Natural Variation in Volatile Emissions of the Invasive Weed Calluna vulgaris in New Zealand

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
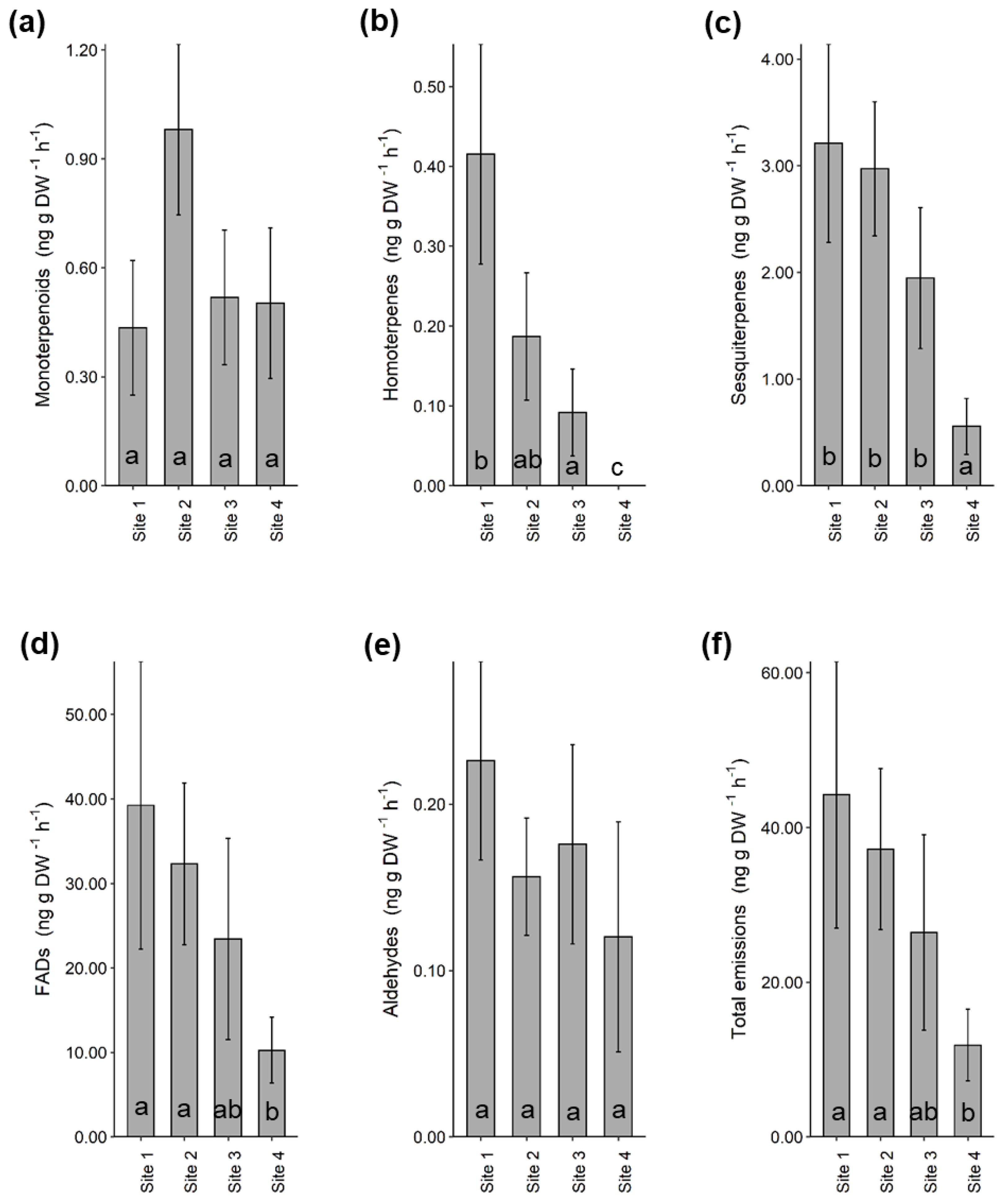
2.1. Volatile Emissions by Heather
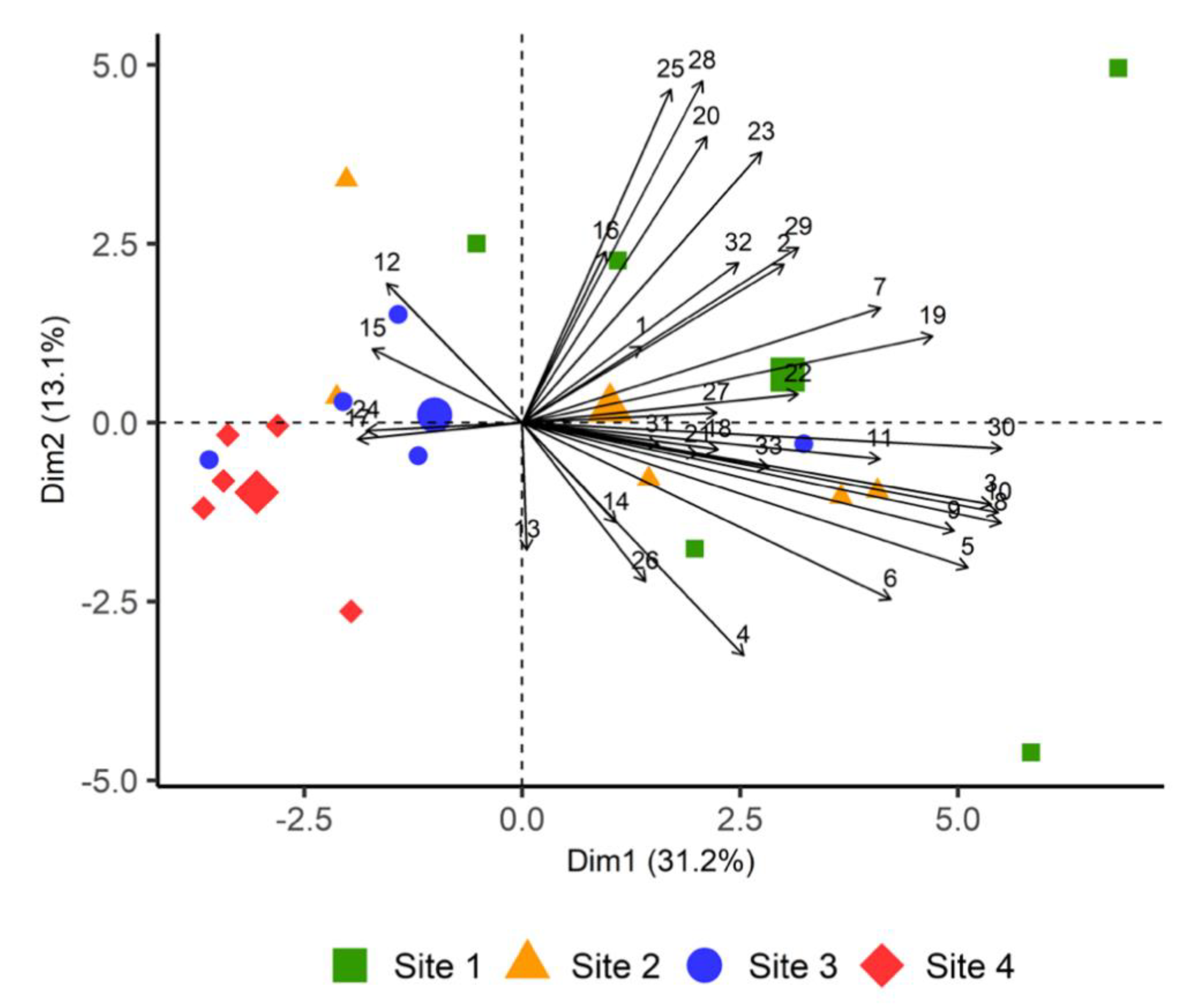
2.2. Principal Component Analysis Based on Volatiles Emissions at Different Sites
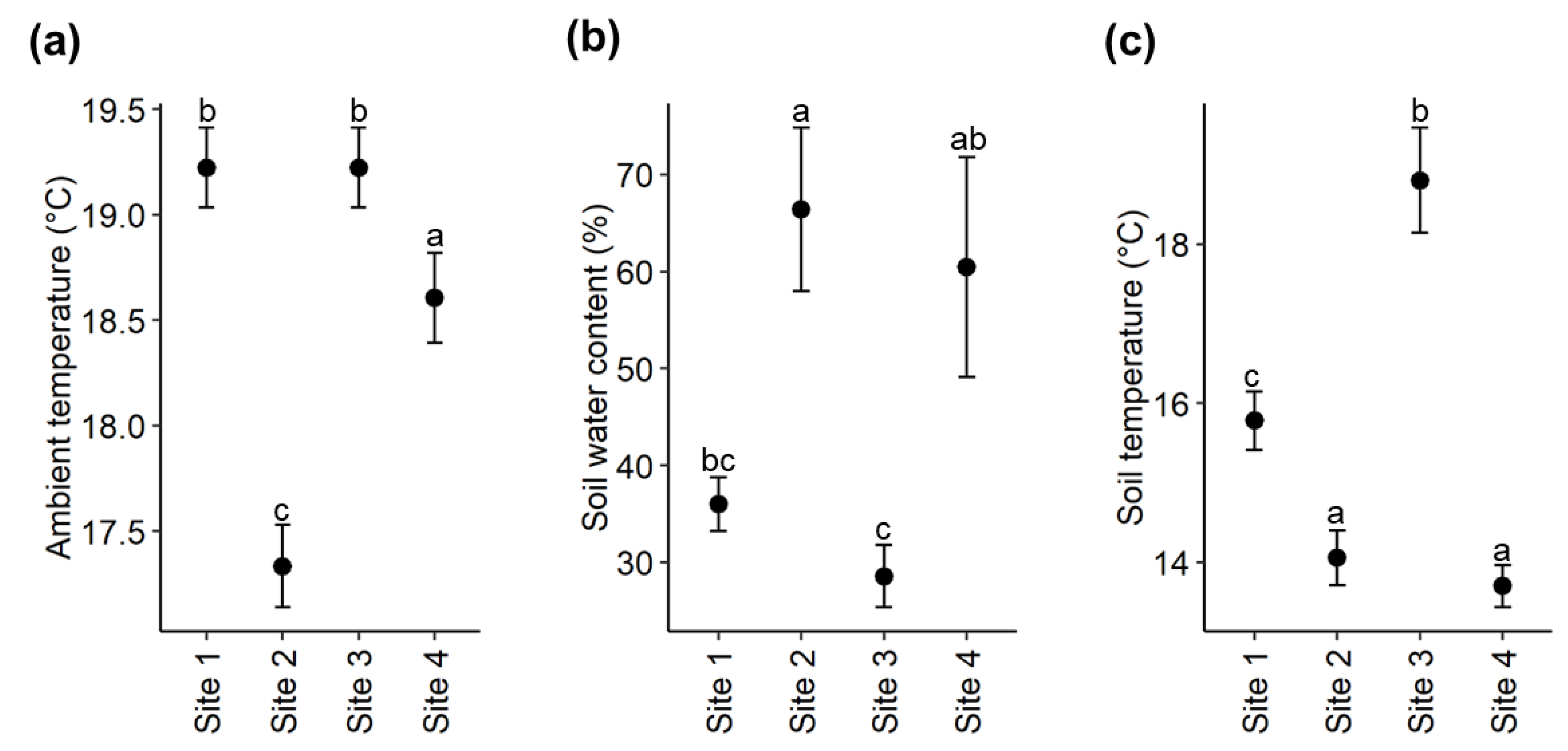
2.3. Soil Nutrients and Environmental Variables
2.4. Arthropod Community Composition
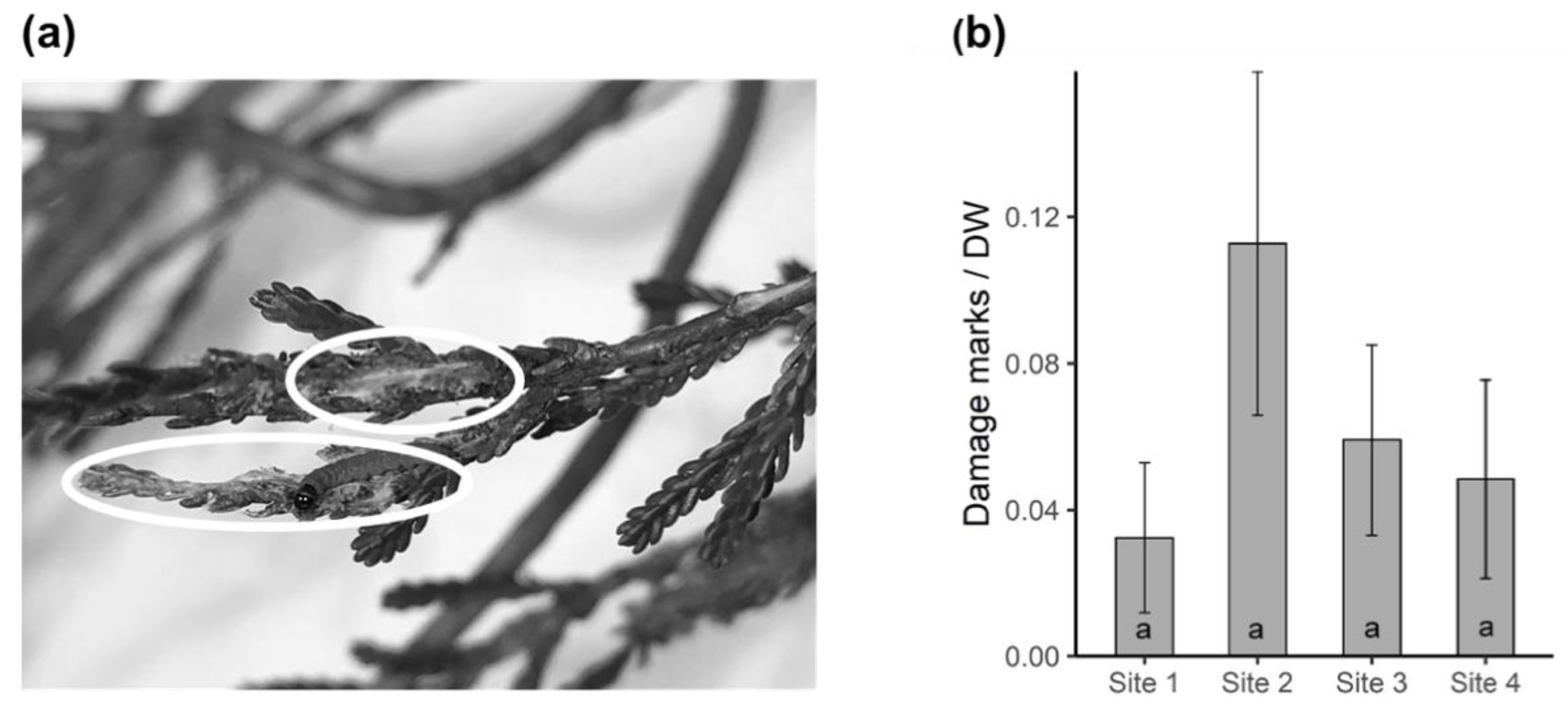
2.5. Herbivore Damage on Heather
2.6. Effect of Biotic and Abiotic Factors on Volatile Emissions
3. Discussion
4. Conclusions
5. Materials and Methods
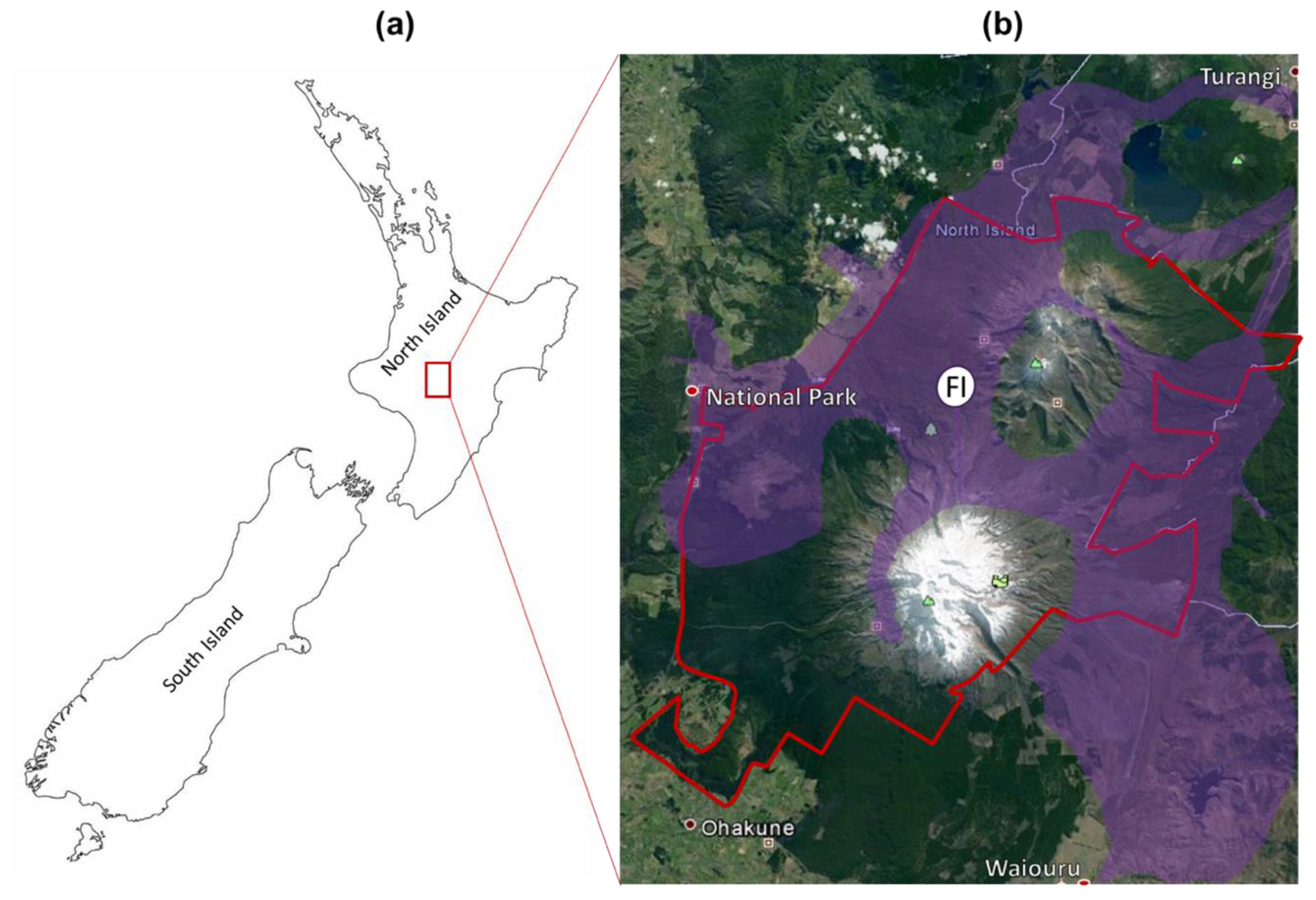
5.1. Study Area
5.2. Sampling of Volatiles
5.3. Soil Sampling
5.4. Ambient and Soil Temperature Measurements
5.5. Arthropods on Heather
5.6. Herbivore Damage on Heather
5.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nentwig, W. Biological Invasions: Why it Matters. In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–6. [Google Scholar]
- Seebens, H.; Essl, F.; Dawson, W.; Fuentes, N.; Moser, D.; Pergl, J.; Pyšek, P.; van Kleunen, M.; Weber, E.; Winter, M.; et al. Global trade will accelerate plant invasions in emerging economies under climate change. Glob. Chang. Biol. 2015, 21, 4128–4140. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.S.; Makra, L.; Albertini, R.; Bonini, M.; Páldy, A.; Rodinkova, V.; Šikoparija, B.; Weryszko-Chmielewska, E.; Bullock, J.M. Modelling the introduction and spread of non-native species: International trade and climate change drive ragweed invasion. Glob. Chang. Biol. 2016, 22, 3067–3079. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Oduor, A.M.O.; Zhang, Z.; Manea, A.; Tooth, I.M.; Leishman, M.R.; Xu, X.; van Kleunen, M. Do invasive alien plants benefit more from global environmental change than native plants? Glob. Chang. Biol. 2017, 23, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Aschehoug, E.T. Invasive Plants Versus Their New and Old Neighbors: A Mechanism for Exotic Invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and Exotic Plant Invasion: From Molecules and Genes to Species Interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef]
