High-Temperature and Drought-Resilience Traits among Interspecific Chromosome Substitution Lines for Genetic Improvement of Upland Cotton

 , ,
, ,  ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Results and Discussion
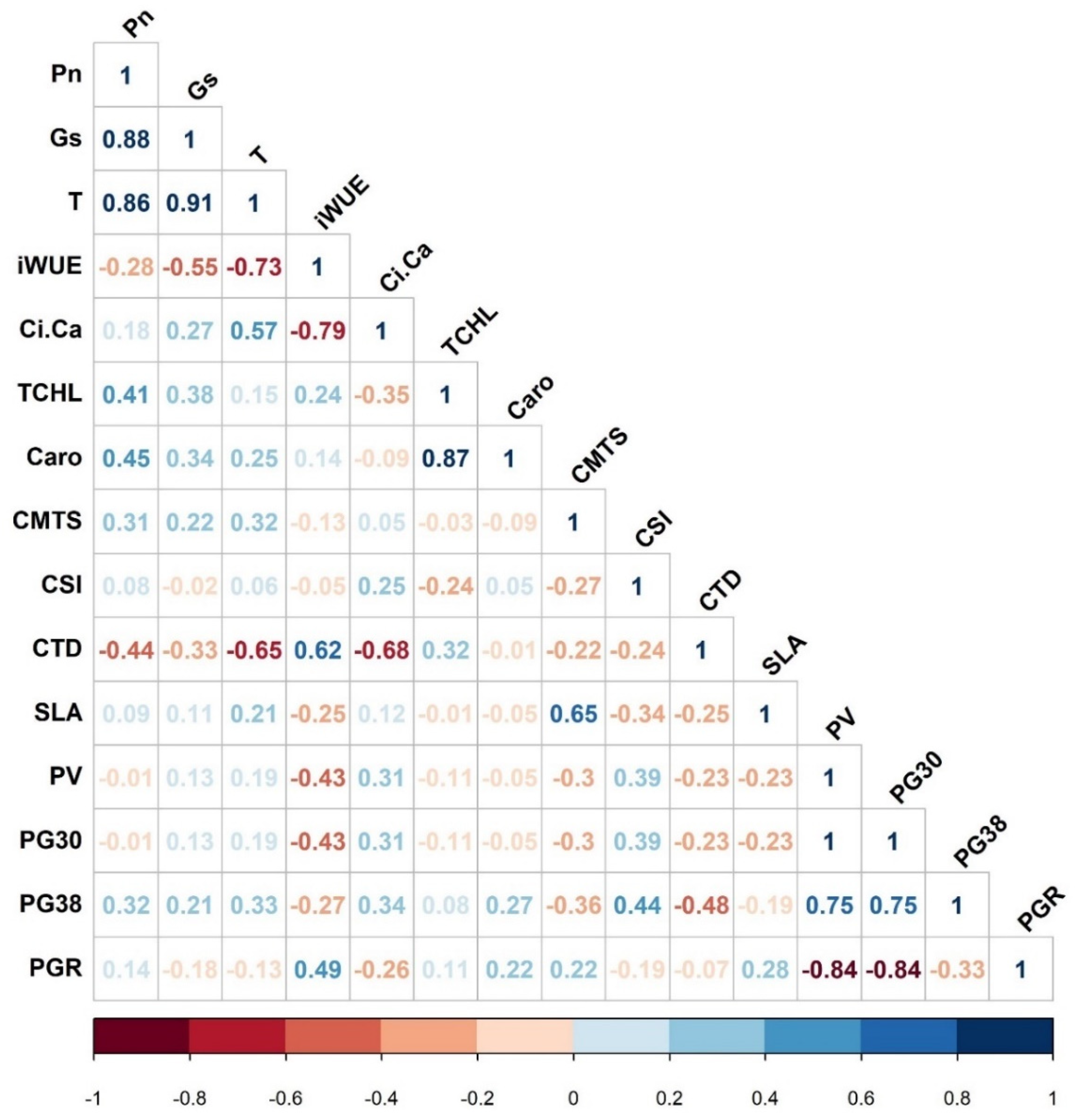
2.1. Leaf Gas Exchange and Pigment Parameters
2.2. Biophysical and Temperature-Tolerant Indices
2.3. Pollen Viability and Germination Parameters
2.4. Promising CS Lines for Future Studies
2.5. Comparative Analysis of CS Lines Physiological and Pollen Phenotype and Modern Advanced Upland Cotton Cultivars
2.6. Compositional Analysis
2.7. Outlook
3. Materials and Methods
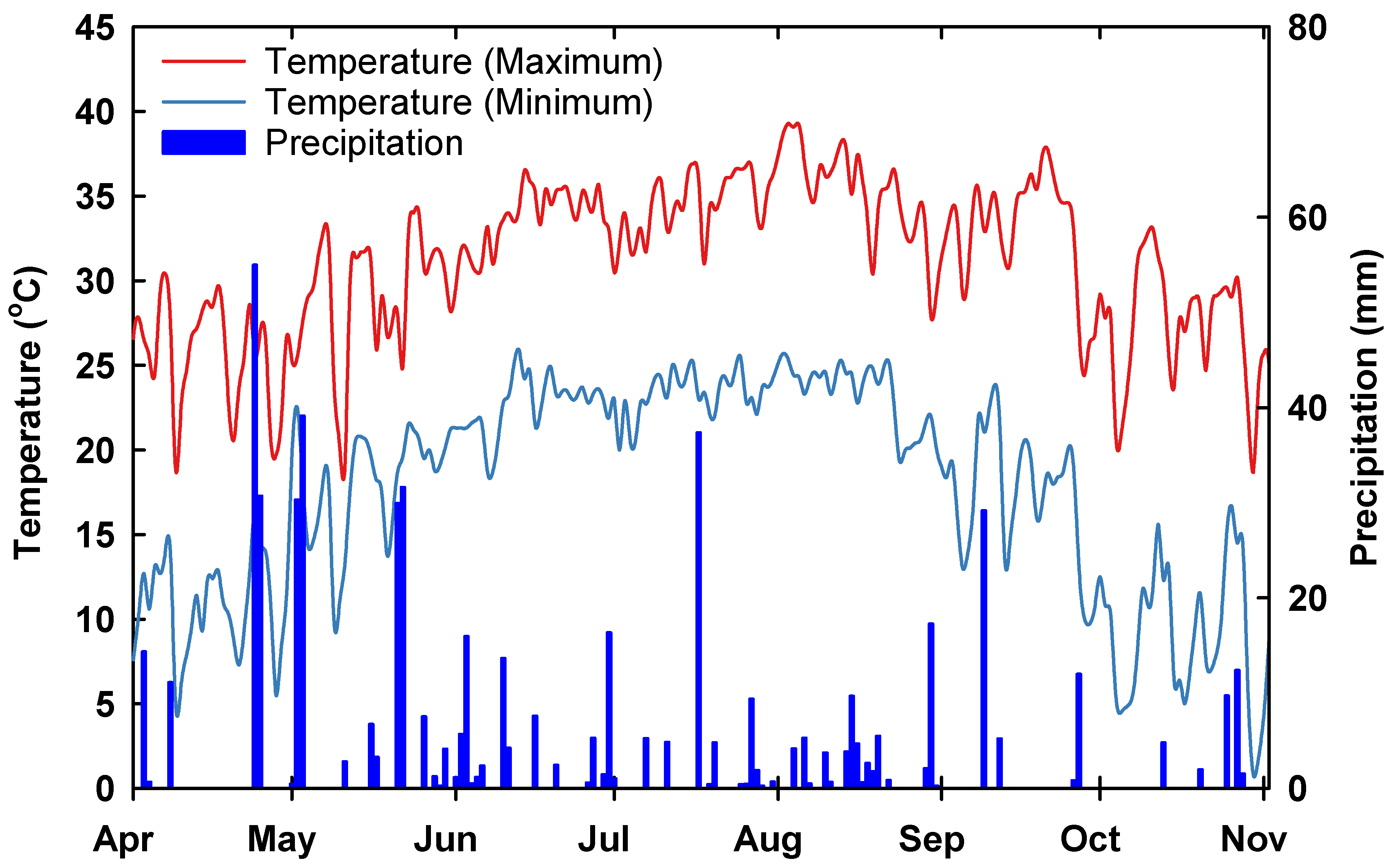
3.1. Planting Materials
3.2. Pollen Viability under Field Conditions and Germination Performance under High-Temperature Stress
3.3. Gas Exchange Parameters
3.4. Leaf Pigment Content and Chlorophyll Stability Index (CSI)
3.5. Leaf Cell Membrane Thermostability (CMTS)
3.6. Canopy Temperature Depression
3.7. Specific Leaf Area
3.8. SSR Analysis
3.9. SNP Analysis
3.10. Stress Response Indices and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Car | Carotenoids |
| Ci/Ca | Ratio of leaf internal to ambient carbon dioxide concentration |
| CMTS | Cell membrane thermostability |
| CSI | Chlorophyll stability index |
| CTD | Canopy temperature depression |
| DSRI | Drought stress response index |
| Gs | Stomatal conductance |
| HSRI | Heat stress response index |
| iWUE | Instantaneous leaf water use efficiency |
| PG | Pollen germination |
| PGR | Pollen germination response |
| Pn | Leaf net photosynthesis |
| PV | Pollen viability |
| SLA | Specific leaf area |
| SNP | Single nucleotide polymorphism |
| SSR | Simple sequence repeat |
| TCHL | Total leaf chlorophyll |
| T | Transpiration |
References
- ICAC. Global warming and cotton production—Part 2. ICAC Rec. 2009, 27, 9–13. [Google Scholar]
- Zahid, K.R.; Ali, F.; Shah, F.; Younas, M.; Shah, T.; Shahwar, D.; Hassan, W.; Ahmad, Z.; Qi, C.; Lu, Y. Response and tolerance mechanism of cotton Gossypium hirsutum L. to elevated temperature stress: A review. Front. Plant Sci. 2016, 7, 937. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- FAO. World Food and Agriculture; Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Kukal, M.S.; Irmak, S. Climate-driven crop yield and yield variability and climate change impacts on the US Great Plains agricultural production. Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef]
- Ehlig, C.; LeMert, R. Effects of Fruit Load, Temperature, and relative humidity on boll retention of cotton 1. Crop Sci. 1973, 13, 168–171. [Google Scholar] [CrossRef]
- Khan, A.; Pan, X.; Najeeb, U.; Tan, D.K.Y.; Fahad, S.; Zahoor, R.; Luo, H. Coping with drought: Stress and adaptive mechanisms, and management through cultural and molecular alternatives in cotton as vital constituents for plant stress resilience and fitness. Biol. Res. 2018, 51, 47. [Google Scholar] [CrossRef]
