Molecular Basis of Root Nodule Symbiosis between Bradyrhizobium and ‘Crack-Entry’ Legume Groundnut (Arachis hypogaea L.)

 ,
, 
 , ,
, , 


 and
and 
Abstract
1. Introduction
2. Diversity among Bacterial Strains Associated with Groundnut RNS
3. Effect of Abiotic Stress on Groundnut Root Nodule Symbiosis
3.1. Temperature
3.2. Osmotic and Saline Stress
3.3. Soil pH
4. Processes Involved during Groundnut Root-Nodule Symbiosis
4.1. Pre-Infection
4.2. Role of Rhizobial Exopolysaccharides in Root-Nodule Symbiosis
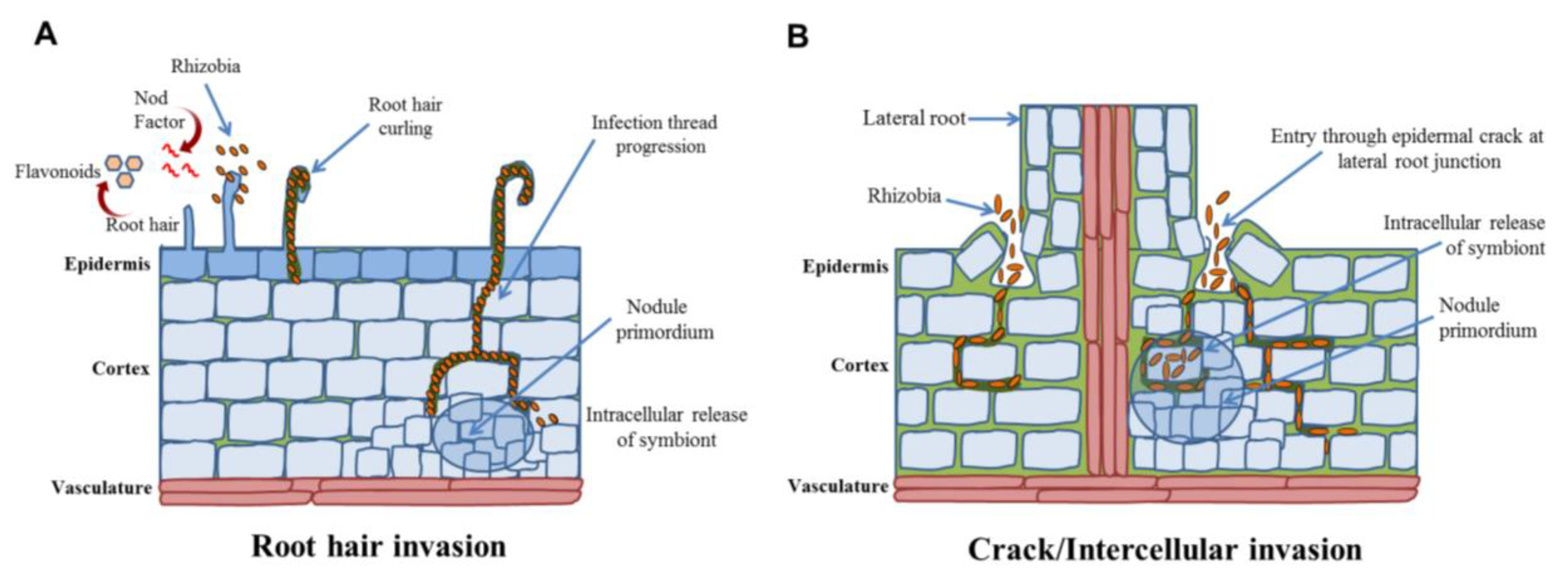
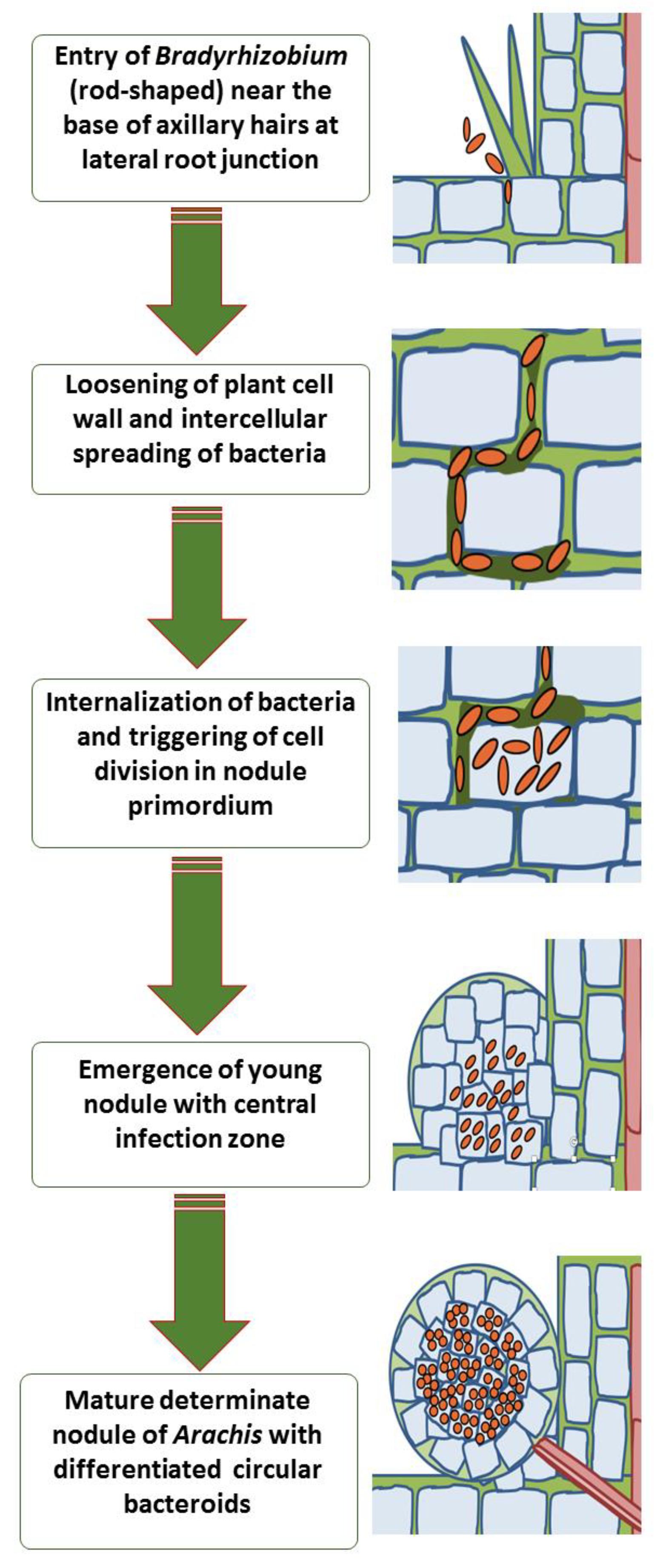
4.3. Routes of Bacterial Invasion
4.4. Structural Features of Nodule
5. Molecular Basis of Root Nodule Symbiosis Initiation in Groundnut
6. Components of Symbiotic Signalling Pathway in Groundnut
6.1. AhNFR1 and AhNFR5
6.2. AhSYMRK
6.3. AhCCaMK
6.4. AhCYCLOPS
6.5. AhHK1
7. Signalling and Phytohormone Pathways Involved during Nodulation in Groundnut
7.1. Auxin
7.2. Cytokinin
7.3. Gibberellins
7.4. Ethylene
7.5. Abscisic Acid
8. Utilization of Existing Knowledge to Enhance N2 Use Efficiency by Groundnut and Cereals
8.1. Application of Transgenic Approaches and Genome Editing
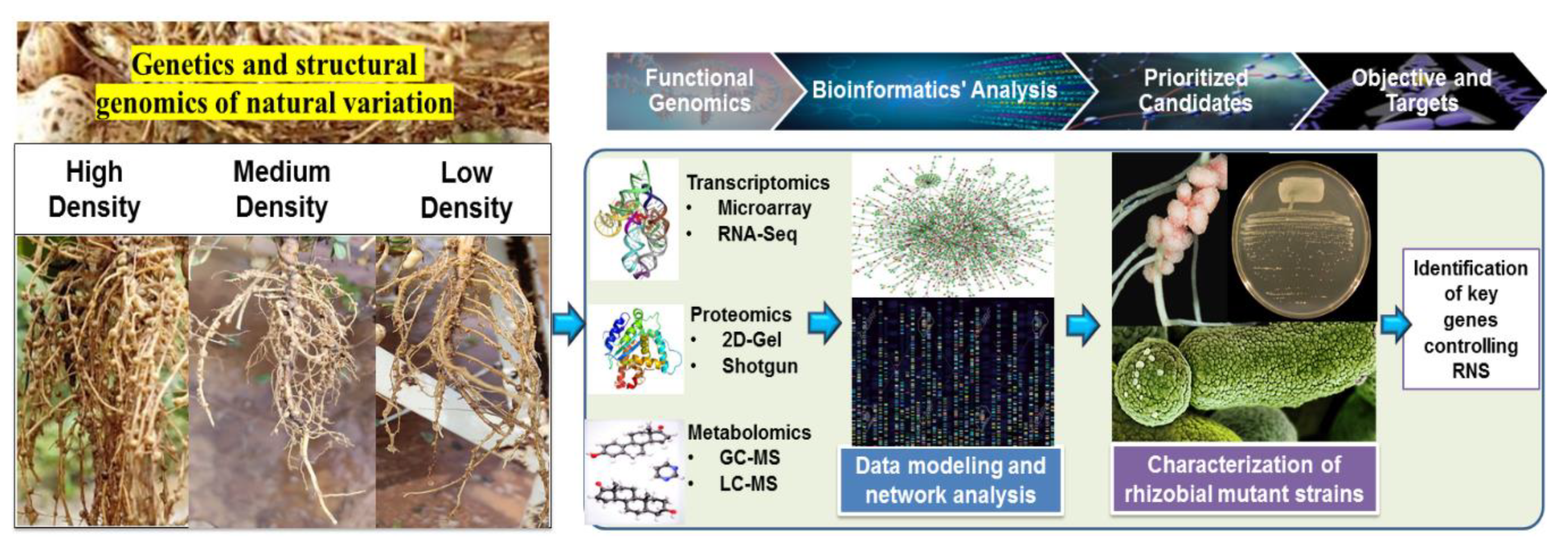
8.2. Context of Advanced Bioinformatics in Groundnut Research
8.3. Strategies to Extend Nitrogen Fixation to Non-Legumes
9. Current Menace of Inorganic Fertilizers—Key Challenges
10. Enhancement of Biological Nitrogen Content in Soil by Intercropping with Groundnut
11. Harnessing the Genetic Variation through Conventional and Molecular Breeding for Improving Nitrogen Fixation
12. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Azani, N.; Babineau, M.; Bailey, C.D.; Banks, H.; Barbosa, A.; Pinto, R.B.; Boatwright, J.; Borges, L.; Brown, G.; Bruneau, A.; et al. A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny – The Legume Phylogeny Working Group (LPWG). TAXON 2017, 66, 44–77. [Google Scholar] [CrossRef]
- Lewis, G.; Schrire, B.; Mackinder, B.; Rico, L.; Clark, R. A linear sequence of legume genera set in a phylogenetic context — A tool for collections management and taxon sampling. South Afr. J. Bot. 2013, 89, 76–84. [Google Scholar] [CrossRef]
- Lavin, M.; Herendeen, P.S.; Wojciechowski, M.F. Evolutionary Rates Analysis of Leguminosae Implicates a Rapid Diversification of Lineages during the Tertiary. Syst. Biol. 2005, 54, 575–594. [Google Scholar] [CrossRef] [PubMed]