- Hierro, J.L.; Callaway, R.M. Allelopathy and exotic plant invasion. Plant Soil 2003, 256, 29–39. [Google Scholar] [CrossRef]
- Orr, S.P.; Rudgers, J.A.; Clay, K. Invasive Plants can Inhibit Native Tree Seedlings: Testing Potential Allelopathic Mechanisms. Plant Ecol. 2005, 181, 153–165. [Google Scholar] [CrossRef]
- Effah, E.; Holopainen, J.K.; McCormick, A.C. Potential roles of volatile organic compounds in plant competition. Perspect. Plant Ecol. Evol. Syst. 2019, 38, 58–63. [Google Scholar] [CrossRef]
- Holopainen, J.K. Multiple functions of inducible plant volatiles. Trends Plant Sci. 2004, 9, 529–533. [Google Scholar] [CrossRef]
- Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G.; Mancuso, S. The network of plants volatile organic compounds. Sci. Rep. 2017, 7, 11050. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.C.; Reinecke, A.; Gershenzon, J.; Unsicker, S.B. Feeding Experience Affects the Behavioral Response of Polyphagous Gypsy Moth Caterpillars to Herbivore-induced Poplar Volatiles. J. Chem. Ecol. 2016, 42, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Barney, J.N.; Sparks, J.P.; Greenberg, J.; Whitlow, T.H.; Guenther, A. Biogenic volatile organic compounds from an invasive species: Impacts on plant–plant interactions. Plant Ecol. 2009, 203, 195–205. [Google Scholar] [CrossRef][Green Version]
- Ens, E.J.; Bremner, J.B.; French, K.; Korth, J. Identification of volatile compounds released by roots of an invasive plant, bitou bush (Chrysanthemoides monilifera spp. rotundata), and their inhibition of native seedling growth. Biol. Invasions 2009, 11, 275–287. [Google Scholar] [CrossRef]
- Wang, R.-L.; Staehelin, C.; Peng, S.-L.; Wang, W.-T.; Xie, X.-M.; Lu, H.-N. Responses of Mikania micrantha, an Invasive Weed to Elevated CO2: Induction of β-Caryophyllene Synthase, Changes in Emission Capability and Allelopathic Potential of β-Caryophyllene. J. Chem. Ecol. 2010, 36, 1076–1082. [Google Scholar] [CrossRef]
- Araniti, F.; Lupini, A.; Sunseri, F.; Abenavoli, M.R. Allelopatic Potential of Dittrichia viscosa (L.) W. Greuter Mediated by VOCs: A Physiological and Metabolomic Approach. PLoS ONE 2017, 12, e0170161. [Google Scholar] [CrossRef]