- Sabagh, A.E.; Hossain, A.; Islam, M.S.; Barutcular, C.; Ratnasekera, D.; Gormus, O.; Amanet, K.; Mubeen, M.; Nasim, W.; Fahad, S. Drought and heat stress in cotton (Gossypium hirsutum L.): Consequences and their possible mitigation strategies. In Agronomic Crops; Springer: Berlin/Hedelberg, Germany, 2020; pp. 613–634. [Google Scholar]
- Reddy, K.R.; Hodges, H.F.; Reddy, V.R. Temperature effects on cotton fruit retention. Agron. J. 1992, 84, 26–30. [Google Scholar] [CrossRef]
- Pettigrew, W. The effect of higher temperatures on cotton lint yield production and fiber quality. Crop Sci. 2008, 48, 278–285. [Google Scholar] [CrossRef]
- Bheemanahalli, R.; Sathishraj, R.; Tack, J.; Nalley, L.L.; Muthurajan, R.; Jagadish, K.S. Temperature thresholds for spikelet sterility and associated warming impacts for sub-tropical rice. Agric. For. Meteorol. 2016, 221, 122–130. [Google Scholar] [CrossRef]
- Reddy, K.R.; Reddy, V.R.; Hodges, H.F. Temperature effects on early season cotton growth and development. Agron. J. 1992, 84, 229–237. [Google Scholar] [CrossRef]
- Soliz, L.M.A.; Oosterhuis, D.M.; Coker, D.L.; Brown, R.S. Physiological response of cotton to high night temperature. Am. J. Sci. Biotechnol. 2008, 2, 63–68. [Google Scholar]
- Jain, M.; Prasad, P.V.; Boote, K.J.; Hartwell, A.L.; Chourey, P.S. Effects of season-long high temperature growth conditions on sugar-to-starch metabolism in developing microspores of grain sorghum (Sorghum bicolor L. Moench). Planta 2007, 227, 67–79. [Google Scholar] [CrossRef]
- Saini, H.; Sedgley, M.; Aspinall, D. Effect of heat stress during floral development on pollen tube growth and ovary anatomy in wheat (Triticum aestivum L.). Funct. Plant Biol. 1983, 10, 137–144. [Google Scholar] [CrossRef]
- Kakani, V.G.; Reddy, K.R.; Koti, S.; Wallace, T.; Prasad, P.V.; Reddy, V.R.; Zhao, D. Differences in in vitro pollen germination and pollen tube growth of cotton cultivars in response to high temperature. Ann. Bot. 2005, 96, 59–67. [Google Scholar] [CrossRef]
- Hedhly, A.; Hormaza, J.; Herrero, M. Effect of temperature on pollen tube kinetics and dynamics in sweet cherry, Prunus avium (Rosaceae). Am. J. Bot. 2004, 91, 558–564. [Google Scholar] [CrossRef]
- Reddy, V.R.; Reddy, K.R.; Baker, D.N. Temperature effect on growth and development of cotton during the fruiting period. Agron. J. 1991, 83, 211–217. [Google Scholar] [CrossRef]
- Reddy, K.R.; Hodges, H.F.; McKinion, J.M. A temperature model for cotton phenology. Biotronics 1993, 22, 47–59. [Google Scholar]
- Reddy, K.; Hodges, H.; McKinion, J. Carbon dioxide and temperature effects on pima cotton growth. Agric. Ecosyst. Environ. 1995, 54, 17–29. [Google Scholar] [CrossRef]
