Physiological Responses of Basil (Ocimum Basilicum L.) Cultivars to Rhizophagus Irregularis Inoculation under Low Phosphorus Availability




Abstract
1. Introduction
2. Results
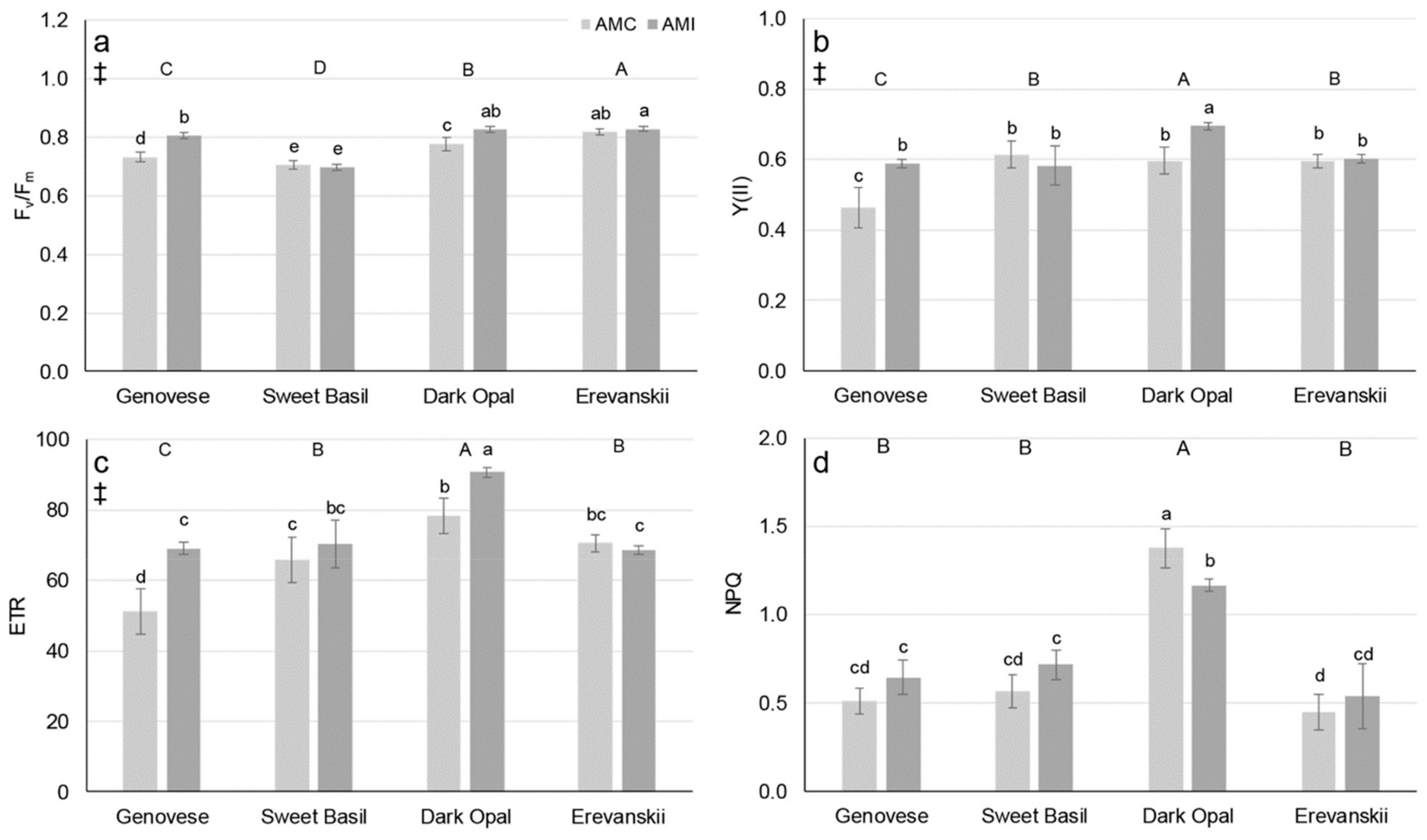
2.1. Chlorophyll Fluorescence
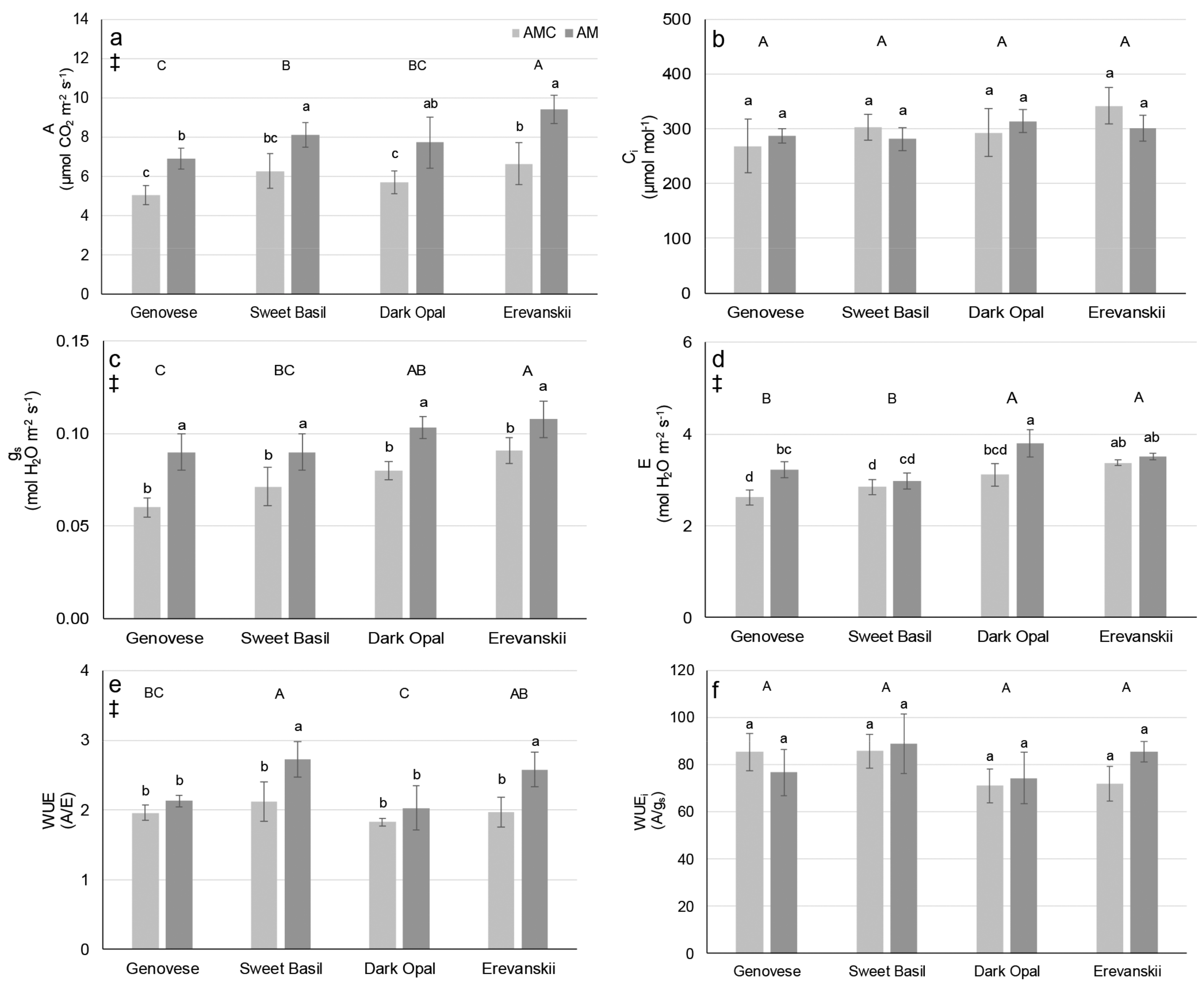
2.2. Gas Exchange