- Kegge, W.; Pierik, R. Biogenic volatile organic compounds and plant competition. Trends Plant Sci. 2010, 15, 126–132. [Google Scholar] [CrossRef]
- Rogers, G.M. North Island seral tussock grasslands 1. Origins and land-use history. N. Z. J. Bot. 1994, 32, 271–286. [Google Scholar] [CrossRef][Green Version]
- Bagnall, A.G. Heather at Tongariro. A study of a weed introduction. Tussock Grasslands Mt. Lands Inst. Rev. 1982, 41, 17–21. [Google Scholar]
- Chapman, H.M.; Bannister, P. The spread of heather, Calluna vulgaris (L.) Hull, into indigenous plant communities of Tongariro National Park. N. Z. J. Ecol. 1990, 14, 7–16. [Google Scholar]
- Keesing, V.F. Impacts of Invasion on Community Structure: Habitat and Invertebrate Assemblage Responses to Calluna Vulgaris (L.) Hull Invasion, in Tongariro National Park, New Zealand. Ph.D. Thesis, Massey University, Palmerston, New Zealand, 1995. [Google Scholar]
- Rogers, G.M. Control, Demography, and Post-control Response of Heather in the Central North Island. Sci. Conserv. 1995, 9, 20. [Google Scholar]
- Rogers, G.M. Control, demography, and post-control response of heather in the central North Island: Part 2. Sci. Conserv. 1996, 29, 1–35. [Google Scholar]
- Rogers, G.M.; Leathwick, J.R. North Island seral tussock grasslands. 3. The influence of heather (Calluna vulgaris) on rates of change from tussock grassland to shrubland. N. Z. J. Bot. 1996, 34, 473–487. [Google Scholar] [CrossRef]
- Peterson, P.G.; Fowler, S.V.; Barrett, P. Is the poor establishment and performance of heather beetle in Tongariro National Park due to the impact of parasitoids predators or disease. N. Z. Plant Prot. 2004, 57, 89–93. [Google Scholar] [CrossRef]
- Tiiva, P.; Tang, J.; Michelsen, A.; Rinnan, R. Monoterpene emissions in response to long-term night-time warming, elevated CO2 and extended summer drought in a temperate heath ecosystem. Sci. Total Environ. 2017, 580, 1056–1067. [Google Scholar] [CrossRef] [PubMed]
- Moisan, K.; Cordovez, V.; van de Zande, E.M.; Raaijmakers, J.M.; Dicke, M.; Lucas-Barbosa, D. Volatiles of pathogenic and non-pathogenic soil-borne fungi affect plant development and resistance to insects. Oecologia 2019, 190, 589–604. [Google Scholar] [CrossRef]
