Roots Structure and Development of Austrobaileya scandens (Austrobaileyaceae) and Implications for Their Evolution in Angiosperms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
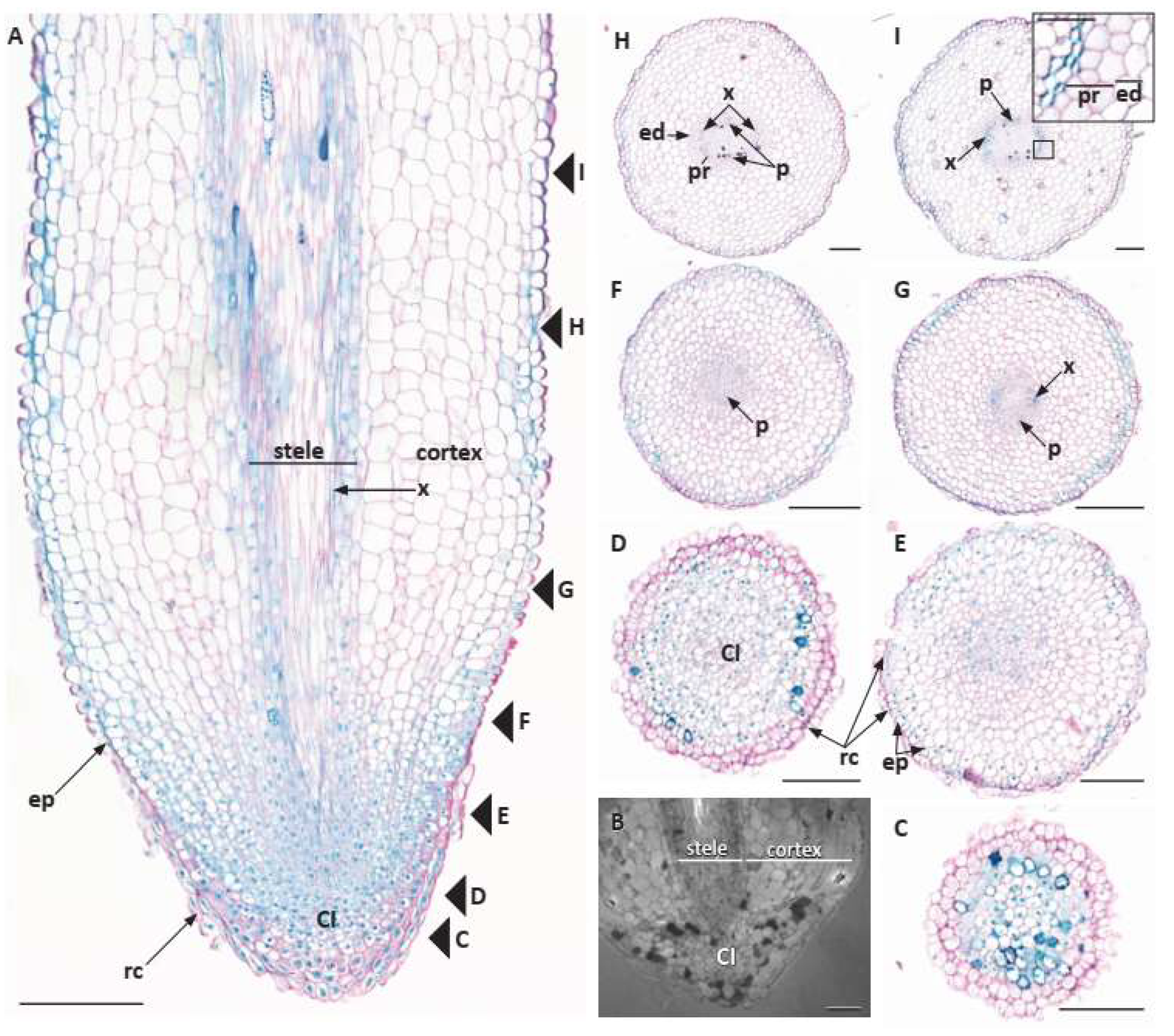
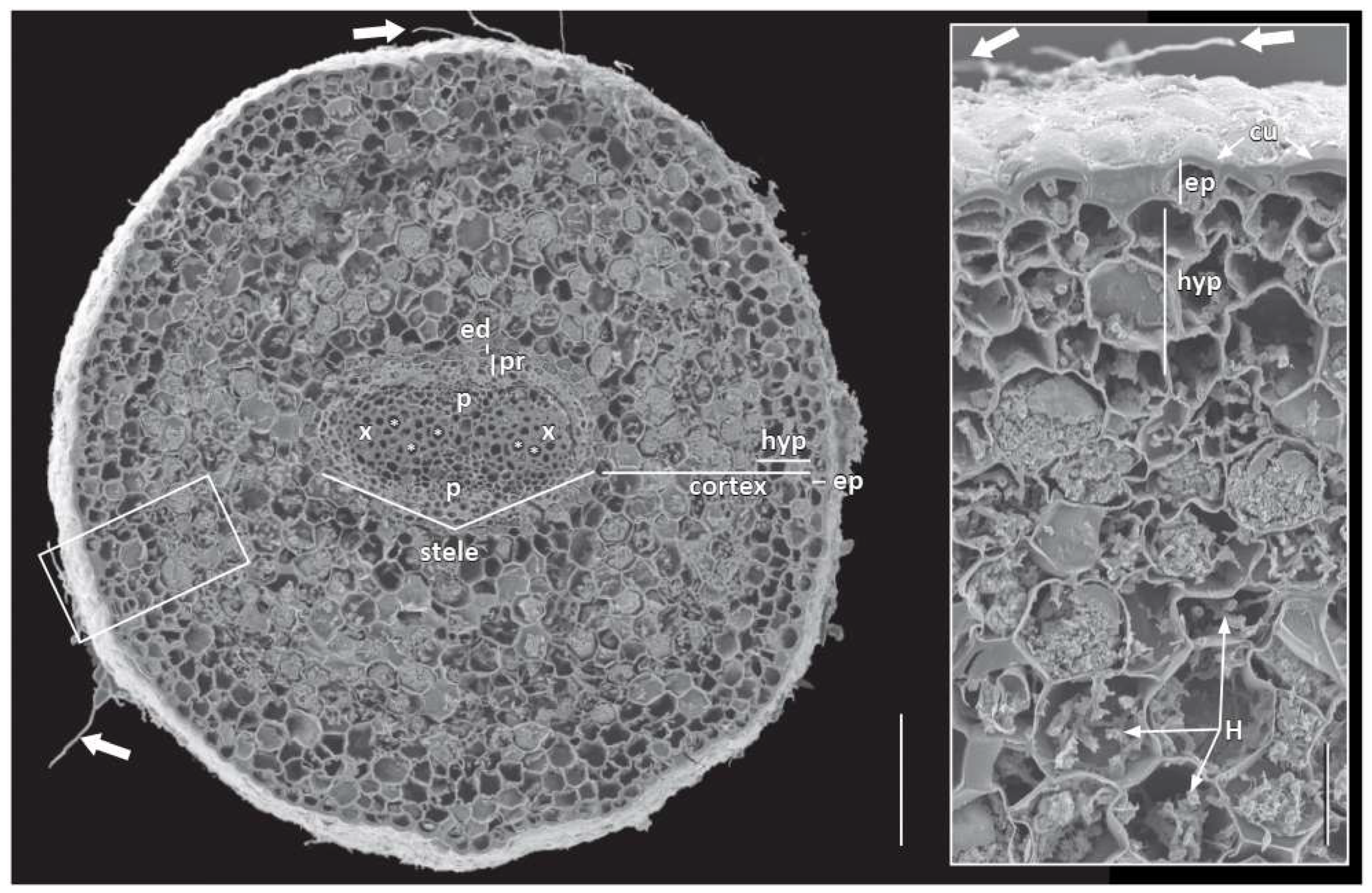
3.1. Root System and Apex Structure
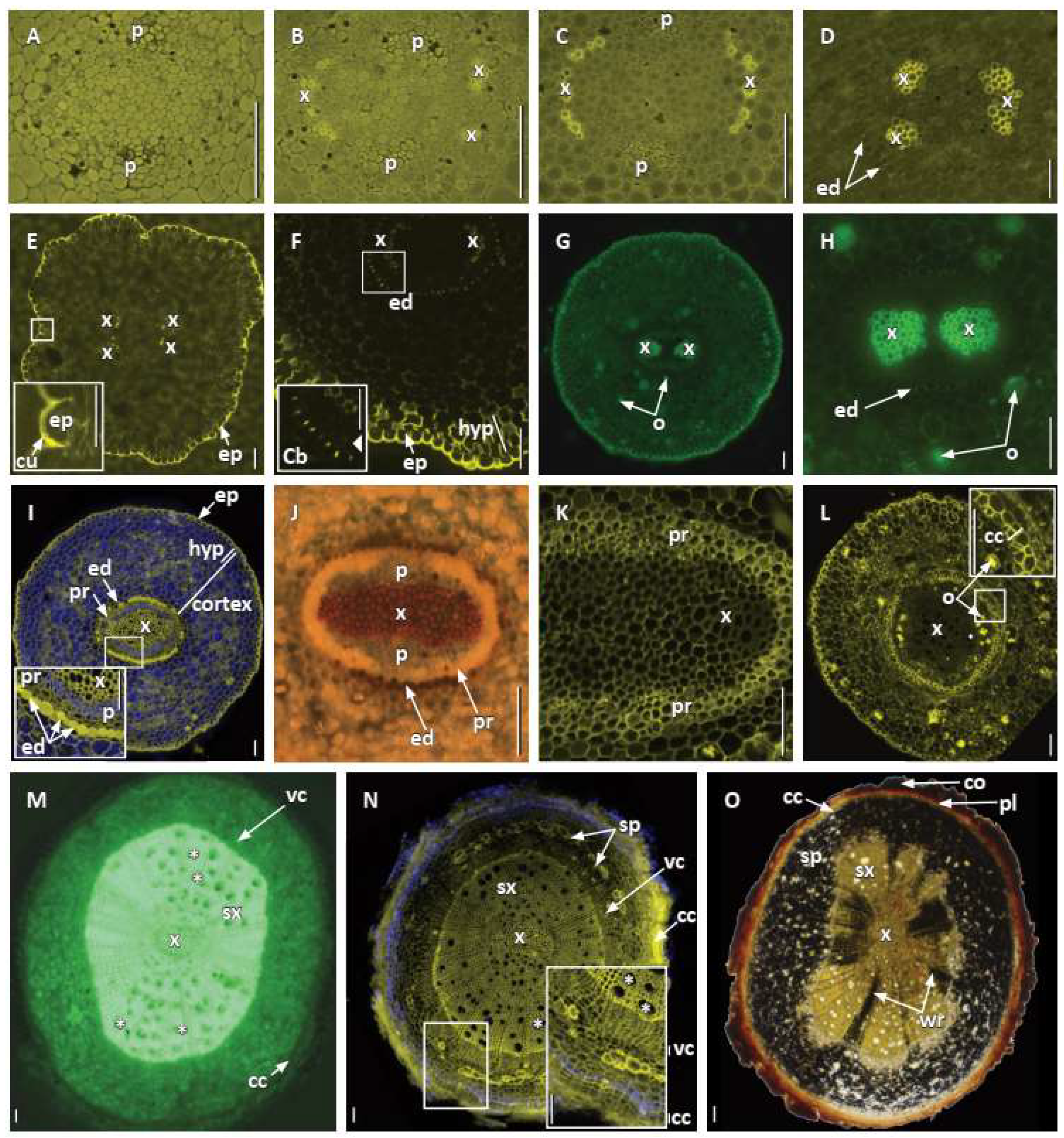
3.2. Primary Root Growth and Tissue Differentiation
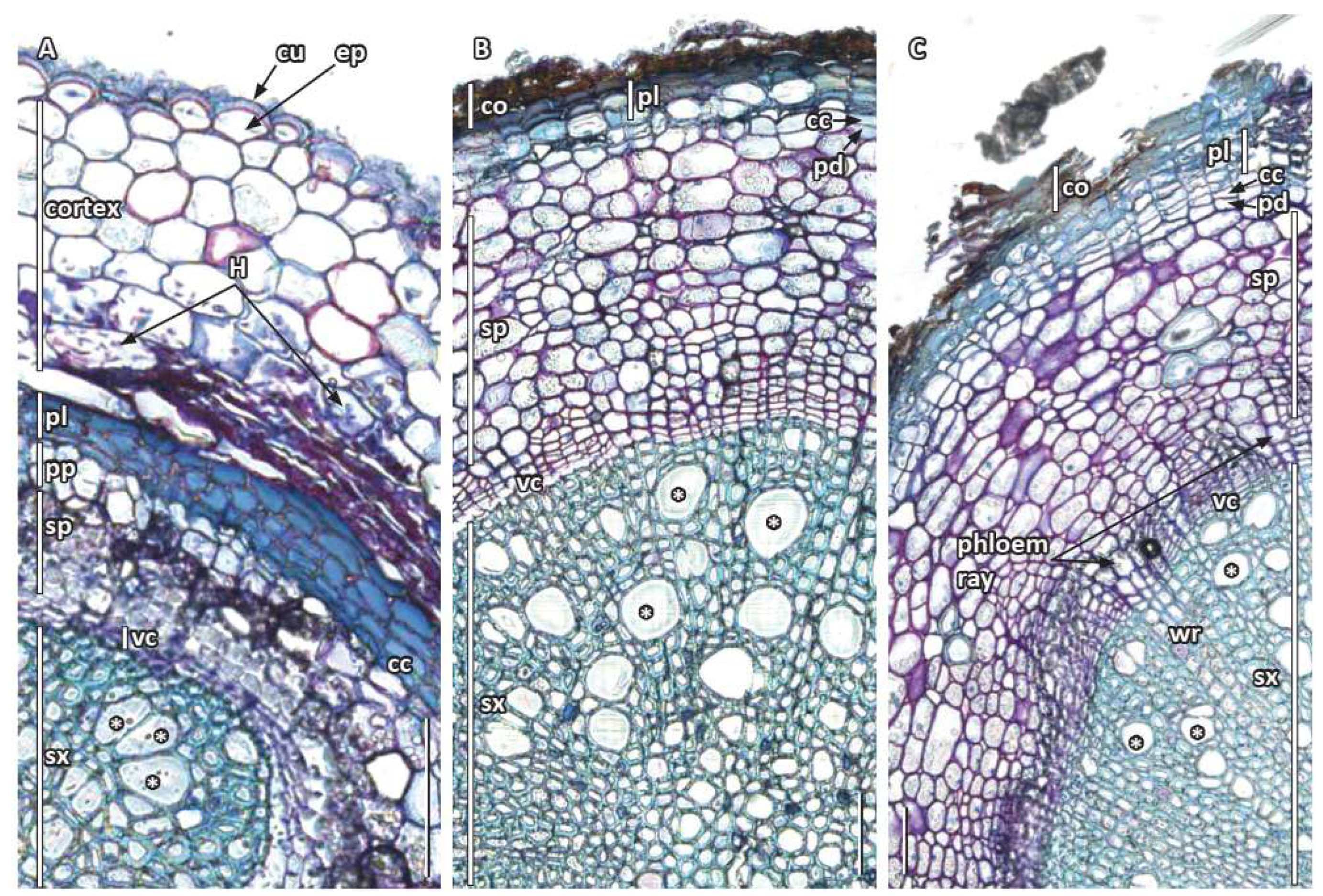
3.3. Secondary Root Growth and Tissue Differentiation
4. Discussion
5. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sauquet, H.; von Balthazar, M.; Doyle, J.A.; Endress, P.K.; Bailes, E.J.; Carrive, L.; Chartier, M.; Chomicki, G.; Little, S.A.; Luna, J.A.; et al. The ancestral flower of angiosperms and its early diversification. Nat. Commun. 2017, 16047, 1–10. [Google Scholar] [CrossRef]
- Soltis, D.; Soltis, P.; Endress, P.K.; Chase, M.; Manchester, S.; Judd, W.; Majure, L.; Mavrodiev, E. Phylogeny and Evolution of the Angiosperms; The University of Chicago Press: Chicago, IL, USA, 2018. [Google Scholar]
- Seago, J.L., Jr.; Marsh, L.C.; Stevens, K.J.; Soukup, A.; Votrubová, O.; Enstone, D.E. A re-examination of the root cortex in wetland flowering plants with respect to aerenchyma. Ann. Bot. 2005, 96, 565–579. [Google Scholar] [CrossRef]