- Ulloa, M.; De Santiago, L.M.; Hulse-Kemp, A.M.; Stelly, D.M.; Burke, J.J. Enhancing Upland cotton for drought resilience, productivity, and fiber quality: Comparative evaluation and genetic dissection. Mol. Genet. Genom. 2020, 295, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Witt, T.W.; Ulloa, M.; Schwartz, R.C.; Ritchie, G.L. Response to deficit irrigation of morphological, yield and fiber quality traits of upland (Gossypium hirsutum L.) and Pima (G. barbadense L.) cotton in the Texas High Plains. Field Crop. Res. 2020, 249, 107759. [Google Scholar] [CrossRef]
- Saha, S.; Jenkins, J.N.; Wu, J.; McCarty, J.C.; Gutiérrez, O.A.; Percy, R.G.; Cantrell, R.G.; Stelly, D.M. Effects of chromosome-specific introgression in upland cotton on fiber and agronomic traits. Genetics 2006, 172, 1927–1938. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Percival, A.; Wendel, J.; Stewart, J. Taxonomy and Germplasm Resources; Agronomy Monographs; John Wiley and Sons, Inc.: New York, NY, USA, 1999. [Google Scholar] [CrossRef]
- Stelly, D.; Saha, S.; Raska, D.; Jenkins, J.; McCarty, J., Jr.; Gutierrez, O. Registration of 17 upland (Gossypium hirsutum) cotton germplasm lines disomic for different G. barbadense chromosome or arm substitutions. Crop Sci. 2005, 45, 263–265. [Google Scholar] [CrossRef]
- Jenkins, J.N.; McCarty, J.C.; Campbell, B.T.; Hayes, R.; Wu, J.; Saha, S.; Stelly, D. Genotypic comparisons of chromosomes 01, 04, and 18 from three tetraploid species of Gossypium in topcrosses with five elite cultivars of G. hirsutum L. Euphytica 2017, 213, 107. [Google Scholar] [CrossRef]
- Jenkins, J.N.; McCarty, J.C., Jr.; Campbell, B.T.; Hayes, R.; Wu, J.; Saha, S.; Stelly, D. Genetic effects of chromosomes 1, 4, and 18 from three tetraploid Gossypium species in topcrosses with five elite cultivars. Crop Sci. 2017, 57, 1338–1346. [Google Scholar] [CrossRef]
- Saha, S.; Wu, J.; Jenkins, J.N.; McCarty, J.C.; Campbell, B.T.; Hayes, R.W.; Stelly, D.M. Tri-species shuffling of chromosomes to study the effects on fiber traits using chromosome substitution lines. Crop Sci. 2017, 57, 1211–1226. [Google Scholar] [CrossRef]
- Awasthi, A.; Reddy, K.R.; Saha, S.; Jenkins, J.N.; Stelly, D.M. Morph-physiological responses of cotton interspecific chromosome substitution lines to low temperature and drought stresses. Euphytica 2018, 214, 218. [Google Scholar] [CrossRef]
- Zhai, H.; Gong, W.; Tan, Y.; Liu, A.; Song, W.; Li, J.; Deng, Z.; Kong, L.; Gong, J.; Shang, H. Identification of chromosome segment substitution lines of Gossypium barbadense introgressed in G. hirsutum and quantitative trait locus mapping for fiber quality and yield traits. PLoS ONE 2016, 11, e0159101. [Google Scholar] [CrossRef]
- USDA-ERS. USDA ERS—Background [WWW Document]. Available online: https://www.ers.usda.gov/data-products/cotton-wool-and-textile-data/cotton-and-wool-yearbook/ (accessed on 24 August 2020).
