Effect of Gibberellic Acid on Growing-Point Development of Non-Vernalized Wheat Plants under Long-Day Conditions

 ,
,  , ,
, ,
Abstract
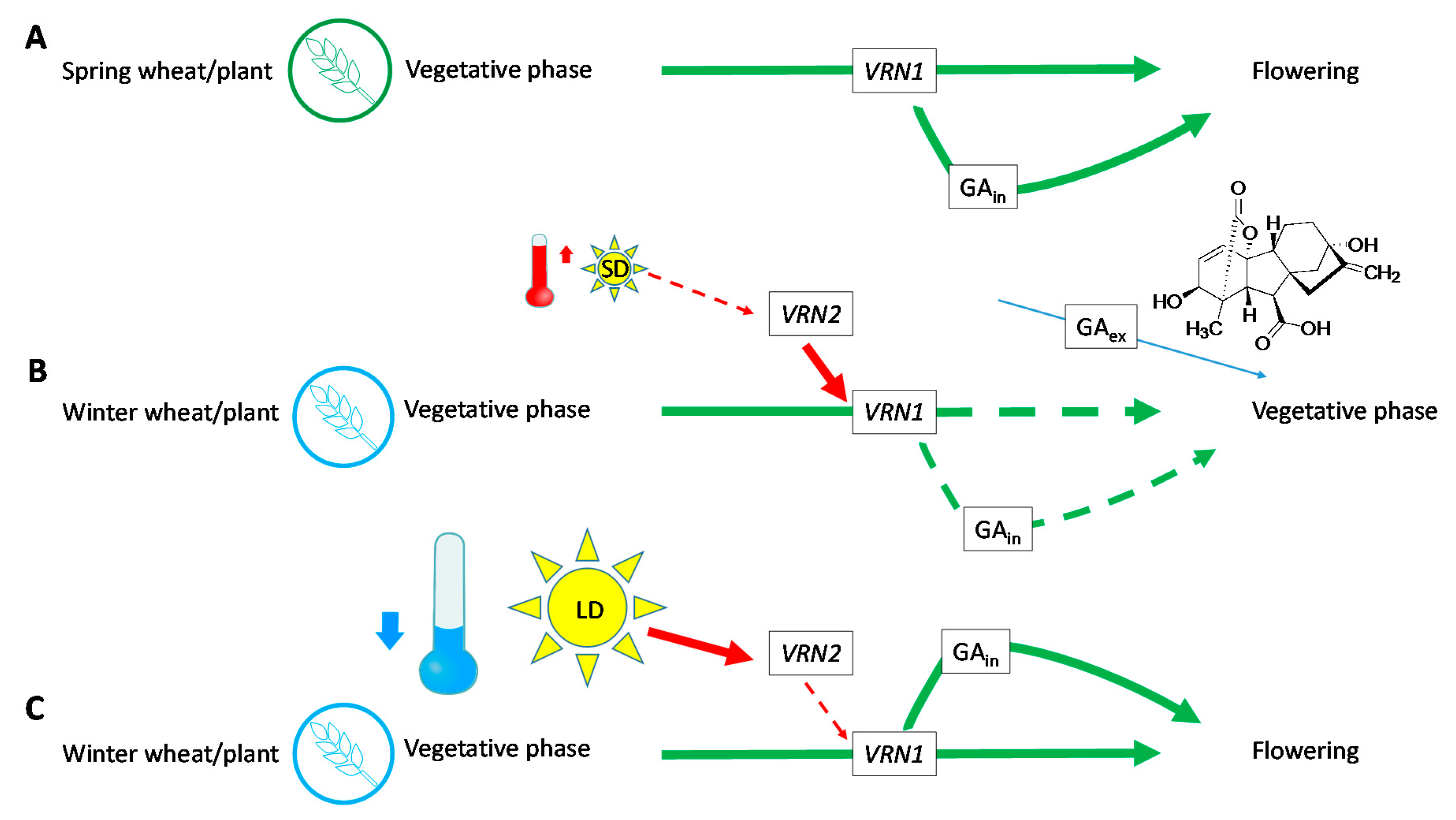
1. Introduction
2. Results and Discussion
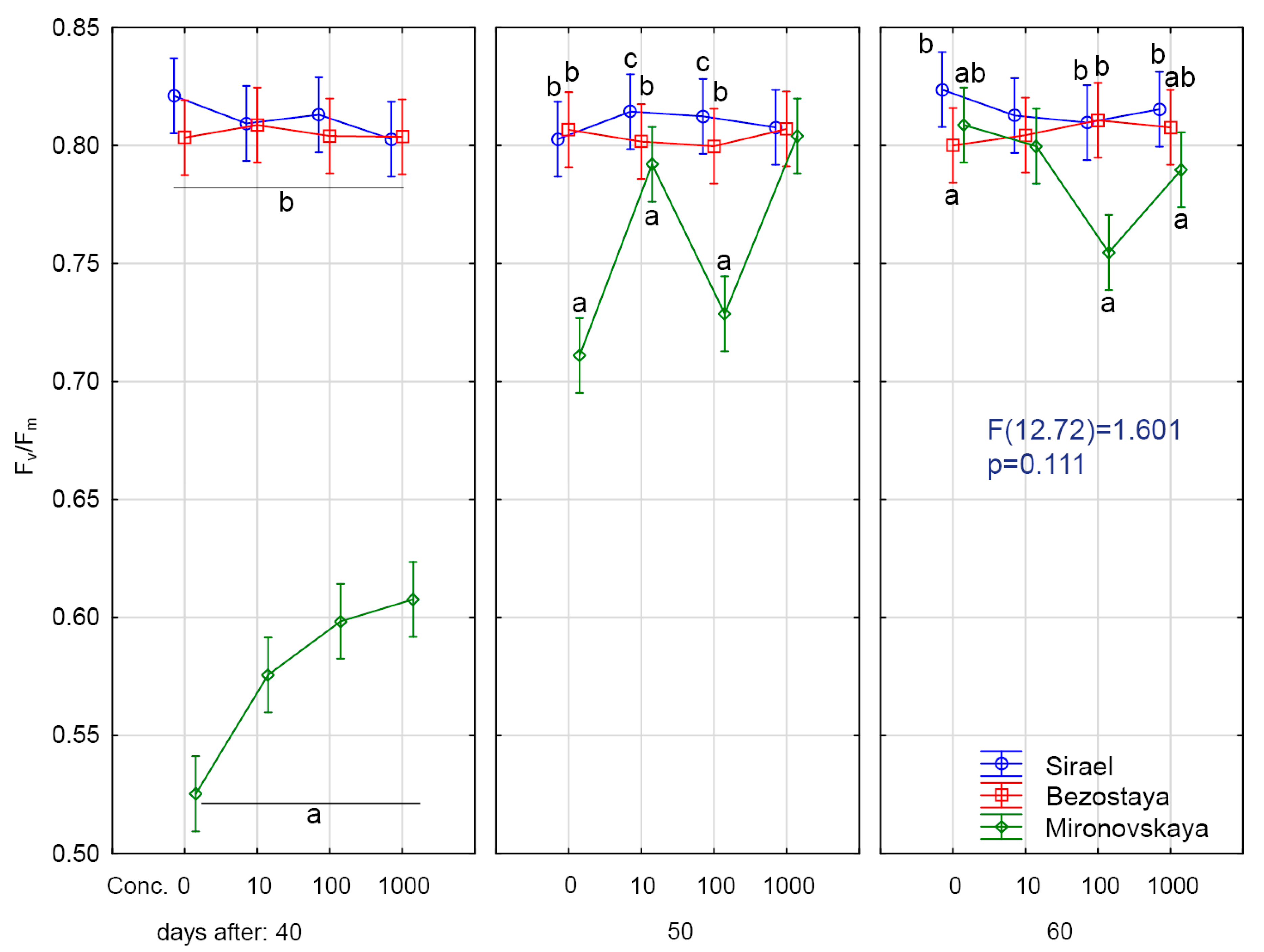
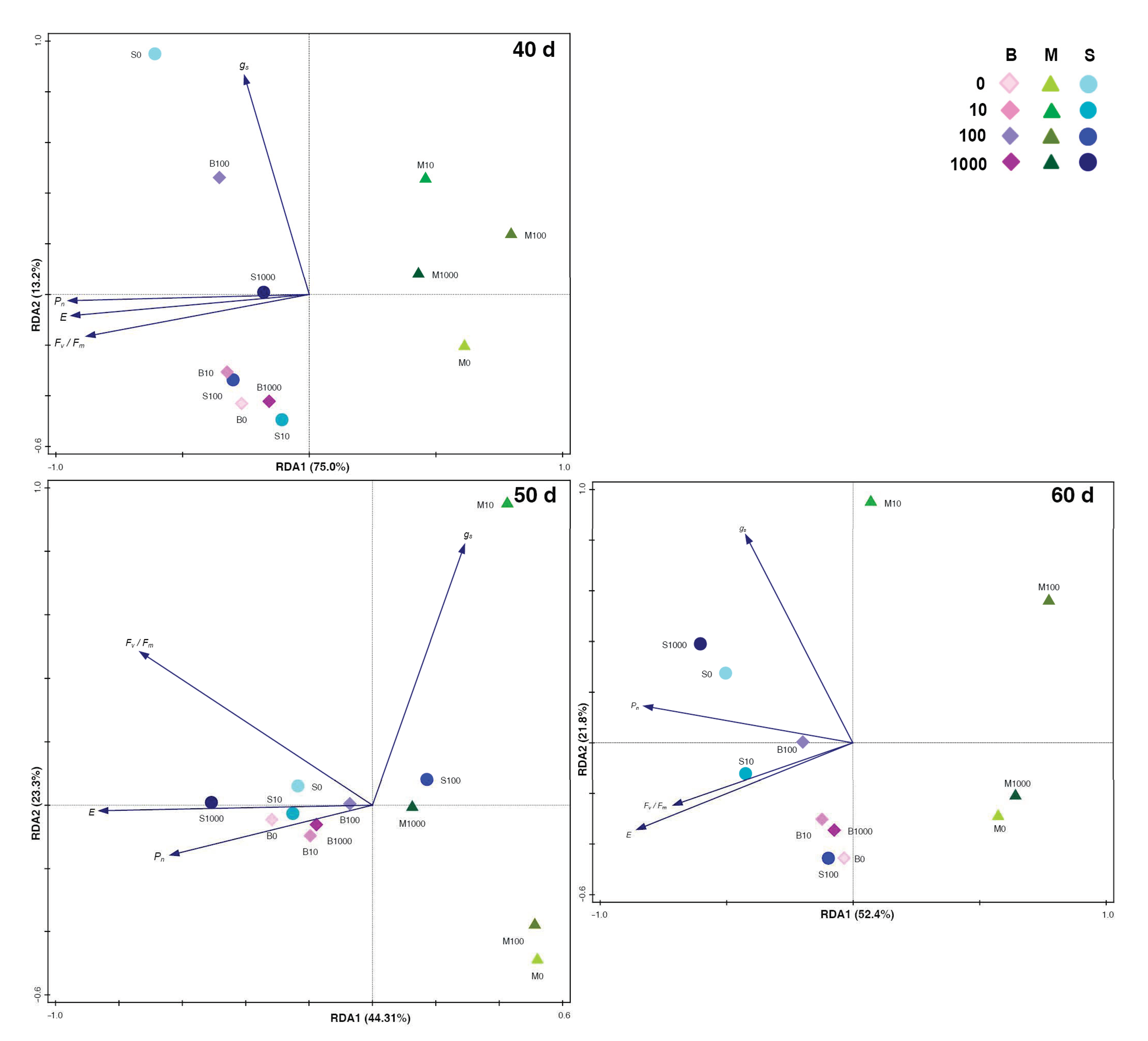
2.1. Results at Forty Days of Growth
2.2. Results at Fifty Days of Growth
2.3. Results at Sixty Days of Growth
2.4. Development of the Growing Point
3. Materials and Methods
3.1. Plant Material
3.2. Gibberellin GA3 Treatment of Seeds
3.3. Growing Conditions
3.4. Photosynthesis Measurement
3.5. Identifying the Microphenological Stage of the Growing Point
3.6. Statistical Analysis
4. Conclusions
Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cockram, J.; Jones, H.; Leigh, F.J.; O’Sullivan, D.; Powell, W.; Laurie, D.A.; Greenland, A.J. Control of flowering time in temperate cereals: Genes, domestication, and sustainable productivity. J. Exp. Bot. 2007, 58, 1231–1244. [Google Scholar] [CrossRef] [PubMed]
- Kippes, N.; Kippes, N.; Vangessel, C.; Hamilton, J.; Akpinar, A.; Budak, H.; Dubcovsky, J.; Dubcovsky, J.; Pearce, S. Effect of phyB and phyC loss-of-function mutations on the wheat transcriptome under short and long day photoperiods. BMC Plant Biol. 2020, 20, 297. [Google Scholar] [CrossRef] [PubMed]
- Royo, C.; Dreisigacker, S.; Soriano, J.M.; Lopes, M.S.; Ammar, K.; Villegas, D. Allelic Variation at the Vernalization Response (Vrn-1) and Photoperiod Sensitivity (Ppd-1) Genes and Their Association With the Development of Durum Wheat Landraces and Modern Cultivars. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Chen, A.; Li, C.; Hu, W.; Lau, M.Y.; Lin, H.; Rockwell, N.C.; Martin, S.S.; Jernstedt, J.A.; Lagarias, J.C.; Dubcovsky, J. PHYTOCHROME C plays a major role in the acceleration of wheat flowering under long-day photoperiod. Proc. Nat. Acad. Sci. USA 2014, 111, 10037–10044. [Google Scholar] [CrossRef]