- Heimsch, C.; Seago, J.L., Jr. Organization of the root apical meristem in angiosperms. Am. J. Bot. 2008, 95, 1–21. [Google Scholar] [CrossRef]
- Seago, J.L., Jr.; Fernando, D.D. Anatomical aspects of angiosperm root evolution. Ann. Bot. 2013, 112, 223–238. [Google Scholar] [CrossRef]
- Esau, K. Plant Anatomy; John Wiley & Sons: New York, NY, USA, 1953. [Google Scholar]
- Pillai, A. Root apical organization in gymnosperms—Some cycads and Ginkgo biloba. Proc. Indian Acad. Sci. B 1963, 57, 211–222. [Google Scholar]
- Pillai, A. Root apical organization in gymnosperms—Some conifers. Bull. Torrey Bot. Club 1964, 91, 1–13. [Google Scholar] [CrossRef]
- Bailey, I.W.; Swamy, B.G.L. The morphology and relationships of Austrobaileya. J. Arnold Arbor. 1949, 30, 211–226. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons: Leaves, Stem, and Wood in Relation to Taxonomy with Notes on Economic Uses; Oxford University Press: London, UK, 1950. [Google Scholar]
- Dickison, W.C.; Endress, P.K. Ontogeny of the stem-node-leaf vascular continuum of Austrobaileya. Am. J. Bot. 1983, 70, 906–911. [Google Scholar] [CrossRef]