- Singh, R.P.; Prasad, P.V.; Sunita, K.; Giri, S.; Reddy, K.R. Influence of high temperature and breeding for heat tolerance in cotton: A review. Adv. Agron. 2007, 93, 313–385. [Google Scholar]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed]
- Tuberosa, R. Phenotyping for drought tolerance of crops in the genomics era. Front. Physiol. 2012, 3, 347. [Google Scholar] [CrossRef] [PubMed]
- Tricker, P.J.; ElHabti, A.; Schmidt, J.; Fleury, D. The physiological and genetic basis of combined drought and heat tolerance in wheat. J. Exp. Bot. 2018, 69, 3195–3210. [Google Scholar] [CrossRef] [PubMed]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crop. Prod. 2019, 130, 118–129. [Google Scholar] [CrossRef]
- Saha, S.; Stelly, D.M.; Raska, D.A.; Wu, J.; Jenkins, J.N.; McCarty, J.C.; Makamov, A.; Gotmare, V.; Abdurakhmonov, I.Y.; Campbell, B. Chromosome substitution lines: Concept, development and utilization in the genetic improvement of Upland cotton. In Plant Breeding; IntechOpen: London, UK, 2012. [Google Scholar]
- Singh, K.; Wijewardana, C.; Gajanayake, B.; Lokhande, S.; Wallace, T.; Jones, D.; Reddy, K.R. Genotypic variability among cotton cultivars for heat and drought tolerance using reproductive and physiological traits. Euphytica 2018, 214, 57. [Google Scholar] [CrossRef]
- Kusumi, K.; Hirotsuka, S.; Kumamaru, T.; Iba, K. Increased leaf photosynthesis caused by elevated stomatal conductance in a rice mutant deficient in SLAC1, a guard cell anion channel protein. J. Exp. Bot. 2012, 63, 5635–5644. [Google Scholar] [CrossRef] [PubMed]
- Kondamudi, R.; Swamy, K.; Rao, Y.V.; Kiran, T.V.; Suman, K.; Rao, D.S.; Rao, P.R.; Subrahmanyam, D.; Sarla, N.; Kumari, B.R. Gas exchange, carbon balance and stomatal traits in wild and cultivated rice (Oryza sativa L.) genotypes. Acta Physiol. Plant. 2016, 38, 160. [Google Scholar] [CrossRef]
- Radin, J.W.; Lu, Z.; Percy, R.G.; Zeiger, E. Genetic variability for stomatal conductance in Pima cotton and its relation to improvements of heat adaptation. Proc. Natl. Acad. Sci. USA 1994, 91, 7217–7221. [Google Scholar] [CrossRef]
- Lu, Z.; Radin, J.W.; Turcotte, E.L.; Percy, R.; Zeiger, E. High yields in advanced lines of Pima cotton are associated with higher stomatal conductance, reduced leaf area and lower leaf temperature. Physiol. Plant. 1994, 92, 266–272. [Google Scholar] [CrossRef]
- Udayakumar, M.; Sheshshayee, M.; Nataraj, K. Why has breeding for water use efficiency not been successful? An analysis and alternate approach to exploit this trait for crop improvement. Curr. Sci. 1998, 74, 994–1000. [Google Scholar]
- Ma, B.; Morrison, M.; Voldeng, H. Leaf greenness and photosynthetic rates in soybean. Crop Sci. 1995, 35, 1411–1414. [Google Scholar] [CrossRef]
- Elfeky, S.S.; Osman, M.E.; Hamada, S.M.; Hasan, A.M. Effect of salinity and drought on growth criteria and biochemical analysis of Catharanthus roseus shoot. Int. J. Bot. 2007, 3, 202–207. [Google Scholar] [CrossRef]
- Wang, L.; Deng, F.; Ren, W.-J. Shading tolerance in rice is related to better light harvesting and use efficiency and grain filling rate during grain filling period. Field Crop. Res. 2015, 180, 54–62. [Google Scholar] [CrossRef]
- Kittock, D.; Turcotte, E.; Hofmann, W. Estimation of heat tolerance improvement in recent American pima cotton cultivars. J. Agron. Crop Sci. 1988, 161, 305–309. [Google Scholar] [CrossRef]
- Reddy, K.R.; Hodges, H.F.; McKinion, J.M. A comparison of scenarios for the effect of global climate change on cotton growth and yield. Funct. Plant Biol. 1997, 24, 707–713. [Google Scholar] [CrossRef]
- Mohan, M.; Narayanan, S.L.; Ibrahim, S. Chlorophyll stability index (CSI): Its impact on salt tolerance in rice. Int. Rice Res. Notes 2000, 25, 38–39. [Google Scholar]
- ur Rahman, H.; Malik, S.A.; Saleem, M. Heat tolerance of upland cotton during the fruiting stage evaluated using cellular membrane thermostability. Field Crop. Res. 2004, 85, 149–158. [Google Scholar] [CrossRef]