- Pearce, S.; Vanzetti, L.S.; Dubcovsky, J. Exogenous gibberellins induce wheat spike development under short days only in the presence of VERNALIZATION1. Plant Physiol. 2013, 163, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.; Beales, J.; Faure, S.; Dunford, R.P.; Laurie, D.A. Botany: The pseudo-response regulator Ppd-H1 provides adaptation to photoperiod in barley. Science 2005, 310, 1031–1034. [Google Scholar] [CrossRef]
- Yunze, S.; Shuangsheng, G. Effects of photoperiod on wheat growth, development and yield in CELSS. Acta Astronaut. 2014, 105, 24–29. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Loukoianov, A.; Fu, D.; Valarik, M.; Sanchez, A.; Yan, L. Effect of photoperiod on the regulation of wheat vernalization genes VRN1 and VRN2. Plant Mol. Biol. 2006, 60, 469–480. [Google Scholar] [CrossRef]
- Pugsley, A.T. A genetic analysis of the spring-winter habit of growth in wheat. Aust. J. Agric. Res. 1971, 22, 21–31. [Google Scholar] [CrossRef]
- Chen, Y.; Sidhu, H.S.; Kaviani, M.; McElroy, M.S.; Pozniak, C.J.; Navabi, A. Application of image-based phenotyping tools to identify QTL for in-field winter survival of winter wheat (Triticum aestivum L.). Theor. Appl. Genet. 2019, 132, 2591–2604. [Google Scholar] [CrossRef]
- Sung, S.; Amasino, R.M. Remembering winter: Toward a molecular understanding of vernalization. Annu. Rev. Plant Biol. 2005, 56, 491–508. [Google Scholar] [CrossRef] [PubMed]
- Christy, B.; Riffkin, P.; Richards, R.; Partington, D.; Acuña, T.B.; Merry, A.; Zhang, H.; Trevaskis, B.; O’Leary, G. An allelic based phenological model to predict phasic development of wheat (Triticum aestivum L.). F. Crop. Res. 2020, 249. [Google Scholar] [CrossRef]
- Marcus, A.; Feeley, J.; Volcani, T. Protein Synthesis in Imbibed Seeds III. Kinetics of Amino Acid Incorporation Ribosome Activation, and Polysome Formation. Plant Physiol. 1966, 41, 1167–1172. [Google Scholar] [CrossRef]
- Chen, D.; Osborne, D.J. Ribosomal Genes and DNA Replication in Germinating Wheat Embryos. Nature 1970, 225, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P. Gibberellin metabolism and its regulation. J. Plant Growth Regul. 2001, 20, 317–318. [Google Scholar] [CrossRef]
- Swain, S.M.; Singh, D.P. Tall tales from sly dwarves: Novel functions of gibberellins in plant development. Trends Plant Sci. 2005, 10, 123–129. [Google Scholar] [CrossRef]