- Baranova, M.A. The epidermal structures and systematic position of the Austrobaileyaceae. Bot. Zhurnal 1992, 77, 1–17. [Google Scholar]
- Baranova, M.A. The epidermal structure of Austrobaileya (Austrobaileyaceae): A further comment. Kew Bull. 2004, 59, 489–491. [Google Scholar] [CrossRef]
- Feild, T.S.; Franks, P.J.; Sage, T.L. Ecophysiological shade adaptation in the basal angiosperm Austrobaileya scandens (Austrobaileyaceae). Int. J. Plant Sci. 2003, 164, 313–324. [Google Scholar] [CrossRef]
- Carpenter, K.J. Stomatal architecture and evolution in basal angiosperms. Am. J. Bot. 2005, 92, 1595–1615. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K.J. Specialized structures in the leaf epidermis of basal angiosperms: Morphology, distribution, and homology. Am. J. Bot. 2006, 93, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, L.M. The secondary phloem of Austrobaileya scandens. Can. J. Bot. 1970, 48, 341–359. [Google Scholar] [CrossRef]
- Carlquist, S. Observations on the vegetative anatomy of Austrobaileya: Habital, organographic and phylogenetic conclusions. Bot. J. Linn. Soc. 2001, 135, 1–11. [Google Scholar] [CrossRef][Green Version]
- Jensen, W.A. Botanical Histochemistry; W. H. Freeman: San Francisco, CA, USA, 1962. [Google Scholar]