- Gouinguené, S.P.; Turlings, T.C.J. The Effects of Abiotic Factors on Induced Volatile Emissions in Corn Plants. Plant Physiol. 2002, 129, 1296. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Stewart-Jones, A.; Pulkkinen, J.; Poppy, G.M.; Holopainen, J.K. The influence of different nutrient levels on insect-induced plant volatiles in Bt and conventional oilseed rape plants. Plant Biol. 2008, 10, 97–107. [Google Scholar] [CrossRef]
- Veromann, E.; Toome, M.; Kännaste, A.; Kaasik, R.; Copolovici, L.; Flink, J.; Kovács, G.; Narits, L.; Luik, A.; Niinemets, Ü. Effects of nitrogen fertilization on insect pests, their parasitoids, plant diseases and volatile organic compounds in Brassica napus. Crop Protect. 2013, 43, 79–88. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Wright, D.H.; Harris, N.D. Effect of nitrogen and potassium fertilization on tomato flavor. J. Agric. Food Chem. 1985, 33, 355–358. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Karimi, E.; Ghasemzadeh, A. Primary, Secondary Metabolites, Photosynthetic Capacity and Antioxidant Activity of the Malaysian Herb Kacip Fatimah (Labisia pumila Benth) Exposed to Potassium Fertilization under Greenhouse Conditions. Int. J. Mol. Sci. 2012, 13, 15321–15342. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, L.; Moreira, X.; Zas, R. Costs of constitutive and herbivore-induced chemical defences in pine trees emerge only under low nutrient availability. J. Ecol. 2011, 99, 818–827. [Google Scholar] [CrossRef]
- McCormick, A.C.; Unsicker, S.B.; Gershenzon, J. The specificity of herbivore-induced plant volatiles in attracting herbivore enemies. Trends Plant Sci. 2012, 17, 303–310. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.C.; Irmisch, S.; Reinecke, A.; Boeckler, G.A.; Veit, D.; Reichelt, M.; Hansson, B.s.; Gershenzon, J.; Köllner, T.g.; Unsicker, S.b. Herbivore-induced volatile emission in black poplar: Regulation and role in attracting herbivore enemies. Plant Cell Environ. 2014, 37, 1909–1923. [Google Scholar] [CrossRef] [PubMed]