- Dubert, F.; Marcińska, I.; Biesaga-Kościelniak, J.; Szmider, I. The Effectiveness of Vernalization of Immature Embryos of Winter Wheat var. Grana as Related to Age and Exogenous Phytohormones. J. Agron. Crop Sci. 1993, 170, 234–242. [Google Scholar] [CrossRef]
- King, R.W.; Evans, L.T. Gibberellins and Flowering of Grasses and Cereals: Prizing Open the Lid of the “Florigen” Black Box. Annu. Rev. Plant Biol. 2003, 54, 307–328. [Google Scholar] [CrossRef]
- Lee, I.J.; Foster, K.R.; Morgan, P.W. Effect of gibberellin biosynthesis inhibitors on native gibberellin content, growth and floral initiation in Sorghum bicolor. J. Plant Growth Regul. 1998, 17, 185–195. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Sayed, O.H. Chlorophyll fluorescence as a tool in cereal crop research. Photosynthetica 2003, 41, 321–330. [Google Scholar] [CrossRef]
- Demmig, B.; Björkman, O. Comparison of the effect of excessive light on chlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higher plants. Planta 1987, 171, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.L.; Jing, R.L.; Chang, X.P.; Li, W. Quantitative trait loci mapping for chlorophyll fluorescence and associated traits in Wheat (Triticum aestivum). J. Integr. Plant Biol. 2007, 49, 646–654. [Google Scholar] [CrossRef]
- Liang, Y.; Zhang, K.; Zhao, L.; Liu, B.; Meng, Q.; Tian, J.; Zhao, S. Identification of chromosome regions conferring dry matter accumulation and photosynthesis in wheat (Triticum aestivum L.). Euphytica 2010, 171, 145–156. [Google Scholar] [CrossRef]
- McMaster, G.S.; White, J.W.; Hunt, L.A.; Jamieson, P.D.; Dhillon, S.S.; Ortiz-Monasterio, J.I. Simulating the influence of vernalization, photoperiod and optimum temperature on wheat developmental rates. Ann. Bot. 2008, 102, 561–569. [Google Scholar] [CrossRef]
- Čaňová, I.; Ďurkovič, J.; Hladká, D.; Lukáčik, I. Changes in stomatal characteristics and photochemical efficiency during leaf development in six species of Sorbus. Photosynthetica 2012, 50, 635–640. [Google Scholar] [CrossRef]