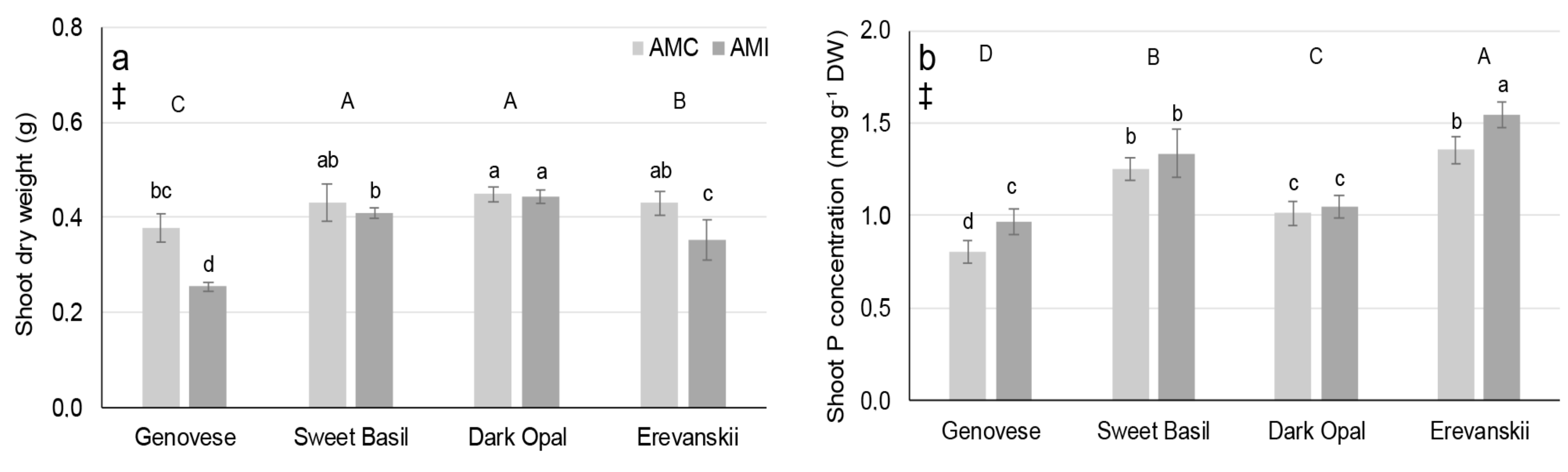
2.3. Shoot Biomass and Shoot Mineral Concentration
2.4. Root Morphology
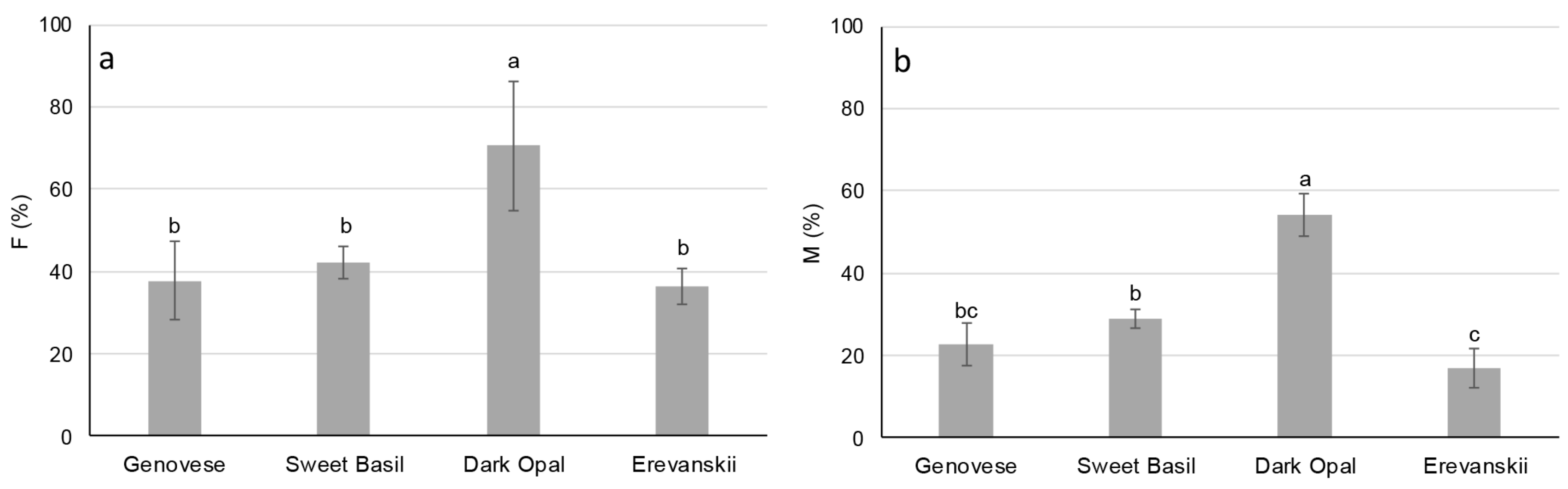
2.5. Arbuscular Mycorrhizas (AM) Root Colonization
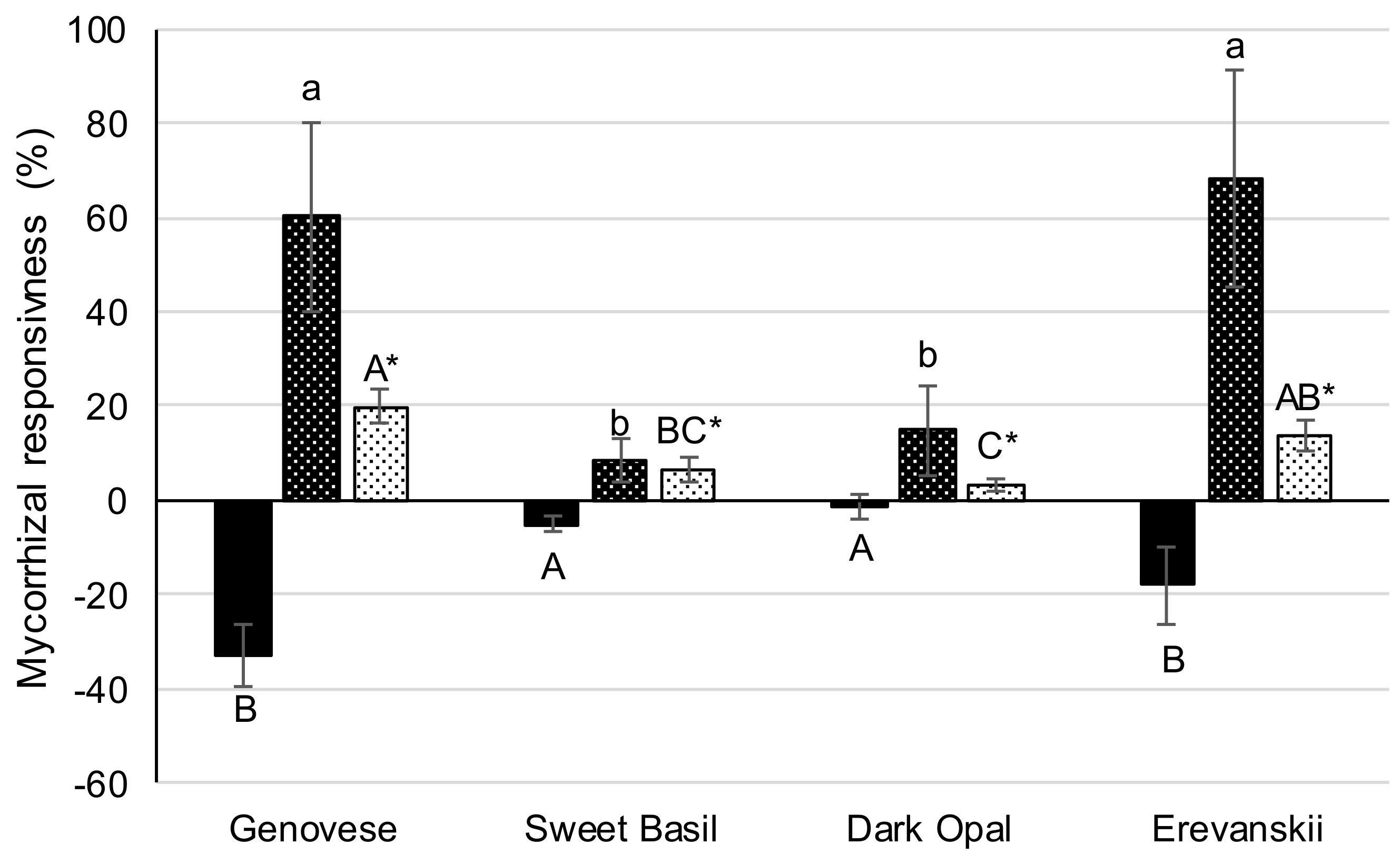
2.6. Mycorrhizal Responsiveness
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Experimental Conditions
5.2. Chlorophyll Fluorescence and Gas-Exchange Measurements
5.3. Shoot Biomass and Mineral Concentration Analysis
5.4. Root System Measurements
5.5. AM Root Colonization Analysis
5.6. Mycorrhizal Responsiveness
- (1)
- The improvement in shoot biomass as the percent difference in the shoot dry weight between the AM and non-mycorrhizal plants [22],
- (2)
- The improvement of P nutrition, as the percent difference in the shoot P concentration between the AM and non-mycorrhizal plants [43]
- (3)
- The improvement in root growth as the percent difference in the total root length.