- Sprent, J.I. Evolving ideas of legume evolution and diversity: A taxonomic perspective on the occurrence of nodulation. New Phytol. 2007, 174, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Young, N.D.; Bharti, A.K. Genome-Enabled Insights into Legume Biology. Annu. Rev. Plant Biol. 2012, 63, 283–305. [Google Scholar] [CrossRef]
- Jensen, E.S.; Hauggaard-Nielsen, H. How can increased use of biological nitrogen fixation in agriculture benefit the environment? Plant Soil 2003, 252, 177–186. [Google Scholar] [CrossRef]
- Garg, N. Geetanjali Symbiotic nitrogen fixation in legume nodules: Process and signaling. A review. Agron. Sustain. Dev. 2007, 27, 59–68. [Google Scholar] [CrossRef]
- Kistner, C.; Parniske, M. Evolution of signal transduction in intracellular symbiosis. Trends Plant Sci. 2002, 7, 511–518. [Google Scholar] [CrossRef]
- Gage, D.J. Infection and Invasion of Roots by Symbiotic, Nitrogen-Fixing Rhizobia during Nodulation of Temperate Legumes. Microbiol. Mol. Biol. Rev. 2004, 68, 280–300. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D.; Downie, J.A. Calcium, kinases and nodulation signalling in legumes. Nat. Rev. Mol. Cell Biol. 2004, 5, 566–576. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Downie, J.A. Nuclear calcium changes at the core of symbiosis signalling. Curr. Opin. Plant Biol. 2006, 9, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Sprent, J.I.; James, E.K. Legume Evolution: Where Do Nodules and Mycorrhizas Fit In? Plant Physiol. 2007, 144, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Fabre, S.; Gully, D.; Poitout, A.; Patrel, D.; Arrighi, J.-F.; Giraud, E.; Czernic, P.; Cartieaux, F. Nod Factor-Independent Nodulation in Aeschynomene evenia Required the Common Plant-Microbe Symbiotic Toolkit1. Plant Physiol. 2015, 169, 2654–2664. [Google Scholar] [PubMed]
- Wang, J. Wang’s Lab UF Home Page. Available online: https://www.wanglabuf.com/projects (accessed on 10 November 2019).
- Karmakar, K.; Kundu, A.; Rizvi, A.Z.; Dubois, E.; Severac, D.; Czernic, P.; Cartieaux, F.; Dasgupta, M. Transcriptomic Analysis With the Progress of Symbiosis in ‘Crack-Entry’ Legume Arachis hypogaea Highlights Its Contrast With ‘Infection Thread’ Adapted Legumes. Mol. Plant-Microbe Interact. 2019, 32, 271–285. [Google Scholar] [CrossRef]
- Peng, Z.; Liu, F.; Wang, L.; Zhou, H.; Paudel, D.; Tan, L.; Maku, J.; Gallo, M.; Wang, J. Transcriptome profiles reveal gene regulation of peanut (Arachis hypogaea L.) nodulation. Sci. Rep. 2017, 7, 40066. [Google Scholar] [CrossRef]
- Weir, B.S. The Current Taxonomy of Rhizobia. NZ Rhizobia Website. Available online: https://www.rhizobia.co.nz/taxonomy/rhizobia (accessed on 10 November 2019).