- Rebetzke, G.; Botwright, T.; Moore, C.; Richards, R.; Condon, A. Genotypic variation in specific leaf area for genetic improvement of early vigour in wheat. Field Crop. Res. 2004, 88, 179–189. [Google Scholar] [CrossRef]
- Kiran, T.; Rao, Y.; Subrahmanyam, D.; Rani, N.; Bhadana, V.; Rao, P.; Voleti, S. Variation in leaf photosynthetic characteristics in wild rice species. Photosynthetica 2013, 51, 350–358. [Google Scholar] [CrossRef]
- Nageswara Rao, R.; Talwar, H.; Wright, G. Rapid assessment of specific leaf area and leaf nitrogen in peanut (Arachis hypogaea L.) using a chlorophyll meter. J. Agron. Crop Sci. 2001, 186, 175–182. [Google Scholar] [CrossRef]
- Sheshshayee, M.; Bindumadhava, H.; Rachaputi, N.; Prasad, T.; Udayakumar, M.; Wright, G.; Nigam, S. Leaf chlorophyll concentration relates to transpiration efficiency in peanut. Ann. Appl. Biol. 2006, 148, 7–15. [Google Scholar] [CrossRef]
- Songsri, P.; Jogloy, S.; Holbrook, C.; Kesmala, T.; Vorasoot, N.; Akkasaeng, C.; Patanothai, A. Association of root, specific leaf area and SPAD chlorophyll meter reading to water use efficiency of peanut under different available soil water. Agric. Water Manag. 2009, 96, 790–798. [Google Scholar] [CrossRef]
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [Google Scholar] [CrossRef]
- Prasad, P.V.; Bheemanahalli, R.; Jagadish, S.K. Field crops and the fear of heat stress—opportunities, challenges and future directions. Field Crop. Res. 2017, 200, 114–121. [Google Scholar] [CrossRef]
- Burke, J.J.; Velten, J.; Oliver, M.J. In vitro analysis of cotton pollen germination. Agron. J. 2004, 96, 359–368. [Google Scholar] [CrossRef]
- Porch, T.; Jahn, M. Effects of high-temperature stress on microsporogenesis in heat-sensitive and heat-tolerant genotypes of Phaseolus vulgaris. Plant Cell Environ. 2001, 24, 723–731. [Google Scholar] [CrossRef]
- Prasad, P.V.; Craufurd, P.; Summerfield, R. Fruit number in relation to pollen production and viability in groundnut exposed to short episodes of heat stress. Ann. Bot. 1999, 84, 381–386. [Google Scholar] [CrossRef]
- Cross, R.; McKay, S.; McHughen, A.; Bonham-Smith, P. Heat-stress effects on reproduction and seed set in Linum usitatissimum L. (flax). Plant Cell Environ. 2003, 26, 1013–1020. [Google Scholar] [CrossRef]
- Chiluwal, A.; Bheemanahalli, R.; Kanaganahalli, V.; Boyle, D.; Perumal, R.; Pokharel, M.; Oumarou, H.; Jagadish, S.K. Deterioration of ovary plays a key role in heat stress-induced spikelet sterility in sorghum. Plant Cell Environ. 2020, 43, 448–462. [Google Scholar] [CrossRef]
- Peet, M.; Sato, S.; Gardner, R. Comparing heat stress effects on male-fertile and male-sterile tomatoes. Plant Cell Environ. 1998, 21, 225–231. [Google Scholar] [CrossRef]
- Prasad, P.V.; Boote, K.J.; Allen, L.H., Jr.; Thomas, J.M. Effects of elevated temperature and carbon dioxide on seed-set and yield of kidney bean (Phaseolus vulgaris L.). Glob. Chang. Biol. 2002, 8, 710–721. [Google Scholar] [CrossRef]
- Vara Prasad, P.; Boote, K.J.; Hartwell Allen, L., Jr.; Thomas, J.M. Super-optimal temperatures are detrimental to peanut (Arachis hypogaea L.) reproductive processes and yield at both ambient and elevated carbon dioxide. Glob. Chang. Biol. 2003, 9, 1775–1787. [Google Scholar] [CrossRef]
- Young, L.W.; Wilen, R.W.; Bonham-Smith, P.C. High temperature stress of Brassica napus during flowering reduces micro-and megagametophyte fertility, induces fruit abortion, and disrupts seed production. J. Exp. Bot. 2004, 55, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.A.; Kakani, V.G.; Koti, S.; Reddy, K.R. Pollen-based screening of soybean genotypes for high temperatures. Crop Sci. 2007, 47, 219–231. [Google Scholar] [CrossRef]