5.7. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Lynch, J.P. Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Brooks, A. Effects of phosphorus nutrition on ribulose-1,5-biophosphate carboxylase activation, photosynthetic quantum yield and amounts of some Calvin cycle metabolites in spinach leaves. Aust. J. Plant Physiol. 1986, 13, 221–237. [Google Scholar]
- Plesnicar, M.; Kastori, R.; Petrovic, N.; Pankovic, D. Photosynthesis and chlorophyll fluorescence in sunflower (Helianthus annuus L.) leaves as affected by phosphorus-nutrition. J. Exp. Bot. 1994, 45, 919–924. [Google Scholar] [CrossRef]
- Usuda, H.; Shimogawara, K. Phosphate deficiency in maize.1. Leaf phosphate status, growth, photosynthesis and carbon partitioning. Plant Cell Physiol. 1991, 32, 497–504. [Google Scholar]
- Lynch, J.; Lauchli, A.; Epstein, E. Vegetative growth of the common bean in response to phosphorus nutrition. Crop Sci. 1991, 31, 380–387. [Google Scholar] [CrossRef]
- Rao, I.M.; Fredeen, A.L.; Terry, N. Influence of phosphorus limitation on photosynthesis, carbon allocation and partitioning in sugar beet and soybean grown with a short photoperiod. Plant Physiol. Biochem. 1993, 31, 223–231. [Google Scholar]
- López-Bucio, J.; Cruz-Ramírez, A.; Herrera-Estrella, L. The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 2003, 6, 280–287. [Google Scholar] [CrossRef]
- Syers, J.K.; Johnston, A.E.; Curtin, D. Efficiency of soil and fertilizer phosphorus use: Reconciling changing concepts of soil phosphorus behaviour with agronomic information. In FAO Fertilizer and Plant Nutrition Bulletin 18; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2008. [Google Scholar]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, MA, USA; Elsevier Ltd.: London, UK, 2008. [Google Scholar]
- Simon, J.E.; Quinn, J.; Murray, R.G. Basil: A source of essential oils. In Advances in New Crops; Janick, J., Simon, J.E., Eds.; Timber Press: Portland, OR, USA, 1990; pp. 484–989. [Google Scholar]
- Carović-Stanko, K.; Šalinović, A.; Grdiša, M.; Liber, Z.; Kolak, I.; Šatović, Z. Efficiency of morphological trait descriptors in discrimination of Ocimum basilicum L. accessions. Plant Biosyst. 2011, 145, 298–305. [Google Scholar] [CrossRef]
- Varga, F.; Carović-Stanko, K.; Ristić, M.; Grdiša, M.; Liber, Z.; Šatović, Z. Morphological and biochemical intraspecific characterization of Ocimum basilicum L. Ind. Crop. Prod. 2017, 109, 611–618. [Google Scholar] [CrossRef]
- Toussaint, J.-P. The effect of the arbuscular mycorrhizal symbiosis on the production of phytochemicals in basil, degree of Doctor of Philosophy. In Faculty of Sciences Discipline of Soil and Land Systems School of Earth and Environmental Sciences; The University of Adelaide: Adelaide, Australia, May 2008. [Google Scholar]
- Scagel, C.F.; Lee, J. Phenolic Composition of Basil Plants Is Differentially Altered by Plant Nutrient Status and Inoculation with Mycorrhizal Fungi. HortScience 2012, 47, 660–671. [Google Scholar] [CrossRef]