- Delamuta, J.R.M.; Ribeiro, R.A.; Menna, P.; Bangel, E.V.; Hungria, M. Multilocus sequence analysis (MLSA) of Bradyrhizobium strains: Revealing high diversity of tropical diazotrophic symbiotic bacteria. Braz. J. Microbiol. 2012, 43, 698–710. [Google Scholar] [CrossRef][Green Version]
- VanInsberghe, D.; Maas, K.R.; Cardenas, E.; Strachan, C.R.; Hallam, S.J.; Mohn, W.W. Non-symbiotic Bradyrhizobium ecotypes dominate North American forest soils. ISME J. 2015, 9, 2435–2441. [Google Scholar] [CrossRef]
- Nkot, L.N.; Krasova-Wade, T.; Etoa, F.; Sylla, S.; Nwaga, D. Genetic diversity of rhizobia nodulating Arachis hypogaea L. in diverse land use systems of humid forest zone in Cameroon. Appl. Soil Ecol. 2008, 40, 411–416. [Google Scholar] [CrossRef]
- Chen, J.Y.; Gu, J.; Wang, E.T.; Ma, X.X.; Kang, S.T.; Huang, L.Z.; Cao, X.P.; Li, L.B.; Wu, Y.L. Wild peanut Arachis duranensis are nodulated by diverse and novel Bradyrhizobium species in acid soils. Syst. Appl. Microbiol. 2014, 37, 525–532. [Google Scholar] [CrossRef]
- Mokgehle, S.N.; Dakora, F.D.; Mathews, C. Variation in N2 fixation and N contribution by 25 groundnut (Arachis hypogaea L.) varieties grown in different agro-ecologies, measured using 15N natural abundance. Agric. Ecosyst. Environ. 2014, 195, 161–172. [Google Scholar] [CrossRef]
- Ibáñez, F.; Wall, L.; Fabra, A. Starting points in plant-bacteria nitrogen-fixing symbioses: Intercellular invasion of the roots. J. Exp. Bot. 2016, 68, 1905–1918. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, V.; Ibañez, F.; Tonelli, M.L.; Valetti, L.; Anzuay, M.S.; Fabra, A. Phenotypic and phylogenetic characterization of native peanut Bradyrhizobium isolates obtained from Córdoba, Argentina. Syst. Appl. Microbiol. 2011, 34, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Grönemeyer, J.L.; Hurek, T.; Bünger, W.; Reinhold-Hurek, B. Bradyrhizobium vignae sp. nov., a nitrogen-fixing symbiont isolated from effective nodules of Vigna and Arachis. Int. J. Syst. Evol. Microbiol. 2016, 66, 62–69. [Google Scholar]
- Zazou, A.Z.; Fonceka, D.; Fall, S.; Fabra, A.; Ibañez, F.; Pignoly, S.; Diouf, A.; Touré, O.; Faye, M.N.; Hocher, V.; et al. Genetic diversity and symbiotic efficiency of rhizobial strains isolated from nodules of peanut (Arachis hypogaea L.) in Senegal. Agric. Ecosyst. Environ. 2018, 265, 384–391. [Google Scholar] [CrossRef]
- Yang, J.K.; Zhou, J.C. Diversity, phylogeny and host specificity of soybean and peanut bradyrhizobia. Biol. Fertil. Soils 2008, 44, 843–851. [Google Scholar] [CrossRef]
- Krasova-Wade, T.; Ndoye, I.; Braconnier, S.; Sarr, B.; De Lajudie, P.; Neyra, M. Diversity of indigeneous bradyrhizobia associated with three cowpea cultivars (Vignaunguiculata (L.) Walp.) grown under limited and favorable water conditions in Senegal (West Africa). Afr. J. Biotech. 2003, 2, 13–22. [Google Scholar]
- Taurian, T.; Ibañez, F.; Fabra, A.; Aguilar, O.M. Genetic Diversity of Rhizobia Nodulating Arachis hypogaea L. in Central Argentinean Soils. Plant Soil 2006, 282, 41–52. [Google Scholar] [CrossRef]
- El-Akhal, M.R.; Rincón, A.; Arenal, F.; Lucas, M.M.; El Mourabit, N.; Barrijal, S.; Pueyo, J.J. Genetic diversity and symbiotic efficiency of rhizobial isolates obtained from nodules of Arachis hypogaea in northwestern Morocco. Soil Biol. Biochem. 2008, 40, 2911–2914. [Google Scholar] [CrossRef]
- Velagaleti, R.R.; Marsh, S. Influence of host cultivars and Bradyrhizobium strains on the growth and symbiotic performance of soybean under salt stress. Plant Soil 1989, 119, 133–138. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-Legume Symbiosis and Nitrogen Fixation under Severe Conditions and in an Arid Climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef]
- Stalker, H. Peanut (Arachis hypogaea L.). Field Crop. Res. 1997, 53, 205–217. [Google Scholar] [CrossRef]
- Kishinevsky, B.D.; Sen, D.; Weaver, R.W. Effect of high root temperature on Bradyrhizobium-peanut symbiosis. Plant Soil 1992, 143, 275–282. [Google Scholar] [CrossRef]
- Dardanelli, M.S.; Woelke, M.R.; Gonzalez, P.S.; Bueno, L.M.; Ghittoni, N.E. The effects of nonionic hyper osmolarity and of high temperature on cell-associated low molecular weight saccharides from two peanut rhizobia strains. Symbiosis 1997, 23, 73–84. [Google Scholar]
- Roughley, R.J.; Dart, P.J. Root temperature and root-hair infection of Trifolium subterraneum L. cv. Cranmore. Plant Soil 1970, 32, 518–520. [Google Scholar] [CrossRef]
- Michiels, J.; Verreth, C.; Vanderleyden, J. Effects of Temperature Stress on Bean-Nodulating Rhizobium Strains. Appl. Environ. Microbiol. 1994, 60, 1206–1212. [Google Scholar] [CrossRef]
- Medeot, D.B.; Bueno, M.A.; Dardanelli, M.S.; De Lema, M.G. Adaptational Changes in Lipids of Bradyrhizobium SEMIA 6144 Nodulating Peanut as a Response to Growth Temperature and Salinity. Curr. Microbiol. 2007, 54, 31–35. [Google Scholar] [CrossRef]
- Dardanelli, M.S.; González, P.S.; Medeot, D.B.; Paulucci, N.S.; Bueno, M.Á.; Garcia, M.B. Effects of peanut rhizobia on the growth and symbiotic performance of Arachis hypogaea under abiotic stress. Symbiosis 2009, 47, 175–180. [Google Scholar] [CrossRef]
- Zahran, H.H.; Sprent, J.I. Effects of sodium chloride and polyethylene glycol on root-hair infection and nodulation of Vicia faba L. plants by Rhizobium leguminosarum. Planta 1986, 167, 303–309. [Google Scholar] [CrossRef]
- Hungria, M.; Vargas, M.A. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crop. Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Angelini, J.; Taurian, T.; Morgante, C.; Ibañez, F.; Castro, S.; Fabra, A. Peanut nodulation kinetics in response to low pH. Plant Physiol. Biochem. 2005, 43, 754–759. [Google Scholar] [CrossRef]
- Angelini, J.; Castro, S.; Fabra, A. Alterations in root colonization and nodC gene induction in the peanut–rhizobia interaction under acidic conditions. Plant Physiol. Biochem. 2003, 41, 289–294. [Google Scholar] [CrossRef]
- Wisniewski-Dyé, F.; Downie, J.A. Quorum-sensing in Rhizobium. Antonie van Leeuwenhoek 2002, 81, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Matthysse, A.G.; Kijne, J.W. Attachment of Rhizobiaceae to Plant Cells. In The Rhizobiaceae; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1998; pp. 235–249. [Google Scholar]