- Steven, S.; Uefune, M.; Ozawa, R.; Takabayashi, J.; Kainoh, Y. Oviposition Experience of Parasitoid Wasps with Nonhost Larvae Affects their Olfactory and Contact-Behavioral Responses toward Host- and Nonhost-Infested Plants. J. Chem. Ecol. 2019, 45, 402–409. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2018, 220, 692–702. [Google Scholar] [CrossRef]
- Langenheim, J.H. Higher plant terpenoids: A phytocentric overview of their ecological roles. J. Chem. Ecol. 1994, 20, 1223–1280. [Google Scholar] [CrossRef]
- Cheng, A.-X.; Lou, Y.-G.; Mao, Y.-B.; Lu, S.; Wang, L.-J.; Chen, X.-Y. Plant Terpenoids: Biosynthesis and Ecological Functions. J. Integr. Plant Biol. 2007, 49, 179–186. [Google Scholar] [CrossRef]
- Loreto, F.; Schnitzler, J.-P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.V.P.; Holopainen, J.K.; Guerrero, A. Olfactory Responses of Plutella xylostella Natural Enemies to Host Pheromone, Larval Frass, and Green Leaf Cabbage Volatiles. J. Chem. Ecol. 2002, 28, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Pierre, P.S.; Jansen, J.J.; Hordijk, C.A.; van Dam, N.M.; Cortesero, A.-M.; Dugravot, S. Differences in Volatile Profiles of Turnip Plants Subjected to Single and Dual Herbivory Above- and Belowground. J. Chem. Ecol. 2011, 37, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Litt, A.r.; Cord, E.e.; Fulbright, T.e.; Schuster, G.l. Effects of Invasive Plants on Arthropods. Conserv. Biol. 2014, 28, 1532–1549. [Google Scholar] [CrossRef] [PubMed]
- Harman, H.M.; Syrett, P.; Hill, R.L.; Jessep, C.T. Arthropod introductions for biological control of weeds in New Zealand, 1929–1995. N. Z. Entomol. 1996, 19, 71–80. [Google Scholar] [CrossRef]
- Copolovici, L.; Kännaste, A.; Pazouki, L.; Niinemets, Ü. Emissions of green leaf volatiles and terpenoids from Solanum lycopersicum are quantitatively related to the severity of cold and heat shock treatments. J. Plant Physiol. 2012, 169, 664–672. [Google Scholar] [CrossRef]
- Rasulov, B.; Bichele, I.; Hüve, K.; Vislap, V.; Niinemets, Ü. Acclimation of isoprene emission and photosynthesis to growth temperature in hybrid aspen: Resolving structural and physiological controls. Plant Cell Environ. 2015, 38, 751–766. [Google Scholar] [CrossRef]