- Bheemanahalli, R.; Sunoj, V.J.; Saripalli, G.; Prasad, P.V.; Balyan, H.; Gupta, P.; Grant, N.; Gill, K.S.; Jagadish, S.K. Quantifying the impact of heat stress on pollen germination, seed set, and grain filling in spring wheat. Crop Sci. 2019, 59, 684–696. [Google Scholar] [CrossRef]
- Rodriguez-Garay, B.; Barrow, J.R. Pollen selection for heat tolerance in cotton. Crop Sci. 1988, 28, 857–859. [Google Scholar] [CrossRef]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef]
- Reddy, A.R.; Reddy, K.R.; Padjung, R.; Hodges, H.F. Nitrogen nutrition and photosynthesis in leaves of Pima cotton. J. Plant Nutr. 1996, 19, 755–770. [Google Scholar] [CrossRef]
- Reddy, K.R.; Hodges, H.F.; Kimball, B.A. Crop ecosystem responses to climatic change: Cotton. In Climate Change and Global Crop Productivity; Reddy, K.R., Hodges, H.F., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 161–187. [Google Scholar]
- Zhao, D.; Reddy, K.R.; Kakani, V.G.; Mohammed, A.R.; Read, J.J.; Gao, W. Leaf and canopy photosynthetic characteristics of cotton (Gossypium hirsutum) under elevated CO2 concentration and UV-B radiation. J. Plant Physiol. 2004, 161, 581–590. [Google Scholar] [CrossRef]
- Hulse-Kemp, A.M.; Lemm, J.; Plieske, J.; Ashrafi, H.; Buyyarapu, R.; Fang, D.D.; Frelichowski, J.; Giband, M.; Hague, S.; Hinze, L.L. Development of a 63K SNP array for cotton and high-density mapping of intraspecific and interspecific populations of Gossypium spp. G3 Genes Genomes Genet. 2015, 5, 1187–1209. [Google Scholar] [CrossRef]
- Saha, S.; Stelly, D.M.; Makamov, A.K.; Ayubov, M.S.; Raska, D.; Gutiérrez, O.A.; Manchali, S.; Jenkins, J.N.; Deng, D.; Abdurakhmonov, I.Y. Molecular confirmation of Gossypium hirsutum chromosome substitution lines. Euphytica 2015, 205, 459–473. [Google Scholar] [CrossRef]
- Hendrix, B.; Stewart, J.M. Estimation of the nuclear DNA content of Gossypium species. Ann. Bot. 2005, 95, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Sreedasyam, A.; Ando, A.; Song, Q.; De Santiago, L.M.; Hulse-Kemp, A.M.; Ding, M.; Ye, W.; Kirkbride, R.C.; Jenkins, J. Genomic diversifications of five Gossypium allopolyploid species and their impact on cotton improvement. Nat. Genet. 2020, 52, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Luo, L. Breeding for water-saving and drought-resistance rice (WDR) in China. J. Exp. Bot. 2010, 61, 3509–3517. [Google Scholar] [CrossRef]
- Manavalan, L.P.; Guttikonda, S.K.; Phan Tran, L.-S.; Nguyen, H.T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef]
- Gajanayake, B.; Trader, B.W.; Reddy, K.R.; Harkess, R.L. Screening ornamental pepper cultivars for temperature tolerance using pollen and physiological parameters. HortScience 2011, 46, 878–884. [Google Scholar] [CrossRef]
- Wijewardana, C.; Henry, W.B.; Reddy, K.R. Evaluation of drought tolerant maize germplasm to induced drought stress. J. Miss. Acad. Sci. 2017, 62, 316–329. [Google Scholar]
- Wagner, W.; Herbst, D.; Sohmer, S. Manual of the Flowering Plants of HAWAII; Two Volumes; Bernice Pauahi Bishop Museum Special Publication; University of Hawaii Press: Honolulu, HI, USA, 1990; Volume 83. [Google Scholar]
- Lee, J.A.; Fang, D.D. Cotton as a world crop: Origin, history, and current status. Cotton 2015, 57, 1–23. [Google Scholar]
- Morrow, M.; Krieg, D. Cotton management strategies for a short growing season environment: Water-nitrogen considerations. Agron. J. 1990, 82, 52–56. [Google Scholar] [CrossRef]
- Bowman, D.T.; Gutiérrez, O.A. Sources of fiber strength in the US upland cotton crop from 1980 to 2000. J. Cotton Sci. 2003, 7, 64–69. [Google Scholar]
- Loka, D.A.; Oosterhuis, D.M. Water Stress and Reproductive Development in Cotton. In Flowering and Fruiting in Cotton; Oosterhuis, D.M., Cothren, J.T., Eds.; Cotton Foundation: Memphis, TN, USA, 2012; pp. 51–58. [Google Scholar]
- Grimes, D.W.; Yamada, H.; Dickens, W. Functions for cotton (Gossypium hirsutum L.) production from irrigation and nitrogen fertilization variables: I. Yield and evapotranspiration. Agron. J. 1969, 61, 769–773. [Google Scholar] [CrossRef]