- Van Rhijn, P.; Fujishige, N.A.; Lim, P.O.; Hirsch, A.M. Sugar-Binding Activity of Pea Lectin Enhances Heterologous Infection of Transgenic Alfalfa Plants by Rhizobium leguminosarum biovar viciae1. Plant Physiol. 2001, 126, 133–144. [Google Scholar] [CrossRef]
- Dardanelli, M.; Angelini, J.; Fabra, A. A calcium-dependent bacterial surface protein is involved in the attachment of rhizobia to peanut roots. Can. J. Microbiol. 2003, 49, 399–405. [Google Scholar] [CrossRef]
- Pellock, B.J.; Cheng, H.-P.; Walker, G.C. Alfalfa Root Nodule Invasion Efficiency Is Dependent on Sinorhizobium meliloti Polysaccharides. J. Bacteriol. 2000, 182, 4310–4318. [Google Scholar] [CrossRef]
- Mazur, A.; Król, J.E.; Wielbo, J.; Urbanik-Sypniewska, T.; Skorupska, A. Rhizobium leguminosarum bv. trifolii PssP protein is required for exopolysaccharide biosynthesis and polymerization. Mol. Plant-Microbe. Interact. 2002, 15, 388–397. [Google Scholar] [CrossRef]
- Stacey, G. A Lipopolysaccharide Mutant of Bradyrhizobium japonicum that Uncouples Plant from Bacterial Differentiation. Mol. Plant-Microbe Interact. 1991, 4, 332. [Google Scholar] [CrossRef]
- Leigh, J.; Coplin, D. Exopolysaccharides in plant bacteria interactions. Ann. Rev. Microbiol. 1992, 46, 307–346. [Google Scholar] [CrossRef]
- Reuber, T.L.; Reed, J.; Glazebrook, J.; Glucksmann, M.A.; Ahmann, D.; Marra, A.; Walker, G.C. Rhizobium meliloti exopolysaccharides: Genetic analyses and symbiotic importance. Biochem. Soc. Trans. 1991, 19, 636–644. [Google Scholar] [CrossRef][Green Version]
- Van Workum, W.A.T.; Van Slageren, S.; Van Brussel, A.A.N.; Kijne, J.W. Role of Exopolysaccharides of Rhizobium leguminosarum bv. viciae as Host Plant-Specific Molecules Required for Infection Thread Formation During Nodulation of Vicia sativa. Mol. Plant-Microbe Interact. 1998, 11, 1233–1241. [Google Scholar] [CrossRef]
- Morgante, C.; Angelini, J.; Castro, S.; Fabra, A. Role of rhizobial exopolysaccharides in crack entry/intercellular infection of peanut. Soil Biol. Biochem. 2005, 37, 1436–1444. [Google Scholar] [CrossRef]
- Morgante, C.; Castro, S.; Fabra, A. Role of rhizobial EPS in the evasion of peanut defense response during the crack-entry infection process. Soil Biol. Biochem. 2007, 39, 1222–1225. [Google Scholar] [CrossRef]
- Brewin, N.J. Plant Cell Wall Remodelling in the Rhizobium–Legume Symbiosis. Crit. Rev. Plant Sci. 2004, 23, 293–316. [Google Scholar] [CrossRef]
- Kijne, J.W.; Bakhuizen, R.; Van Brussel, A.A.N.; Cremers, H.C.J.C.; Diaz, C.L.; De Pater, B.S.; Smit, G.; Spaink, H.P.; Swart, S.; Wijffelman, C.A.; et al. The Rhizobium trap: Root hair curling in root–nodule symbiosis. Perspect. Plant Cell Recognit. 1992, 48, 267–284. [Google Scholar]
- Jones, K.M.; Kobayashi, H.; Davies, B.W.; Taga, M.E.; Walker, G.C. How rhizobial symbionts invade plants: The Sinorhizobium-Medicago model. Nat. Rev. Genet. 2007, 5, 619–633. [Google Scholar] [CrossRef]
- Pueppke, S.G.; Broughton, W.J. Rhizobium sp. strain NGR234 and R. fredii USDA257 share exceptionally broad, nested host ranges. Mol. Plant-Microbe Interact. 1999, 12, 293–318. [Google Scholar] [CrossRef]
- Doyle, J.J. Phylogenetic perspectives on nodulation: Evolving views of plants and symbiotic bacteria. Trends Plant Sci. 1998, 3, 473–478. [Google Scholar] [CrossRef]
- Naisbitt, T.; James, E.K.; Sprent, J.I. The evolutionary significance of the legume genus Chamaecrista, as determined by nodule structure. New Phytol. 1992, 122, 487–492. [Google Scholar] [CrossRef]
- De Faria, S.M.; Hay, G.T.; Sprent, J.I. Entry of Rhizobia into Roots of Mimosa scabrella Bentham Occurs between Epidermal Cells. Microbiology 1988, 134, 2291–2296. [Google Scholar] [CrossRef]
- Dreyfus, B.L. Nitrogen-fixing nodules indeced by Rhizobium on the stem of the tropical legume Sesbania rostrate. FEMS Microbiol. Lett. 1981, 10, 13–23. [Google Scholar] [CrossRef]
- Subba-Rao, N.S.; Mateos, P.F.; Baker, D.; Pankratz, H.S.; Palma, J.; Dazzo, F.B.; Sprent, J.I. The unique root-nodule symbiosis between Rhizobium and the aquatic legume, Neptunia natans (L. f.) Druce. Planta 1995, 196, 311–320. [Google Scholar] [CrossRef]
- Chandler, M.R. Some Observations on Infection of Arachis hypogaea L. by Rhizobium. J. Exp. Bot. 1978, 29, 749–755. [Google Scholar] [CrossRef]
- Vega-Hernández, M.C.; Pérez-Galdona, R.; Dazzo, F.B.; Jarabo-Lorenzo, A.; Alfayate, M.C.; León-Barrios, M. Novel infection process in the indeterminate root nodule symbiosis between Chamaecytisus proliferus (tagasaste) and Bradyrhizobium sp. New Phytol. 2001, 150, 707–721. [Google Scholar] [CrossRef]
- Boogerd, F.C. Nodulation of groundnut by Bradyrhizobium: A simple infection process by crack entry. FEMS Microbiol. Rev. 1997, 21, 5–27. [Google Scholar] [CrossRef]
- Ndoye, I.; De Billy, F.; Vasse, J.; Dreyfus, B.; Truchet, G. Root nodulation of Sesbania rostrata. J. Bacteriol. 1994, 176, 1060–1068. [Google Scholar] [CrossRef][Green Version]
- Chandler, M.R.; Date, R.A.; Roughley, R.J. Infection and Root-Nodule Development in Stylosanthes Species by Rhizobium. J. Exp. Bot. 1982, 33, 47–57. [Google Scholar] [CrossRef]
- Delamuta, J.R.M.; Ribeiro, R.A.; Araújo, J.L.S.; Rouws, L.F.M.; Zilli, J.É.; Parma, M.M.; Melo, I.S.; Hungria, M. Bradyrhizobium stylosanthis sp. nov., comprising nitrogen-fixing symbionts isolated from nodules of the tropical forage legume Stylosanthes spp. Int. J. Syst. Evol. Microbiol. 2016, 66, 3078–3087. [Google Scholar] [CrossRef][Green Version]
- Rivas, R.; Velázquez, E.; Willems, A.; Vizcaíno, N.; Subba-Rao, N.S.; Mateos, P.F.; Gillis, M.; Dazzo, F.B.; Martínez-Molina, E. A New Species of Devosia That Forms a Unique Nitrogen-Fixing Root-Nodule Symbiosis with the Aquatic Legume Neptunia natans (L.f.) Druce. Appl. Environ. Microbiol. 2002, 68, 5217–5222. [Google Scholar] [CrossRef]
- Elliott, G.N.; Chen, W.M.; Chou, J.H.; Wang, H.C.; Sheu, S.Y.; Perin, L.; Reis, V.M.; Moulin, L.; Simon, M.F.; Bontemps, C.; et al. Burkholderia phymatum is a highly effective nitrogen-fixing symbiont of Mimosa spp. and fixes nitrogen ex planta. New Phytol. 2007, 173, 168–180. [Google Scholar] [CrossRef]
- Nguyen, V.Q. Symbiosis between Chamaecrista fasciculata and nitrogen-fixing bacteria: A review. Honors Thesis 2019, 189. [Google Scholar]