- Rasouli-Sadaghiani, M.; Hassani, A.; Mohsen Barin, M.; Danesh, Y.R.; Sefidkon, F. Effects of arbuscular mycorrhizal (AM) fungi on growth, essential oil production and nutrients uptake in basil. J. Med. Plant Res. 2010, 4, 2222–2228. [Google Scholar]
- Hazzoumi, Z.; Moustakime, Y.; Elharchli, E.; Khalid, A.J. Effect of arbuscular mycorrhizal fungi (AMF) and water stress on growth, phenolic compounds, glandular hairs, and yield of essential oil in basil (Ocimum gratissimum L). Chem. Biol. Technol. Agric. 2015, 2, 10. [Google Scholar] [CrossRef]
- Zuccarini, P.; Okurowska, P. Effects of Mycorrhizal Colonization and Fertilization on Growth and Photosynthesis of Sweet Basil Under Salt Stress. J. Plant Nutr. 2008, 31, 497–513. [Google Scholar] [CrossRef]
- Klironomos, J.N. Variation in plant response to native and exotic arbuscular mycorrhizal fungi. Ecology 2003, 84, 2292–2301. [Google Scholar] [CrossRef]
- Janos, D.P. Plant responsiveness to mycorrhizas differs from dependence upon mycorrhizas. Mycorrhiza 2007, 17, 75–91. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A.; Jakobsen, I. Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol. 2003, 133, 16–20. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A.; Jakobsen, I. Functional diversity in arbuscular mycorrhizal AM symbioses: The contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake. New Phytol. 2004, 162, 511–524. [Google Scholar] [CrossRef]
- Liu, H.; Song, F.; Liu, S.; Li, X.; Liu, F.; Zhu, X. Arbuscular mycorrhiza improves nitrogen use efficiency in soybean grown under partial root-zone drying irrigation. Arch. Agron. Soil Sci. 2019, 65, 269–279. [Google Scholar] [CrossRef]
- Feldmann, F.; Gillessen, M.; Hutter, I.; Schneider, C. Should we breed for effective mycorrhiza symbioses? In Crop Plant Resistance to Biotic and Abiotic Factors; Feldmann, F., Alford, D.V., Furk, C., Eds.; Phytomedizinische Gesellschaft: Braunschweig, Germany, 2009; pp. 507–522. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: Cambridge, MA, USA; Elsevier Ltd.: London, UK, 1995; pp. 275–277. [Google Scholar]
- Ličina, V.Đ.; Jelačić, S.Ć.; Beatović, D.V.; Antić Mladenović, S.B. Mineral composition of different basil (Ocimum spp.) genotypes. Hem. Ind. 2014, 68, 501–510. [Google Scholar] [CrossRef]
- Rychter, A.M.; Rao, I.M. Role of phosphorus in photosynthetic carbon metabolism. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Ed.; CRC Press: Cambridge, MA, USA; Taylor & Francis Group: Boca Raton, FL, USA, 2005; pp. 138–163. [Google Scholar]
- Eissenstat, D.M.; Graham, J.H.; Syvertsen, J.P.; Drouillard, D.L. Carbon economy of sour orange in relation to mycorrhizal colonization and phosphorus status. Ann. Bot. 1993, 71, 1–10. [Google Scholar] [CrossRef]
- Ravnskov, S.; Wu, Y.; Graham, J.H. Arbuscular mycorrhizal fungi differentially affect expression of genes coding for sucrose synthases in maize roots. New Phytol. 2003, 157, 539–545. [Google Scholar] [CrossRef]