- Li, T.; Holst, T.; Michelsen, A.; Rinnan, R. Amplification of plant volatile defence against insect herbivory in a warming Arctic tundra. Nat. Plants 2019, 5, 568–574. [Google Scholar] [CrossRef]
- Copolovici, L.; Niinemets, Ü. Environmental Impacts on Plant Volatile Emission. In Deciphering Chemical Language of Plant Communication; Blande, J.D., Glinwood, R., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 35–59. [Google Scholar]
- McCormick, A.C. Can plant–natural enemy communication withstand disruption by biotic and abiotic factors? Ecol. Evol. 2016, 6, 8569–8582. [Google Scholar] [CrossRef]
- Possell, M.; Loreto, F. The Role of Volatile Organic Compounds in Plant Resistance to Abiotic Stresses: Responses and Mechanisms. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Niinemets, Ü., Monson, R.K., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 209–235. [Google Scholar]
- Macdonald, J.A.; Fowler, D.; Hargreaves, K.J.; Skiba, U.; Leith, I.D.; Murray, M.B. Methane emission rates from a northern wetland; response to temperature, water table and transport. Atmos. Environ. 1998, 32, 3219–3227. [Google Scholar] [CrossRef]
- Dinnin, M.; Van de Noort, R. Wetland habitats, their resource potential and exploitation. A case study from the Humber wetlands. In Bog Bodies, Sacred Sites and Wetland Archaeology: Proceedings of a Conference Held by WARP and the National Museum of Denmark, in Conjuction with Silkeborg Museum, Jutland; Coles, B., Coles, J., Shou-Jørgenson, M., Eds.; WARP: Exeter, UK, 1999; pp. 69–78. [Google Scholar]
- Pierik, R.; Visser, E.J.W.; De Kroon, H.; Voesenek, L.A.C.J. Ethylene is required in tobacco to successfully compete with proximate neighbours. Plant Cell Environ. 2003, 26, 1229–1234. [Google Scholar] [CrossRef]
- Kigathi, R.N.; Weisser, W.W.; Veit, D.; Gershenzon, J.; Unsicker, S.B. Plants Suppress Their Emission of Volatiles When Growing with Conspecifics. J. Chem. Ecol. 2013, 39, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Kigathi, R.N.; Weisser, W.W.; Reichelt, M.; Gershenzon, J.; Unsicker, S.B. Plant volatile emission depends on the species composition of the neighboring plant community. BMC Plant Biol. 2019, 19, 58. [Google Scholar] [CrossRef] [PubMed]