- Urquiaga, M.C.D.O.; Klepa, M.S.; Somasegaran, P.; Ribeiro, R.A.; Delamuta, J.R.M.; Hungria, M. Bradyrhizobium frederickii sp. nov., a nitrogen-fixing lineage isolated from nodules of the caesalpinioid species Chamaecrista fasciculata and characterized by tolerance to high temperature in vitro. Int. J. Syst. Evol. Microbiol. 2019, 69, 3863–3877. [Google Scholar] [CrossRef] [PubMed]
- Allen, O.N.; Allen, E.K. Response of the Peanut Plant to Inoculation with Rhizobia, with Special Reference to Morphological Development of the Nodules. Int. J. Plant Sci. 1940, 102, 121–142. [Google Scholar] [CrossRef]
- Corby, H.D.L. Types of rhizobial nodules and their distribution among Leguminosae. Kirkia 1988, 13, 53–124. [Google Scholar]
- Sen, D.; Weaver, R.W.; Bal, A.K. Structure and organisation of effective peanut and cowpea root nodules induced by rhizobial strain 32H1. J. Exp. Bot. 1986, 37, 356–363. [Google Scholar] [CrossRef]
- Bal, A.K.; Hameed, S.; Jayaram, S. Ultrastructural characteristics of the host-symbiont interface in nitrogen-fixing peanut nodules. Protoplasma 1989, 150, 19–26. [Google Scholar] [CrossRef]
- Limpens, E.; Bisseling, T. Signaling in symbiosis. Curr. Opin. Plant Biol. 2003, 6, 343–350. [Google Scholar] [CrossRef]
- Broughton, W.J.; Jabbouri, S.; Perret, X. Keys to Symbiotic Harmony. J. Bacteriol. 2000, 182, 5641–5652. [Google Scholar] [CrossRef]
- Spaink, H.P. Root Nodulation and Infection Factors Produced by Rhizobial Bacteria. Annu. Rev. Microbiol. 2000, 54, 257–288. [Google Scholar] [CrossRef]
- Noisangiam, R.; Teamtisong, K.; Tittabutr, P.; Boonkerd, N.; Toshiki, U.; Minamisawa, K.; Teaumroong, N. Genetic Diversity, Symbiotic Evolution, and Proposed Infection Process of Bradyrhizobium Strains Isolated from Root Nodules of Aeschynomene americana L. in Thailand. Appl. Environ. Microbiol. 2012, 78, 6236–6250. [Google Scholar] [CrossRef]
- Ibáñez, F.; Fabra, A. Rhizobial Nod factors are required for cortical cell division in the nodule morphogenetic programme of the Aeschynomeneae legume Arachis. Plant Biol. 2011, 13, 794–800. [Google Scholar] [CrossRef]
- Guha, S.; Sarkar, M.; Ganguly, P.; Uddin, R.; Mandal, S.; Dasgupta, M. Segregation of nod -containing and nod -deficient bradyrhizobia as endosymbionts of Arachis hypogaea and as endophytes of Oryza sativa in intercropped fields of Bengal Basin, India. Environ. Microbiol. 2016, 18, 2575–2590. [Google Scholar] [CrossRef] [PubMed]
- Madsen, E.B.; Antolín-Llovera, M.; Grossmann, C.; Ye, J.; Vieweg, S.; Broghammer, A.; Krusell, L.; Radutoiu, S.; Jensen, O.N.; Stougaard, J.; et al. Autophosphorylation is essential for the in vivo function of the Lotus japonicus Nod factor receptor 1 and receptor-mediated signalling in cooperation with Nod factor receptor 5. Plant J. 2011, 65, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Ibañez, F.; Angelini, J.; Figueredo, M.S.; Muñoz, V.; Tonelli, M.L.; Fabra, A. Sequence and expression analysis of putative Arachis hypogaea (peanut) Nod factor perception proteins. J. Plant Res. 2015, 128, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Stracke, S.; Kistner, C.; Yoshida, S.; Mulder, L.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; Stougaard, J.; Szczyglowski, K.; et al. A plant receptor-like kinase required for both bacterial and fungal symbiosis. Nature 2002, 417, 959–962. [Google Scholar] [CrossRef]
- Samaddar, S.; Dutta, A.; Sinharoy, S.; Paul, A.; Bhattacharya, A.; Saha, S.; Chien, K.-Y.; Goshe, M.B.; Dasgupta, M. Autophosphorylation of gatekeeper tyrosine by symbiosis receptor kinase. FEBS Lett. 2013, 587, 2972–2979. [Google Scholar] [CrossRef]
- Saha, S.; Dutta, A.; Bhattacharya, A.; Dasgupta, M. Intracellular Catalytic Domain of Symbiosis Receptor Kinase Hyperactivates Spontaneous Nodulation in Absence of Rhizobia1. Plant Physiol. 2014, 166, 1699–1708. [Google Scholar] [CrossRef]
- Saha, S.; Paul, A.; Herring, L.; Dutta, A.; Bhattacharya, A.; Samaddar, S.; Goshe, M.B.; Das Gupta, M. Gatekeeper Tyrosine Phosphorylation of SYMRK Is Essential for Synchronizing the Epidermal and Cortical Responses in Root Nodule Symbiosis1. Plant Physiol. 2016, 171, 71–81. [Google Scholar] [CrossRef]
- Lévy, J.; Bres, C.; Geurts, R.; Chalhoub, B.; Kulikova, O.; Duc, G.; Journet, E.P.; Ané, J.M.; Lauber, E.; Bisseling, T.; et al. A putative Ca2+ and calmodulin-dependent protein kinase required for bacterial and fungal symbioses. Science 2004, 303, 1361–1364. [Google Scholar]
- Mitra, R.M.; Gleason, C.A.; Edwards, A.; Hadfield, J.; Downie, J.A.; Oldroyd, G.E.D.; Long, S.R. A Ca2+/calmodulin-dependent protein kinase required for symbiotic nodule development: Gene identification by transcript-based cloning. Proc. Natl. Acad. Sci. USA 2004, 101, 4701–4705. [Google Scholar] [CrossRef]
- Sinharoy, S.; Dasgupta, M. RNA Interference Highlights the Role of CCaMK in Dissemination of Endosymbionts in the Aeschynomeneae Legume Arachis. Mol. Plant-Microbe Interact. 2009, 22, 1466–1475. [Google Scholar] [CrossRef]
- Yano, K.; Yoshida, S.; Müller, J.; Singh, S.; Banba, M.; Vickers, K.; Markmann, K.; White, C.; Schuller, B.; Sato, S.; et al. CYCLOPS, a mediator of symbiotic intracellular accommodation. Proc. Natl. Acad. Sci. USA 2008, 105, 20540–20545. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Katzer, K.; Lambert, J.; Cerri, M.; Parniske, M. CYCLOPS, A DNA-Binding Transcriptional Activator, Orchestrates Symbiotic Root Nodule Development. Cell Host Microbe 2014, 15, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Cerri, M.R.; Wang, Q.; Stolz, P.; Folgmann, J.; Frances, L.; Katzer, K.; Li, X.; Heckmann, A.B.; Wang, T.L.; Downie, J.A.; et al. The ERN 1 transcription factor gene is a target of the CC a MK/CYCLOPS complex and controls rhizobial infection in Lotus japonicus. New Phytol. 2017, 215, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Genet. 2013, 11, 252–263. [Google Scholar] [CrossRef]
- Das, D.R.; Horváth, B.; Kundu, A.; Kaló, P.; Dasgupta, M. Functional conservation of CYCLOPS in crack entry legume Arachis hypogaea. Plant Sci. 2019, 281, 232–241. [Google Scholar] [CrossRef]
- Gonzalez-Rizzo, S.; Crespi, M.; Frugier, F. The Medicago truncatula CRE1 Cytokinin Receptor Regulates Lateral Root Development and Early Symbiotic Interaction with Sinorhizobium meliloti. Plant Cell 2006, 18, 2680–2693. [Google Scholar] [CrossRef]