- Clowes, F.A.L. Pattern in root meristem development in angiosperms. New Phytol. Trust 2000, 146, 83–94. [Google Scholar] [CrossRef]
- Groot, E.P.; Doyle, J.A.; Nichol, S.A.; Rost, T.L. Phylogenetic distribution and evolution of root apical meristem organization in dicotyledonous angiosperms. Int. J. Plant Sci. 2004, 165, 87–105. [Google Scholar] [CrossRef]
- Clowes, F.A.L. The difference between open and closed meristems. Ann. Bot. 1981, 48, 761–767. [Google Scholar] [CrossRef]
- Bailey, I.W.; Swamy, B.G.L. Amborella trichopoda Baill, a new morphological type of dicotyledon. J. Arnold Arbor. 1948, 29, 245–254. [Google Scholar]
- Carlquist, S.; Schneider, E.L. Vegetative anatomy of the New Caledonian endemic Amborella trichopoda: Relationships with the Illiciales and implications for vessel origin. Pac. Sci. 2001, 55, 305–312. [Google Scholar] [CrossRef]
- Clowes, F.A.L. The structure of mycorrhizal roots of Fagus sylvatica. New Phytol. 1951, 50, 1–16. [Google Scholar] [CrossRef]
- Baylis, G.T.S. The Magnoliid mycorrhiza and mycotrophy in root systems derived from it. In Endomycorrhizas; Sanders, E.R., Mosse, B., Tinker, P.B., Eds.; Academic Press: London, UK, 1975; pp. 373–389. [Google Scholar]