- Leathwick, J.R.; Mitchell, N.D. Forest pattern, climate and vulcanism in central North Island, New Zealand. J. Veg. Sci. 1992, 3, 603–616. [Google Scholar] [CrossRef]
- Tholl, D.; Boland, W.; Hansel, A.; Loreto, F.; Röse, U.S.R.; Schnitzler, J.-P. Practical approaches to plant volatile analysis. Plant J. 2006, 45, 540–560. [Google Scholar] [CrossRef] [PubMed]
- Runyon, J.B.; Birdsall, J.L. Costs of induced defenses for the invasive plant houndstongue (Cynoglossum officinale L.) and the potential importance for weed biocontrol. Arthropod-Plant Interact. 2016, 10, 383–391. [Google Scholar] [CrossRef]
- Martini, X.; Coy, M.; Kuhns, E.; Stelinski, L.L. Temporal Decline in Pathogen-Mediated Release of Methyl Salicylate Associated With Decreasing Vector Preference for Infected Over Uninfected Plants. 2018; Volume 6. [Google Scholar]
- Chapman, H.M.; Bannister, P. Flowering, shoot extension, and reproductive performance of heather, Calluna vulgaris (L.) Hull, in Tongariro National Park, New Zealand. N. Z. J. Bot. 1995, 33, 111–119. [Google Scholar] [CrossRef]
- Haney, R.L.; Haney, E.B. Simple and Rapid Laboratory Method for Rewetting Dry Soil for Incubations. Commun. Soil Sci. Plant Anal. 2010, 41, 1493–1501. [Google Scholar] [CrossRef]
- Wang, F.; Dang, C.; Chang, X.; Tian, J.; Lu, Z.; Chen, Y.; Ye, G. Variation among conventional cultivars could be used as a criterion for environmental safety assessment of Bt rice on nontarget arthropods. Sci. Rep. 2017, 7, 41918. [Google Scholar] [CrossRef]
- Shearer, P.W.; Amarasekare, K.G.; Castagnoli, S.P.; Beers, E.H.; Jones, V.P.; Mills, N.J. Large-plot field studies to assess impacts of newer insecticides on non-target arthropods in Western U.S. orchards. Biol. Control 2016, 102, 26–34. [Google Scholar] [CrossRef][Green Version]
- Team, R. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Husson, F.; Josse, J.; Le, S.; Mazet, J. Multivariate Exploratory Data Analysis and Data Mining. Package ‘FactoMineR’. 2018. Available online: https://mran.microsoft.com/snapshot/2018-03-30/web/packages/FactoMineR/index.html (accessed on 18 February 2020).