- Tirichine, L.; Sandal, N.; Madsen, L.H.; Radutoiu, S.; Albrektsen, A.S.; Sato, S.; Asamizu, E.; Tabata, S.; Stougaard, J. A Gain-of-Function Mutation in a Cytokinin Receptor Triggers Spontaneous Root Nodule Organogenesis. Science 2007, 315, 104–107. [Google Scholar] [CrossRef]
- Murray, J.; Karas, B.J.; Sato, S.; Tabata, S.; Amyot, L.; Szczyglowski, K. A Cytokinin Perception Mutant Colonized by Rhizobium in the Absence of Nodule Organogenesis. Science 2007, 315, 101–104. [Google Scholar] [CrossRef]
- Plet, J.; Wasson, A.; Ariel, F.; Le Signor, C.; Baker, D.; Mathesius, U.; Crespi, M.; Frugier, F. MtCRE1-dependent cytokinin signaling integrates bacterial and plant cues to coordinate symbiotic nodule organogenesis in Medicago truncatula. Plant J. 2011, 65, 622–633. [Google Scholar] [CrossRef]
- Kundu, A.; DasGupta, M. Silencing of putative cytokinin receptor histidine kinase1 inhibits both inception and differentiation of root nodules in Arachis hypogaea. Mol. Plant-Microbe. Interact. 2017, 31, 187–199. [Google Scholar] [CrossRef]
- Ng, J.L.P.; Hassan, S.; Truong, T.T.; Hocart, C.H.; Laffont, C.; Frugier, F.; Mathesius, U. Flavonoids and Auxin Transport Inhibitors Rescue Symbiotic Nodulation in the Medicago truncatula Cytokinin Perception Mutant cre1. Plant Cell 2015, 27, 2210–2226. [Google Scholar] [CrossRef] [PubMed]
- Breakspear, A.; Liu, C.; Roy, S.; Stacey, N.; Rogers, C.; Trick, M.; Morieri, G.; Mysore, K.S.; Wen, J.; Oldroyd, G.E.; et al. The root hair “infectome” of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for Auxin signaling in rhizobial infection. Plant Cell 2014, 26, 4680–4701. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Maekawa-Yoshikawa, M.; Takeda, N.; Imaizumi-Anraku, H.; Murooka, Y.; Hayashi, M. Gibberellin controls the nodulation signaling pathway in Lotus japonicus. Plant J. 2009, 58, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Asamizu, E.; Shimoda, Y.; Kouchi, H.; Tabata, S.; Sato, S. A Positive Regulatory Role for LjERF1 in the Nodulation Process Is Revealed by Systematic Analysis of Nodule-Associated Transcription Factors of Lotus japonicus. Plant Physiol. 2008, 147, 2030–2040. [Google Scholar] [CrossRef] [PubMed]
- Penmetsa, R.V.; Uribe, P.; Anderson, J.; Lichtenzveig, J.; Gish, J.C.; Nam, Y.W.; Engstrom, E.; Xu, K.; Sckisel, G.; Pereira, M.; et al. The Medicago truncatula ortholog of Arabidopsis EIN2, sickle, is a negative regulator of symbiotic and pathogenic microbial associations. Plant J. 2008, 55, 580–595. [Google Scholar] [CrossRef]
- Liang, Y.; Harris, J.M. Response of root branching to abscisic acid is correlated with nodule formation both in legumes and non -legumes. Am. J. Bot. 2005, 92, 1675–1683. [Google Scholar] [CrossRef]
- Ding, Y.; Kalo, P.; Yendrek, C.; Sun, J.; Liang, Y.; Marsh, J.F.; Harris, J.M.; Oldroyd, G.E. Abscisic Acid Coordinates Nod Factor and Cytokinin Signaling during the Regulation of Nodulation in Medicago truncatula. Plant Cell 2008, 20, 2681–2695. [Google Scholar] [CrossRef]
- Jardinaud, M.-F.; Boivin, S.; Rodde, N.; Catrice, O.; Kisiala, A.; Lepage, A.; Moreau, S.; Roux, B.; Cottret, L.; Sallet, E.; et al. A Laser Dissection-RNAseq Analysis Highlights the Activation of Cytokinin Pathways by Nod Factors in the Medicago truncatula Root Epidermis. Plant Physiol. 2016, 171, 2256–2276. [Google Scholar] [CrossRef]
- Herrbach, V.; Chirinos, X.; Rengel, D.; Agbevenou, K.; Vincent, R.; Pateyron, S.; Huguet, S.; Balzergue, S.; Pasha, A.; Provart, N.; et al. Nod factors potentiate auxin signaling for transcriptional regulation and lateral root formation in Medicago truncatula. J. Exp. Bot. 2017, 68, 569–583. [Google Scholar]
- Held, M.; Hou, H.; Miri, M.; Huynh, C.; Ross, L.; Hossain, S.; Sato, S.; Tabata, S.; Perry, J.; Wang, T.L.; et al. Lotus japonicus cytokinin receptors work partially redundantly to mediate nodule formation. Plant Cell 2014, 26, 678–694. [Google Scholar] [CrossRef]
- Frugier, F.; Kosuta, S.; Murray, J.D.; Crespi, M.; Szczyglowski, K. Cytokinin: Secret agent of symbiosis. Trends Plant Sci. 2008, 13, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Mathesius, U.; Charon, C.; Rolfe, B.G.; Kondorosi, A.; Crespi, M. Temporal and spatial order of events during the induction of cortical cell divisions in white clover by Rhizobium leguminosarum bv. trifolii inoculation or localized cytokinin addition. Mol. Plant-Microbe. Interact. 2000, 13, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Suzaki, T.; Yano, K.; Ito, M.; Umehara, Y.; Suganuma, N.; Kawaguchi, M. Positive and negative regulation of cortical cell division during root nodule development in Lotus japonicus is accompanied by auxin response. Development 2012, 139, 3997–4006. [Google Scholar] [CrossRef] [PubMed]
- Perilli, S.; Pérez-Pérez, J.M.; Di Mambro, R.; Peris, C.L.; Diaz-Trivino, S.; Del Bianco, M.; Pierdonati, E.; Moubayidin, L.; Cruz-Ramírez, A.; Costantino, P.; et al. Retinoblastoma-related Protein Stimulates Cell Differentiation in the Arabidopsis Root Meristem by Interacting with Cytokinin Signaling. Plant Cell 2013, 25, 4469–4478. [Google Scholar] [CrossRef]
- Franssen, H.J.; Xiao, T.T.; Kulikova, O.; Wan, X.; Bisseling, T.; Scheres, B.; Heidstra, R. Root developmental programs shape the Medicago truncatula nodule meristem. Development 2015, 142, 2941–2950. [Google Scholar] [CrossRef]
- Fonouni-Farde, C.; Kisiala, A.; Brault, M.; Emery, R.N.; Diet, A.; Frugier, F. DELLA1-Mediated Gibberellin Signaling Regulates Cytokinin-Dependent Symbiotic Nodulation. Plant Physiol. 2017, 175, 1795–1806. [Google Scholar] [CrossRef]
- Vernie, T.; Moreau, S.; de Billy, F.; Plet, J.; Combier, J.P.; Rogers, C.; Oldroyd, G.; Frugier, F.; Niebel, A.; Gamas, P. EFD Is an ERF Transcription Factor Involved in the Control of Nodule Number and Differentiation in Medicago truncatula. Plant Cell 2008, 20, 2696–2713. [Google Scholar] [CrossRef]
- Zaat, S.A.J.; Van Brussel, A.A.N.; Tak, T.; Lugtenberg, B.J.J.; Kijne, J.W. The ethylene-inhibitor aminoethoxyvinylglycine restores normal nodulation by Rhizobium leguminosarum biovar. viciae on Vicia sativa subsp. nigra by suppressing the ‘Thick and short roots’ phenotype. Planta 1989, 177, 141–150. [Google Scholar] [CrossRef]
- Heidstra, R.; Yang, W.C.; Yalcin, Y.; Peck, S.; Emons, A.M.; Van Kammen, A.; Bisseling, T. Ethylene provides positional information on cortical cell division but is not involved in Nod factor-induced root hair tip growth in Rhizobium-legume interaction. Development 1997, 124, 1781–1787. [Google Scholar]