- Losada, J.M.; Bachelier, J.B.; Friedman, W.E. Prolonged embryogenesis in Austrobaileya scandens (Austrobaileyaceae): Its ecological and evolutionary significance. New Phytol. 2017, 215, 851–864. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bachelier, J.B.; Razik, I.; Schauer, M.; Seago, J.L., Jr. Roots Structure and Development of Austrobaileya scandens (Austrobaileyaceae) and Implications for Their Evolution in Angiosperms. Plants 2020, 9, 54. https://doi.org/10.3390/plants9010054
Bachelier JB, Razik I, Schauer M, Seago JL Jr. Roots Structure and Development of Austrobaileya scandens (Austrobaileyaceae) and Implications for Their Evolution in Angiosperms. Plants. 2020; 9(1):54. https://doi.org/10.3390/plants9010054
Chicago/Turabian StyleBachelier, Julien B., Imran Razik, Maria Schauer, and James L. Seago, Jr. 2020. "Roots Structure and Development of Austrobaileya scandens (Austrobaileyaceae) and Implications for Their Evolution in Angiosperms" Plants 9, no. 1: 54. https://doi.org/10.3390/plants9010054
APA StyleBachelier, J. B., Razik, I., Schauer, M., & Seago, J. L., Jr. (2020). Roots Structure and Development of Austrobaileya scandens (Austrobaileyaceae) and Implications for Their Evolution in Angiosperms. Plants, 9(1), 54. https://doi.org/10.3390/plants9010054