- Johnson, R.M.; Downer, R.G.; Bradow, J.M.; Bauer, P.J.; Sadler, E.J. Variability in cotton fiber yield, fiber quality, and soil properties in a southeastern coastal plain. Agron. J. 2002, 94, 1305–1316. [Google Scholar] [CrossRef]
- Lokhande, S.; Reddy, K.R. Reproductive and fiber quality responses of upland cotton to moisture deficiency. Agron. J. 2014, 106, 1060–1069. [Google Scholar] [CrossRef]
- Kakani, V.; Prasad, P.; Craufurd, P.; Wheeler, T. Response of in vitro pollen germination and pollen tube growth of groundnut (Arachis hypogaea L.) genotypes to temperature. Plant Cell Environ. 2002, 25, 1651–1661. [Google Scholar] [CrossRef]
- Kohel, R.J.; Yu, J.; Park, Y.-H.; Lazo, G.R. Molecular mapping and characterization of traits controlling fiber quality in cotton. Euphytica 2001, 121, 163–172. [Google Scholar] [CrossRef]
- Aslam, M.; Brown, M.; Kohel, R. Evaluation of seven tetrazolium salts as vital pollen stains in cotton Gossypium hirsutum L. 1. Crop Sci. 1964, 4, 508–510. [Google Scholar] [CrossRef]
- Taylor, R.M. Germination of cotton (Gossypium hirsutum L.) Pollen on an artificial medium 1. Crop Sci. 1972, 12, 243–244. [Google Scholar] [CrossRef]
- Chapple, C.; Vogt, T.; Ellis, B.E.; Somerville, C.R. An Arabidopsis mutant defective in the general phenylpropanoid pathway. Plant Cell 1992, 4, 1413–1424. [Google Scholar]



{kind=link}
{kind=link}
| CS Line | Pn | Gs | T | iWUE | Ci/Ca | TCHL | Caro | CTD | CMTS | CSI | SLA | PV | PG30 | PG38 | PGR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CS-B01 | 33.3 | 0.51 | 10.4 | 3.20 | 0.71 | 43.5 | 9.8 | 3.40 | 38.5 | 92.0 | 117.0 | 45.0 | 42.2 | 32.0 | 75.9 |
| CS-B04 | 38.1 | 0.70 | 12.5 | 3.10 | 0.73 | 44.3 | 9.9 | −1.70 | 48.1 | 88.0 | 131.0 | 50.0 | 47.2 | 37.5 | 79.1 |
| CS-B07 | 30.8 | 0.50 | 10.1 | 3.10 | 0.70 | 42.9 | 9.3 | 2.90 | 35.4 | 80.0 | 124.0 | 52.0 | 49.3 | 32.7 | 66.3 |
| CS-B15sh | 37.2 | 0.83 | 12.3 | 3.00 | 0.71 | 47.5 | 10.3 | 3.50 | 38.9 | 82.0 | 125.0 | 50.0 | 47.1 | 33.9 | 71.9 |
| CS-B18 | 34.0 | 0.56 | 10.8 | 3.20 | 0.71 | 49.8 | 11.0 | 1.40 | 35.7 | 87.0 | 127.0 | 50.0 | 46.5 | 37.2 | 79.8 |
| CS-T01 | 33.4 | 0.63 | 11.7 | 2.90 | 0.74 | 43.3 | 9.7 | 0.90 | 45.5 | 83.0 | 138.0 | 36.0 | 33.3 | 28.0 | 84.0 |
| CS-T04 | 32.8 | 0.47 | 9.5 | 3.50 | 0.68 | 45.9 | 10.1 | 5.00 | 39.5 | 85.0 | 124.0 | 33.0 | 29.6 | 27.0 | 91.3 |
| CS-T07 | 38.1 | 0.86 | 12.4 | 3.10 | 0.71 | 49.4 | 10.4 | 3.70 | 35.3 | 86.0 | 121.0 | 43.0 | 40.2 | 33.2 | 74.6 |
| CS-T08sh | 35.4 | 0.64 | 11.5 | 3.10 | 0.72 | 42.7 | 9.9 | −0.70 | 29.0 | 94.0 | 123.0 | 49.0 | 45.8 | 37.8 | 82.4 |
| CS-T15sh | 37.4 | 0.84 | 13.3 | 2.80 | 0.74 | 44.6 | 10.2 | −2.50 | 42.0 | 90.0 | 123.0 | 50.0 | 46.9 | 33.1 | 70.7 |
| CS-T18 | 37.8 | 0.76 | 11.8 | 3.20 | 0.67 | 47.4 | 10.2 | 1.90 | 46.8 | 83.0 | 132.0 | 38.0 | 34.6 | 30.4 | 88.4 |
| TM-1 | 33.1 | 0.63 | 10.7 | 3.10 | 0.70 | 45.2 | 9.9 | 4.30 | 39.8 | 94.0 | 129.0 | 55.0 | 52.4 | 33.1 | 63.3 |
| LSD (p = 0.05) | 2.9 | 0.1 | 1.7 | 0.28 | ns | 4.7 | ns | 3.3 | 3.8 | 4.4 | 6.9 | 6.5 | 6.52 | 5.79 | 9.7 |
| Significance | *** | *** | ** | ** | * | *** | *** | *** | *** | *** | *** | ** | *** | ||
| CS lines mean + | 35.3 | 0.66 | 11.5 | 3.11 | 0.71 | 45.6 | 10.1 | 1.62 | 39.5 | 86.4 | 125.9 | 45.1 | 42.1 | 33.0 | 78.6 |
| Advanced Upland cotton cultivars # | |||||||||||||||
| Minimum | 17 | 0.17 | 4.5 | 1.60 | 0.43 | 30.4 | 6.7 | 1.2 | 15.6 | 72 | 155.9 | 43.8 | 25.6 | 15.4 | 42.8 |
| Maximum | 38.6 | 0.85 | 14.4 | 4.40 | 0.84 | 38.1 | 10.2 | 4.3 | 40.4 | 92.9 | 199.6 | 66.3 | 49.9 | 41.8 | 94.4 |
| Mean | 29.3 | 0.51 | 10.1 | 3.20 | 0.67 | 34.0 | 8.00 | 2.9 | 26.1 | 82.1 | 184 | 60.2 | 40.1 | 27.7 | 68.9 |
| CS Line | Pn | Gs | T | iWUE | CTD | CMTS | CSI | SLA | PV | PG30 | PGR |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CS-B01 | −11.9 | −10.2 | −10.2 | +12.6 | |||||||
| CS-B04 | +4.9 | −6.0 | +8.3 | +15.8 | |||||||