- Nakashima, T.; Araki, T.; Ueno, O. Photoprotective function of betacyanin in leaves of Amaranthus cruentus L. under water stress. Photosynthetica 2011, 49, 497–506. [Google Scholar] [CrossRef]
- Weng, J.H.; Chien, C.T.; Chen, C.W.; Lai, X.M. Effects of osmotic- and high-light stresses on PSII efficiency of attached and detached leaves of three tree species adapted to different water regimes. Photosynthetica 2011, 49, 555–563. [Google Scholar] [CrossRef]
- Manetas, Y.; Buschmann, C. The interplay of anthocyanin biosynthesis and chlorophyll catabolism in senescing leaves and the question of photosystem II photoprotection. Photosynthetica 2011, 49, 515–522. [Google Scholar] [CrossRef]
- Li, X.T.; Cao, P.; Wang, X.G.; Cao, M.J.; Yu, H.Q. Comparison of gas exchange and chlorophyll fluorescence of low-potassium-tolerant and -sensitive soybean [Glycine max (L.) Merr.] cultivars under low-potassium condition. Photosynthetica 2011, 49, 633–636. [Google Scholar] [CrossRef]
- Fracheboud, Y.; Jompuk, C.; Ribaut, J.M.; Stamp, P.; Leipner, J. Genetic analysis of cold-tolerance of photosynthesis in maize. Plant Mol. Biol. 2004, 56, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Zwar, J.A.; Chandler, P.M. α-Amylase production and leaf protein synthesis in a gibberellin-responsive dwarf mutant of “Himalaya” barley (Hordeum vulgare L.). Planta 1995, 197, 39–48. [Google Scholar] [CrossRef]
- Wang, L.L.; Chen, X.Y.; Yang, Y.; Wang, Z.; Xiong, F. Effects of exogenous gibberellic acid and abscisic acid on germination, amylases, and endosperm structure of germinating wheat seeds. Seed Sci. Technol. 2016, 44, 64–76. [Google Scholar] [CrossRef]
- Pavlista, A.D.; Baltensperger, D.D.; Santra, D.K.; Hergert, G.W.; Knox, S. Gibberellic Acid Promotes Early Growth of Winter Wheat and Rye. Am. J. Plant Sci. 2014, 05, 2984–2996. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef]
- Ghobadi, M.; Shafiei Abnavi, M.; Honarmand, S.J.; Ghobadi, M.E.; Reza Mohammadi, G. Effect of Hormonal Priming (GA3) and Osmopriming on Behavior of Seed Germination in Wheat (Triticum aestivum L.). J. Agric. Sci. 2012, 4. [Google Scholar] [CrossRef]