- Oldroyd, G.E.; Engstrom, E.M.; Long, S.R. Ethylene inhibits the Nod factor signal transduction pathway of Medicago truncatula. Plant Cell 2001, 13, 1835–1849. [Google Scholar] [CrossRef]
- Giraudat, J.; Parcy, F.; Bertauche, N.; Gosti, F.; Leung, J.; Morris, P.C.; Bouvier-Durand, M.; Vartanian, N. Current advances in abscisic acid action and signalling. Plant Mol. Biol. 1994, 26, 1557–1577. [Google Scholar] [CrossRef] [PubMed]
- De Smet, I.; Signora, L.; Beeckman, T.; Inzé, D.; Foyer, C.H.; Zhang, H. An abscisic acid-sensitive checkpoint in lateral root development of Arabidopsis. Plant J. 2003, 33, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Rosenblueth, M.; Ormeño-Orrillo, E.; López-López, A.; Rogel, M.A.; Reyes-Hernández, B.J.; Martínez-Romero, J.C.; Reddy, P.M.; Martínez-Romero, E. Nitrogen Fixation in Cereals. Front. Microbiol. 2018, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, M.; Duan, L.; Zhang, T.; Cao, Y.; Zhang, Z. Domain Swap Approach Reveals the Critical Roles of Different Domains of SYMRK in Root Nodule Symbiosis in Lotus japonicus. Front. Plant Sci. 2018, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Rubio, M.C.; Xin, X.; Zhang, B.; Fan, Q.; Wang, Q.; Ning, G.; Becana, M.; Duanmu, D. CRISPR/Cas9 knockout of leghemoglobin genes in Lotus japonicus uncovers their synergistic roles in symbiotic nitrogen fixation. New Phytol. 2019, 224, 818–832. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.Y.; Dickerson, J.; Xu, N. Bioinformatics and its applications in plant biology. Annu. Rev. Plant Biol. 2006, 57, 335–360. [Google Scholar] [CrossRef]
- Pandey, M.K.; Monyo, E.; Ozias-Akins, P.; Liang, X.; Guimarães, P.; Nigam, S.N.; Upadhyaya, H.D.; Janila, P.; Zhang, X.; Guo, B.; et al. Advances in Arachis genomics for peanut improvement. Biotechnol. Adv. 2012, 30, 639–651. [Google Scholar] [CrossRef]
- Duan, X.; Schmidt, E.; Li, P.; Lenox, D.; Liu, L.; Shu, C.; Zhang, J.; Liang, C. PeanutDB: An integrated bioinformatics web portal for Arachis hypogaea transcriptomics. BMC Plant Biol. 2012, 12, 94. [Google Scholar] [CrossRef]
- Campbell, M.S.; Law, M.; Holt, C.; Stein, J.C.; Moghe, G.D.; Hufnagel, D.E.; Lei, J.; Achawanantakun, R.; Jiao, D.; Lawrence, C.J.; et al. MAKER-p: A tool kit for the rapid creation, management, and quality control of plant genome annotations. Plant Physiol. 2014, 64, 513–524. [Google Scholar] [CrossRef]
- Dash, S.; Cannon, E.K.; Kalberer, S.R.; Farmer, A.D.; Cannon, S.B. PeanutBase and Other Bioinformatic Resources for Peanut; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 241–252. [Google Scholar]
- Rogers, C.; Oldroyd, G.E.D. Synthetic biology approaches to engineering the nitrogen symbiosis in cereals. J. Exp. Bot. 2014, 65, 1939–1946. [Google Scholar] [CrossRef]
- Delaux, P.M.; Radhakrishnan, G.; Oldroyd, G. Tracing the evolutionary path to nitrogen-fixing crops. Curr. Opin. Plant Biol. 2015, 26, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Mondy, S.; Lenglet, A.; Libanga, C.; Ratet, P.; Faure, D.; Dessaux, Y.; Beury-Cirou, A. An increasing opine carbon bias in artificial exudation systems and genetically modified plant rhizospheres leads to an increasing reshaping of bacterial populations. Mol. Ecol. 2014, 23, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.; Steadman, C.E.; Stevenson, D.; Coyle, M.; Rees, R.M.; Skiba, U.M.; Sutton, M.A.; Cape, J.N.; Dore, A.J.; Vieno, M.; et al. Effects of global change during the 21st century on the nitrogen cycle. Atmos. Chem. Phys. 2015, 15, 13849–13893. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef]
- Xu, R.T.; Pan, S.F.; Chen, J.; Chen, G.S.; Yang, J.; Dangal, S.R.S.; Shepard, J.P.; Tian, H.Q. Half-century ammonia emissions from agricultural systems in Southern Asia: Magnitude, spatiotemporal patterns, and implications for human health. GeoHealth 2017, 2, 40–53. [Google Scholar] [CrossRef]
- Francis, C.A. Macmillan publishing company, New york, USA. 1986, 1-17. In Distribution and Importance of Multiple Cropping. Cropping System; Macmillan publishing company: New york, NY, USA, 1986; pp. 1–17. [Google Scholar]
- Bodade, V.N. Mixed cropping of groundnut and Jowar. Indian Oil Seed J. 1964, 8, 297–301. [Google Scholar]
- Enyi, B.A.C. Effects of intercropping maize or sorghum with cowpeas, pigeonpeas or beans. Exp. Agr. 1973, 9, 83–90. [Google Scholar] [CrossRef]
- Moynihan, J.M.; Simmons, S.R.; Sheaffer, C.C. Intercropping annual medic with Conventional height and semi-dwarf barley grown for grain. Agron. J. 1996, 88, 823–828. [Google Scholar] [CrossRef]
- Ghosh, P.K.; Bandyopadhyay, K.K.; Wanjari, R.H.; Manna, M.C.; Misra, A.K.; Mohanty, M.; Rao, A.S. Legume Effect for Enhancing Productivity and Nutrient Use-Efficiency in Major Cropping Systems–An Indian Perspective: A Review. J. Sustain. Agric. 2007, 30, 59–86. [Google Scholar] [CrossRef]
- Chu, G.; Shen, Q.; Cao, J. Nitrogen fixation and N transfer from peanut to rice cultivated in aerobic soil in an intercropping system and its effect on soil N fertility. Plant Soil 2004, 263, 17–27. [Google Scholar] [CrossRef]
- LaRue, T.A. Selecting and Breeding Legumes for Enhanced Nitrogen Fixation (Recommendations for Research and Proceedings of a Workshop); Boyce Thompson Institute at Cornell: Ithaca, NY, USA, 1978; p. 23. [Google Scholar]
- Nutman, P.S. Improving nitrogen fixation in legumes by plant breeding: The relevance of host selection experiments in red clover (Trifolium pratense L.) and subterranean clover (T. subterraneum L.). Plant Soil 1984, 82, 285–301. [Google Scholar] [CrossRef]
- Mytton, L.R. Host plant selection and breeding for improved symbiotic efficiency. In The Physiology, Genetics and Nodulation of Temperate Legumes; Jones, D.G., Davies, D.R., Eds.; Pitman: London, UK, 1983; pp. 373–393. [Google Scholar]
