, Yukio Ozaki, Yuki Mizunoe, Kaori Sakai, and Wasimullah Zaland. 2020. "Tetraploid Induction by Colchicine Treatment and Crossing with a Diploid Reveals Less-Seeded Fruit Production in Pointed Gourd (Trichosanthes dioica Roxb.)" Plants 9, no. 3: 370. https://doi.org/10.3390/plants9030370
APA StyleHassan, J., Miyajima, I., Ozaki, Y., Mizunoe, Y., Sakai, K., & Zaland, W. (2020). Tetraploid Induction by Colchicine Treatment and Crossing with a Diploid Reveals Less-Seeded Fruit Production in Pointed Gourd (Trichosanthes dioica Roxb.). Plants, 9(3), 370. https://doi.org/10.3390/plants9030370