- Ritchie, G. A Chlorophyll Fluorescence: What Is It and What Do the Numbers Mean? USDA For. Serv. Proc. 2006, 34–43. [Google Scholar]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.O.; Rosenqvist, E. Wheat cultivars selected for high Fv/Fm under heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter. Physiol. Plant. 2015, 153, 284–298. [Google Scholar] [CrossRef]
- Xu, S.; Chong, K. Remembering winter through vernalisation. Nat. Plants 2018, 4, 997–1009. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Aziz, U.; Sahli, A.A.; Alyemeni, M.N.; Ahmad, P. Combined kinetin and spermidine treatments ameliorate growth and photosynthetic inhibition in vigna angularis by up-regulating antioxidant and nitrogen metabolism under cadmium stress. Biomolecules 2020. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Karim, F.; Rasul, E. Interactive effects of gibberellic acid (GA3) and salt stress on growth, ion accumulation and photosynthetic capacity of two spring wheat (Triticum aestivum L.) cultivars differing in salt tolerance. Plant Growth Regul. 2002, 36, 49–59. [Google Scholar] [CrossRef]
- Holá, D.; Benešová, M.; Honnerová, J.; Hnilička, F.; Rothová, O.; Kočová, M.; Hniličková, H. The evaluation of photosynthetic parameters in maize inbred lines subjected to water deficiency: Can these parameters be used for the prediction of performance of hybrid progeny? Photosynthetica 2010, 48, 545–558. [Google Scholar] [CrossRef]
- Pintó-Marijuan, M.; Munné-Bosch, S. Photo-oxidative stress markers as a measure of abiotic stress-induced leaf senescence: Advantages and limitations. J. Exp. Bot. 2014, 65, 3845–3857. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M.; Kalaji, H.M.; Carpentier, R.; Allakhverdiev, S.I. Photosystem II thermostability in situ: Environmentally induced acclimation and genotype-specific reactions in Triticum aestivum L. Plant Physiol. Biochem. 2012, 57, 93–105. [Google Scholar] [CrossRef]
- Stapleton, J.; Jones, M.B. Effects of vernalization on the subsequent rates of leaf extension and photosynthesis of perennial ryegrass (Lolium perenne L.). Grass Forage Sci. 1987, 42, 27–31. [Google Scholar] [CrossRef]