- Graham, P.H.; Temple, S.R. Selection for improved nitrogen fixation in Glycine max (L.) Merr. and Phaseolus vulgaris L. Plant Soil 1984, 82, 315–327. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Gao, D.; Dudchenko, O.; Seijo, G.; Leal-Bertioli, S.; Ren, L.; Farmer, A.; Pandey, M.K.; et al. The genome sequence of peanut (Arachis hypogaea), a segmental allotetraploid. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, Q.; Liu, H.; Zhang, J.; Hong, Y.; Lan, H.; Li, H.; Wang, J.; Liu, H.; Li, S.; et al. Sequencing of Cultivated Peanut, Arachis hypogaea, Yields Insights into Genome Evolution and Oil Improvement. Mol. Plant 2019, 12, 920–934. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.; Pandey, M.K.; Zhang, C.; Chang, W.C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The Arachis hypogaea genome elucidates legume karyotypes, polyploid evolution and crop domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.K.; Agarwal, G.; Kale, S.M.; Clevenger, J.; Nayak, S.N.; Sriswathi, M.; Chitikineni, A.; Chavarro, C.; Chen, X.; Upadhyaya, H.D.; et al. Development and Evaluation of a High Density Genotyping ‘Axiom_Arachis’ Array with 58 K SNPs for Accelerating Genetics and Breeding in Groundnut. Sci. Rep. 2017, 7, 40577. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ‘Crack-entry’ Legumes | Systematic Position within Leguminosae (Fabaceae) | Symbiotic Partner Rhizobia | Distinctive Features of Nodule | Reference |
|---|---|---|---|---|
| Arachis hypogaea | Sub family: Papilionoideae Tribe: Dalbergieae (Dalbergioid) | Bradyrhizobium sp. | Aeschynomenoid type, oblate-spheroid, root junctions with axillary hairs | [67] |
| Sesbania rostrata | Sub family: Papilionoideae Tribe: Sesbanieae (Robinioid) | Azorhizobium caulinodans | Aeschynomenoid affinities, spherical, ‘open basket’ nodule meristem. | [68] |
| Stylosanthes guianensis | Sub family: Papilionoideae Tribe: Dalbergieae (Dalbergioid) | Bradyrhizobium stylosanthis Rhizobium sp. Strains: CIAT1460, CB2126, CB1650. | Aeschynomenoid type, oblate, root junctions with axillary hairs. | [69,70] |
| Aeschynomene afraspera | Sub family: Papilionoideae Tribe: Dalbergieae (Dalbergioid) | Bradyrhizobium sp. | Aeschynomenoid type, spheroid, root and stem nodules. | [12] |
| Neptunia natans | Sub family: Mimosoideae Tribe: Mimoseae (Mimosoid, MCC) | Devosia riboflavina | Mimosoid, unbranched- elongated, aquatic infection environment. | [71] |
| Mimosa pudica | Sub family: Mimosoideae Tribe: Mimoseae (Mimosoid, MCC) | Burkholderia phymatum, Cupravidus taiwanensis | Mimosoid, unbranched- elongated, broad symbiont range. | [72] |
| Chamaecrista fasciculata | Sub family: Caesalpinioideae Tribe: Cassieae (MCC) | Burkholderia tuberum, Rhizobium tropici Bradyrhizobium frederickii | Caesalpiniod type, rigid, hemispherical with fixation threads. | [73,74] |
| RNS Progress | Key Genes /TFs | Functional Description |
|---|---|---|
| Bacterial Recognition | NFR1/LYK3 | Initiate host response |
| LYR3 | Recognition of symbiotic signals | |
| EPR3 | Role in Recognition of Nod factor and host-symbiont compatibility | |
| Early signalling and SYM pathway | CASTOR | Encode putative ion channel protein |
| CNGC | Plays a role in symbiotic calcium oscillations in SYM pathway | |
| CYCLOPS | DNA-binding transcriptional activator induces nodule development | |
| Early Transcription factor | NIN, NSP2, and ERF1 | Role in transcriptional reprogramming for initiation of root nodule symbiosis |
| ERN1 | Controls rhizobial infection | |
| Infection | ARPC1 | Encode heptameric ARP2/3 nucleator—essential for the intracellular accommodation of rhizobial bacteria |
| CERBERUS | Role in infection thread formation, growth, and differentiation of nodules | |
| Cell division | HK1/CRE1 | Role in nodule organogenesis |
| bHLH476 | Plays a role in cytokinin pathway which positively regulates symbiotic nodulation | |
| Nodule regulation | EIN2 | Plays a key role in plant–microbe interaction |
| AP2 | Transcriptional regulator of symbiotic nodule development | |
| Cytokinin oxidase/dehydrogenase 6 | Maintaining cytokinin homeostasis during root and nodule development | |
| CLE13 | Role in systematic autoregulation of nodulation (AON) pathway that negatively regulates nodule number | |
| F-box/kelch-repeat protein | Long-distance regulation of legume–rhizobium symbiosis | |
| Protein kinase superfamily protein | Role in infection and nodule development | |
| RWP-RK family protein | Key regulators of nitrogen responses and of gametophyte development |
| Plant Hormones | Overall Influence on Nodulation | Mechanism of Regulation | Reference |
|---|---|---|---|
| Cytokinin | Positive | ● Ligand for HK1, essential for cortical signalling ● Delimits polar auxin transport during growth of nodule primordia | [101] |
| Auxin | Positive | ● Required in nodule meristem for cortical division ● Regulated by NF signalling | [104,105] |
| Gibberellin | Negative | ● Nod factor triggered negative feedback maintenance | [106] |
| Ethylene | Negative / Positive | ● Downregulates defense response gene Lj Pr-10 ● Downregulates early genes of symbiotic pathway | [107,108] |
| Abscisic acid | Negative / Positive | ● Increases lateral root density (LDR) in legumes ● Negatively regulates ENOD 11 and RIP1 | [109,110] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, V.; Bhattacharyya, S.; Kumar, R.; Kumar, A.; Ibañez, F.; Wang, J.; Guo, B.; Sudini, H.K.; Gopalakrishnan, S.; DasGupta, M.; et al. Molecular Basis of Root Nodule Symbiosis between Bradyrhizobium and ‘Crack-Entry’ Legume Groundnut (Arachis hypogaea L.). Plants 2020, 9, 276. https://doi.org/10.3390/plants9020276
Sharma V, Bhattacharyya S, Kumar R, Kumar A, Ibañez F, Wang J, Guo B, Sudini HK, Gopalakrishnan S, DasGupta M, et al. Molecular Basis of Root Nodule Symbiosis between Bradyrhizobium and ‘Crack-Entry’ Legume Groundnut (Arachis hypogaea L.). Plants. 2020; 9(2):276. https://doi.org/10.3390/plants9020276
Chicago/Turabian StyleSharma, Vinay, Samrat Bhattacharyya, Rakesh Kumar, Ashish Kumar, Fernando Ibañez, Jianping Wang, Baozhu Guo, Hari K. Sudini, Subramaniam Gopalakrishnan, Maitrayee DasGupta, and et al. 2020. "Molecular Basis of Root Nodule Symbiosis between Bradyrhizobium and ‘Crack-Entry’ Legume Groundnut (Arachis hypogaea L.)" Plants 9, no. 2: 276. https://doi.org/10.3390/plants9020276
APA StyleSharma, V., Bhattacharyya, S., Kumar, R., Kumar, A., Ibañez, F., Wang, J., Guo, B., Sudini, H. K., Gopalakrishnan, S., DasGupta, M., Varshney, R. K., & Pandey, M. K. (2020). Molecular Basis of Root Nodule Symbiosis between Bradyrhizobium and ‘Crack-Entry’ Legume Groundnut (Arachis hypogaea L.). Plants, 9(2), 276. https://doi.org/10.3390/plants9020276