- Tekaya, M.; Mechri, B.; Mbarki, N.H.; Cheheb, H.M.; Hammami, M.; Attia, F. Arbuscular mycorrhizal fungus Rhizophagus irregularis influences key physiological parameters of olive trees (Olea europaea L.) and mineral nutrient profile. Photosynthetica 2011, 55, 308–316. [Google Scholar] [CrossRef]
- Paul, M.J.; Foyer, C.H. Sink regulation of photosynthesis. J. Exp. Bot. 2011, 52, 1383–1400. [Google Scholar] [CrossRef]
- Fitter, A.H. Water relations of red clover, Trifolium pratense L., as affected by VA mycorrhizal infection and phosphorus supply before and during drought. J. Exp. Bot. 1988, 39, 595–604. [Google Scholar] [CrossRef]
- Druge, U.; Schonbeck, F. Effect of vesicular-arbuscular mycorrhizal infection on transpiration, photosynthesis and growth of flax Linum usitatissimum L. in relation to cytokinin levels. J. Plant Physiol. 1993, 141, 40–48. [Google Scholar] [CrossRef]
- Allen, M.F.; Smith, W.K.; Moore, T.S.; Christensen, M. Comparative water relations and non-mycorrhizal Bouteloua gracilis H.B.K. Lag ex Steud. New Phytol. 1981, 88, 683–693. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef]
- AOAC. Official Method of Analysis of AOAC International, 16th ed.; AOAC International: Arlington, VA, USA, 1995; Volume I. [Google Scholar]
- Schreiber, U.; Bilge, W.; Neubauer, C. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis; Schulze, E.D., Caldwell, M.M., Eds.; Springer-Verlag: Berlin, Germany, 1994; pp. 49–70. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio RFd of leaves with the PAM fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Meesure du taux de mycorhization VA d’un système radiculaire. Recherche des méthodes d’estimation ayant une signification fonctionnelle. In Physiological Aspect of Mycorrizae; Bay, G.-P., Gianinazzi, S., Eds.; INRA: Paris, France, 1984; pp. 217–221. [Google Scholar]
- Mycocalc program-INRA de Dijon. Available online: https://www2.dijon.inra.fr/mychintec/Mycocalc-prg/download.html (accessed on 4 July 2019).
- Zhu, Y.-G.; Smith, S.E.; Barritt, A.R.; Smith, F.A. Phosphorus (P) efficiencies and mycorrhizal responsivness of old and modern wheat cultivars. Plant Soil 2001, 237, 249–255. [Google Scholar] [CrossRef]
- SAS Institute Inc. Base SAS® 9.3 Procedures Guide; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | AM inoculation | Ca (%) | Mg (%) | Fe (mg g−1 DW) | Mn (mg g−1 DW) | Zn (mg g−1 DW) |
|---|---|---|---|---|---|---|
| ‘Genovese’ | AMC | 1.29 ± 0.30 b | 0.45 ± 0.06 a | 86.2 ± 7.9 ab | 54.1 ± 4.9 a | 22.7 ± 1.5 b |
| AMI | 1.16 ± 0.18 b | 0.43 ± 0.06 a | 82.3 ± 8.2 ab | 52.5 ± 6.5 a | 20.5 ± 1.5 b | |
| ‘Sweet Basil’ | AMC | 1.26 ± 0.17 b | 0.44 ± 0.09 a | 83.0 ± 6.1 ab | 49.1 ± 3.3 a | 20.7 ± 1.4 b |
| AMI | 1.12 ± 0.17 b | 0.45 ± 0.08 a | 83.0 ± 7.0 ab | 46.8 ± 3.2 a | 22.3 ± 2.7 b | |
| ‘Dark Opal’ | AMC | 1.08 ± 0.23 b | 0.30 ± 0.06 b | 93.6 ± 9.3 a | 47.6 ± 4.8 a | 31.5 ± 3.2 a |
| AMI | 1.23 ± 0.22 b | 0.31 ± 0.05 b | 90.8 ± 10.2 a | 49.9 ± 7.0 a | 32.5 ± 2.3 a | |