- Gardner, J.S.; Hess, W.M.; Trione, E.J. Development of the Young Wheat Spike: A Sem Study of Chinese Spring Wheat. Am. J. Bot. 1985, 72, 548. [Google Scholar] [CrossRef]
- Limin, A.; Corey, A.; Hayes, P.; Fowler, D.B. Low-temperature acclimation of barley cultivars used as parents in mapping populations: Response to photoperiod, vernalization and phenological development. Planta 2007, 226, 139–146. [Google Scholar] [CrossRef]
- Chen, L.; Hao, L.; Condon, A.G.; Hu, Y.G. Exogenous GA3 application can compensate the morphogenetic effects of the GA-responsive dwarfing gene Rht12 in bread wheat. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Mosaad, M.G.; Ortiz-Ferrara, G.; Mahalakshmi, V.; Fischer, R.A. Phyllochron response to vernalization and photoperiod in spring wheat. Crop. Sci. 1995, 35, 168–171. [Google Scholar] [CrossRef]
- Flood, R.G.; Halloran, G.M. Genetics and Physiology of Vernalization Response in Wheat. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 1986; Volume 39, pp. 87–125. [Google Scholar]
- Deng, W.; Casao, M.C.; Wang, P.; Sato, K.; Hayes, P.M.; Finnegan, E.J.; Trevaskis, B. Direct links between the vernalization response and other key traits of cereal crops. Nat. Commun. 2015. [Google Scholar] [CrossRef] [PubMed]
- Košner, J.; Pánková, K. The effect of the homoeologous group 5 chromosomes with different Vrn loci on growth phases and quantitative characters of wheat. Euphytica 2001, 119, 289–299. [Google Scholar] [CrossRef]
- González, F.G.; Slafer, G.A.; Miralles, D.J. Vernalization and photoperiod responses in wheat pre-flowering reproductive phases. F. Crop. Res. 2002, 74, 183–195. [Google Scholar] [CrossRef]
- Whitechurch, E.M.; Slafer, G.A.; Miralles, D.J. Variability in the duration of stem elongation in wheat genotypes and sensitivity to photoperiod and vernalization. J. Agron. Crop Sci. 2007, 193, 131–137. [Google Scholar] [CrossRef]
- Evans, L. Short Day Induction of Inflorescence Initiation in Some Winter Wheat Varieties. Funct. Plant Biol. 1987, 14, 277. [Google Scholar] [CrossRef]