- Csata, E.; Timuş, N.; Witek, M.; Casacci, L.P.; Lucas, C.; Bagnères, A.-G.; Sztencel-Jabłonka, A.; Barbero, F.; Bonelli, S.; Rákosy, L.; et al. Lock-picks: Fungal infection facilitates the intrusion of strangers into ant colonies. Sci. Rep. 2017, 7, 46323. [Google Scholar] [CrossRef] [PubMed]
- Dowdy, N.J.; Conner, W.E. Nonchalant Flight in Tiger Moths (Erebidae: Arctiinae) Is Correlated With Unpalatability. Front. Ecol. Evol. 2019, 7, 480. [Google Scholar] [CrossRef]
- Larsson, B.; Dragioti, E.; Grimby-Ekman, A.; Gerdle, B.; Björk, J. Predictors of Chronic Pain Intensity, Spread and Sensitivity in the General Population: A Two-year Follow-up Study from the Swepain Cohort. J. Rehabil. Med. 2019, 51, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Chauhan, D.K.; Prasad, S.M.; Dubey, N.K. Role of Macronutrients in Plant Growth and Acclimation: Recent Advances and Future Prospective. In Improvement of Crops in the Era of Climatic Changes: Volume 2; Ahmad, P., Wani, M.R., Azooz, M.M., Phan Tran, L.-S., Eds.; Springer: New York, NY, USA, 2014; pp. 197–216. [Google Scholar]
- Kuhn, M.; Johnson, K. Applied Predictive Modeling; Springer: New York, NY, USA, 2013; Volume 26. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Site 1 | Site 2 | Site 3 | Site 4 | Reference (Medium Range) |
|---|---|---|---|---|---|
| Total nitrogen (%) | 0.19 | 0.30 | 0.17 | 0.26 | 0.30–0.60 |
| Total carbon (%) | 3.70 | 6.00 | 3.10 | 2.80 | NA |
| Olsen phosphorus (me/100 g) | 5.00 | 3.00 | 4.00 | 3.00 | 20–30 |
| Sodium (me/100 g) | 0.06 | 0.06 | 0.10 | <0.05 | 0.20–0.50 |
| Magnesium (me/100 g) | 0.48 | 0.31 | 0.34 | 0.16 | 1.00–1.60 |
| Calcium (me/100 g) | 2.70 | 1.60 | 1.50 | 0.70 | 4.0–10.0 |
| Potassium (me/100 g) | 0.18 | 0.24 | 0.22 | 0.13 | 0.40–0.60 |
| Organic matter (%) | 6.30 | 10.4 | 5.30 | 4.80 | 7.0–17.0 |
| pH | 5.70 | 5.70 | 5.50 | 5.80 | 5.8–6.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Effah, E.; Barrett, D.P.; Peterson, P.G.; Godfrey, A.J.R.; Potter, M.A.; Holopainen, J.K.; Clavijo McCormick, A. Natural Variation in Volatile Emissions of the Invasive Weed Calluna vulgaris in New Zealand. Plants 2020, 9, 283. https://doi.org/10.3390/plants9020283
Effah E, Barrett DP, Peterson PG, Godfrey AJR, Potter MA, Holopainen JK, Clavijo McCormick A. Natural Variation in Volatile Emissions of the Invasive Weed Calluna vulgaris in New Zealand. Plants. 2020; 9(2):283. https://doi.org/10.3390/plants9020283
Chicago/Turabian StyleEffah, Evans, D. Paul Barrett, Paul G. Peterson, A. Jonathan R. Godfrey, Murray A. Potter, Jarmo K. Holopainen, and Andrea Clavijo McCormick. 2020. "Natural Variation in Volatile Emissions of the Invasive Weed Calluna vulgaris in New Zealand" Plants 9, no. 2: 283. https://doi.org/10.3390/plants9020283
APA StyleEffah, E., Barrett, D. P., Peterson, P. G., Godfrey, A. J. R., Potter, M. A., Holopainen, J. K., & Clavijo McCormick, A. (2020). Natural Variation in Volatile Emissions of the Invasive Weed Calluna vulgaris in New Zealand. Plants, 9(2), 283. https://doi.org/10.3390/plants9020283