| ‘Erevanskii’ | AMC | 1.58 ± 0.15 a | 0.34 ± 0.09 b | 79.9 ± 9.7 b | 45.4 ± 6.2 a | 21.4 ± 1.5 b |
| AMI | 1.48 ± 0.15 a | 0.32 ± 0.05 b | 75.3 ± 9.2 b | 43.3 ± 6.3 a | 20.3 ± 2.2 b | |
| Source of Variation | Significance level (P) | |||||
| Cultivar (C) | <0.01 | <0.01 | <0.05 | n.s. | <0.001 | |
| Inoculation Treatment (M) | n.s. | n.s. | n.s. | n.s. | n.s. | |
| C × M | n.s. | n.s. | n.s. | n.s. | n.s | |
| Cultivar | AM Inoculation | Depth (cm) | Width (cm) | Length (cm) | Surface Area (cm2) | Volume (cm3) | Diameter (mm) | Number of Tips |
|---|---|---|---|---|---|---|---|---|
| ‘Genovese’ | AMC | 13.3 ± 0.6 b | 8.1 ± 0.7 ab | 187.6 ± 23.1 b | 6.0 ± 0.4 b | 0.18 ± 0.018 bc | 0.32 ± 0.025 ab | 111.7 ± 14.1 b |
| AMI | 21.3 ± 1.2 a | 10.4 ± 2.1 ab | 334.4 ± 37.1 a | 11.3 ± 1.7 a | 0.32 ± 0.029 a | 0.27 ± 0.014 b | 317.5 ± 88.7 a | |
| ‘Sweet Basil’ | AMC | 12.1 ± 1.0 b | 5.4 ± 1.1 c | 108.7 ± 19.3 b | 3.9 ± 1.0 b | 0.11 ± 0.037 cd | 0.36 ± 0.001 a | 110.0 ± 39.8 b |
| AMI | 13.1 ± 1.2 b | 6.6 ± 1.2 bc | 114.1 ± 45.7 b | 3.8 ± 0.6 b | 0.11 ± 0.040 cd | 0.33 ± 0.016 a | 102.8 ± 31.5 b | |
| ‘Dark Opal’ | AMC | 10.7 ± 1.1 c | 7.3 ± 2.5 bc | 143.0 ± 53.1 b | 3.9 ± 0.5 b | 0.08 ± 0.010 d | 0.27 ± 0.006 b | 98.3 ± 10.62 b |
| AMI | 12.3 ± 2.0 b | 7.5 ± 0.7 bc | 186.1 ± 18.1 b | 5.3 ± 1.5 b | 0.12 ± 0.017 cd | 0.28 ± 0.014 b | 115.8 ± 18.7 b | |
| ‘Erevanskii’ | AMC | 12.6 ± 1.2 b | 8.1 ± 1.1 abc | 197.2 ± 24.2 b | 4.7 ± 0.5 b | 0.12 ± 0.011 cd | 0.26 ± 0.011 bc | 140.0 ± 60.0 b |
| AMI | 21.2 ± 2.4 a | 12.1 ± 1.5 a | 388.2 ± 33.1 a | 10.1 ± 0.9 a | 0.21 ± 0.019 b | 0.23 ± 0.004 c | 331.2 ± 86.5 a | |
| Source of Variation | Significance level (P) | |||||||
| Cultivar (C) | <0.001 | <0.01 | <0.001 | <0.01 | <0.001 | <0.001 | <0.001 | |
| Inoculation Treatment (M) | <0.001 | <0.05 | <0.001 | <0.01 | <0.001 | <0.05 | <0.01 | |
| C × M | <0.001 | n.s. | <0.05 | <0.05 | <0.05 | n.s. | <0.01 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazarević, B.; Carović-Stanko, K.; Šatović, Z. Physiological Responses of Basil (Ocimum Basilicum L.) Cultivars to Rhizophagus Irregularis Inoculation under Low Phosphorus Availability. Plants 2020, 9, 14. https://doi.org/10.3390/plants9010014
Lazarević B, Carović-Stanko K, Šatović Z. Physiological Responses of Basil (Ocimum Basilicum L.) Cultivars to Rhizophagus Irregularis Inoculation under Low Phosphorus Availability. Plants. 2020; 9(1):14. https://doi.org/10.3390/plants9010014
Chicago/Turabian StyleLazarević, Boris, Klaudija Carović-Stanko, and Zlatko Šatović. 2020. "Physiological Responses of Basil (Ocimum Basilicum L.) Cultivars to Rhizophagus Irregularis Inoculation under Low Phosphorus Availability" Plants 9, no. 1: 14. https://doi.org/10.3390/plants9010014
APA StyleLazarević, B., Carović-Stanko, K., & Šatović, Z. (2020). Physiological Responses of Basil (Ocimum Basilicum L.) Cultivars to Rhizophagus Irregularis Inoculation under Low Phosphorus Availability. Plants, 9(1), 14. https://doi.org/10.3390/plants9010014