- Koller, D.; Highkin, H.R.; Caso, O.H. Effects of Gibberellic Acid on Stem Apices of Vernalizable Grasses. Am. J. Bot. 1960. [Google Scholar] [CrossRef]
- Košner, J.; Pánková, K. The detection of allelic variants at the recessive vrn loci of winter wheat. Euphytica 1998, 101, 9–16. [Google Scholar] [CrossRef]
- Pharis, R.P.; Evans, L.T.; King, R.W.; Mander, L.N. Gibberellins, Endogenous and Applied, in Relation to Flower Induction in the Long-Day Plant Lolium temulentum. Plant Physiol. 1987, 84, 1132–1138. [Google Scholar] [CrossRef]
- Yan, L.; Loukoianov, A.; Blechl, A.; Tranquilli, G.; Ramakrishna, W.; SanMiguel, P.; Bennetzen, J.L.; Echenique, V.; Dubcovsky, J. The Wheat VRN2 Gene Is a Flowering Repressor Down-Regulated by Vernalization. Science 2004. [Google Scholar] [CrossRef]
- Jokela, V.; Virkajärvi, P.; Tanskanen, J.; Seppänen, M.M. Vernalization, gibberellic acid and photo period are important signals of yield formation in timothy (Phleum pratense). Physiol. Plant. 2014. [Google Scholar] [CrossRef]
- MacMillan, C.P.; Blundell, C.A.; King, R.W. Flowering of the Grass Lolium perenne. Effects of Vernalization and Long Days on Gibberellin Biosynthesis and Signaling. Plant Physiol. 2005, 138, 1794–1806. [Google Scholar] [CrossRef] [PubMed]
- Milec, Z.; Tomková, L.; Sumíková, T.; Pánková, K. A new multiplex PCR test for the determination of Vrn-B1 alleles in bread wheat (Triticum aestivum L.). Mol. Breed. 2012, 30, 317–323. [Google Scholar] [CrossRef]
- Wahid, A.; Perveen, M.; Gelani, S.; Basra, S.M.A. Pretreatment of seed with H2O2 improves salt tolerance of wheat seedlings by alleviation of oxidative damage and expression of stress proteins. J. Plant Physiol. 2007, 164, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Al-Khassawneh, N.M.; Karam, N.S.; Shibli, R.A. Growth and flowering of black iris (Iris nigricans Dinsm.) following treatment with plant growth regulators. Sci. Hortic. 2006, 107, 187–193. [Google Scholar] [CrossRef]