| CS-B07 | −13.6 | ||||||||||
| CS-B15sh | +4.1 | +0.20 | −11.3 | +8.6 | |||||||
| CS-B18 | −7.2 | +16.5 | |||||||||
| CS-T01 | +5.7 | −11.1 | −19.0 | −19.0 | +20.6 | ||||||
| CS-T04 | +0.42 | −9.0 | −22.8 | −22.8 | +27.9 | ||||||
| CS-T07 | +4.9 | +0.23 | −7.6 | −12.1 | −12.1 | +11.3 | |||||
| CS-T08sh | −5.0 | −10.9 | +19.1 | ||||||||
| CS-T15sh | +4.2 | +0.21 | +2.60 | −6.8 | |||||||
| CS-T18 | +4.6 | +6.9 | −10.6 | −17.8 | −17.8 | +25.1 |
| Heat Tolerant | Drought Tolerant | Combined Heat and Drought Tolerant | ||||||
|---|---|---|---|---|---|---|---|---|
| Low | Moderate | High | Low | Moderate | High | Low | Moderate | High |
| CS-T01 (11.2) | TM-1 (12) | CS-B15sh (12.4) | CS-B04 (10.2) | CS-T04 (10.8) | CS-T07 (11.4) | CS-T01 (12.3) | CS-T04 (12.8) | CS-T07 (13.4) |
| CS-T15sh (11.1) | CS-T04 (11.9) | CS-T07 (12.4) | CS-T15sh (10.1) | CS-B01 (10.4) | CS-B15sh (11.3) | CS-T15sh (12.1) | CS-B18 (12.5) | CS-B15sh (13.4) |
| CS-B07 (11) | CS-B01 (11.5) | CS-T18 (12.1) | CS-B07 (9.9) | CS-B18 (10.3) | CS-T18 (11.1) | CS-B07 (11.9) | CS-B01 (12.4) | CS-T18 (13.1) |
| CS-T08sh (10.9) | CS-B18 (11.5) | CS-T08sh (9.6) | CS-T01 (10.3) | TM-1 (11) | CS-T08sh (11.8) | CS-B04 (12.4) | TM-1 (13) | |
| CS-B04 (11.4) | ||||||||
| CS Line | SSR Markers * | Non Target (>5 Mb) ** | Non Target (<5 Mb) *** | Target **** |
|---|---|---|---|---|
| CS-B01 | BNL2921-158, BNL3580-207, BNL3848-197, BNL-3888-197, CIR009-233, NAU2437-235 | c06, c25 | c09, c25 | Substituted |
| CS-B04 | BNL2821 = 192, BNL3089-124, BNL3988-120, CIR222-276, CIR249-190 | - | c18; c20 | Substituted |
| CS-B07 | - | c11, c16, | c04, c16, c20, c21, c22 | TM-1 |
| CS-B15sh | - | c07 | c02, c10; c13, c20 | TM-1 |
| CS-B18 | BNL0193-111, BNL3479-251, CIR216-141, Gh501-201, TMB2762-205, | - | c12, c16, c17, c19, c24, c25, c26 | Substituted |
| CS-T01 | BNL1693-244, BNL2921-162, BNL3580 = 202, BNL3848-223, BNL3888-182, BNL2437-247 | c25 | c03, c07, c10, c20, c25 | Substituted |
| CS-T04 | - | - | c18 | Substituted |
| CS-T07 | BNL1395-163, BNL1597-228, CIR141-170, CIR169-135, Gh548-113, | c02, c21 | c05, c08 | Substituted |
| CS-T08sh | - | c12, c20 | c05, c06, c09, c13, c21, c25 | Substituted |
| CS-T15sh | - | c09, c12 | c08, c14, c17 | Substituted |
| CS-T18 | BNL0193-113, BNL1721-171, BNL243-135, BNL3479-244, BNL569-142, CIR216-141, CIR216-141 | c01 | c26 | Substituted |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reddy, K.R.; Bheemanahalli, R.; Saha, S.; Singh, K.; Lokhande, S.B.; Gajanayake, B.; Read, J.J.; Jenkins, J.N.; Raska, D.A.; Santiago, L.M.D.; et al. High-Temperature and Drought-Resilience Traits among Interspecific Chromosome Substitution Lines for Genetic Improvement of Upland Cotton. Plants 2020, 9, 1747. https://doi.org/10.3390/plants9121747
Reddy KR, Bheemanahalli R, Saha S, Singh K, Lokhande SB, Gajanayake B, Read JJ, Jenkins JN, Raska DA, Santiago LMD, et al. High-Temperature and Drought-Resilience Traits among Interspecific Chromosome Substitution Lines for Genetic Improvement of Upland Cotton. Plants. 2020; 9(12):1747. https://doi.org/10.3390/plants9121747
Chicago/Turabian StyleReddy, Kambham Raja, Raju Bheemanahalli, Sukumar Saha, Kulvir Singh, Suresh B. Lokhande, Bandara Gajanayake, John J. Read, Johnie N. Jenkins, Dwaine A. Raska, Luis M. De Santiago, and et al. 2020. "High-Temperature and Drought-Resilience Traits among Interspecific Chromosome Substitution Lines for Genetic Improvement of Upland Cotton" Plants 9, no. 12: 1747. https://doi.org/10.3390/plants9121747
APA StyleReddy, K. R., Bheemanahalli, R., Saha, S., Singh, K., Lokhande, S. B., Gajanayake, B., Read, J. J., Jenkins, J. N., Raska, D. A., Santiago, L. M. D., Hulse-Kemp, A. M., Vaughn, R. N., & Stelly, D. M. (2020). High-Temperature and Drought-Resilience Traits among Interspecific Chromosome Substitution Lines for Genetic Improvement of Upland Cotton. Plants, 9(12), 1747. https://doi.org/10.3390/plants9121747