- Waddington, S.R.; Cartwright, P.M.; Wall, P.C. A quantitative scale of spike initial and pistil development in barley and wheat. Ann. Bot. 1983, 51, 119–130. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Bosa, K.; Kościelniak, J.; Żuk-Gołaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Smilauer, P.; Leps, J. Multivariate Analysis of Ecological Data Using Canoco 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; ISBN 9781139627061. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days After | 40 | 50 | 60 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variety/Conc. | 0 | 10 | 100 | 1000 | 0 | 10 | 100 | 1000 | 0 | 10 | 100 | 1000 |
| Mironovskaya | EE | EE | EE | EE | SR | SR | SR | SR | PSR | PRS | PSR | PSR |
| Bezostaya | EE | EE | EE | EE | SR | SR | SR | SR | PSR | PSR | PSR | PSR |
| Sirael | 2 ND | 2 ND | 2 ND | 2 ND | SE | SE | SE | SE | H | H | H | H |
| Time (16 h day/8 h night) | Temperature °C | Relative Humidity Φ (%) | Photosynthetic Photon Flux Density (PPDF) µmol m−2 s−1 |
|---|---|---|---|
| Dawn (5:00–6:00) | 17 | 65 | 280 |
| Day (6:00–20:00) | 20 | 60 | 560 |
| Nightfall (20:00–21:00) | 17 | 65 | 280 |
| Night (21:00–5:00) | 15 | 70 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skalicky, M.; Kubes, J.; Vachova, P.; Hajihashemi, S.; Martinkova, J.; Hejnak, V. Effect of Gibberellic Acid on Growing-Point Development of Non-Vernalized Wheat Plants under Long-Day Conditions. Plants 2020, 9, 1735. https://doi.org/10.3390/plants9121735
Skalicky M, Kubes J, Vachova P, Hajihashemi S, Martinkova J, Hejnak V. Effect of Gibberellic Acid on Growing-Point Development of Non-Vernalized Wheat Plants under Long-Day Conditions. Plants. 2020; 9(12):1735. https://doi.org/10.3390/plants9121735
Chicago/Turabian StyleSkalicky, Milan, Jan Kubes, Pavla Vachova, Shokoofeh Hajihashemi, Jaroslava Martinkova, and Vaclav Hejnak. 2020. "Effect of Gibberellic Acid on Growing-Point Development of Non-Vernalized Wheat Plants under Long-Day Conditions" Plants 9, no. 12: 1735. https://doi.org/10.3390/plants9121735
APA StyleSkalicky, M., Kubes, J., Vachova, P., Hajihashemi, S., Martinkova, J., & Hejnak, V. (2020). Effect of Gibberellic Acid on Growing-Point Development of Non-Vernalized Wheat Plants under Long-Day Conditions. Plants, 9(12), 1735. https://doi.org/10.3390/plants9121735