HD-AGPs as Speciation Genes: Positive Selection on a Proline-Rich Domain in Non-Hybridizing Species of Petunia, Solanum, and Nicotiana

Abstract

1. Introduction
2. Results
2.1. Breeding Behavior in Petunia Accessions
2.2. Time Course of Conspecific and Interspecific Pollination
2.3. HD-AGPs Sequences from Petunia Species
2.4. Domain Structure of Petunia HD-AGPs
2.5. Tetrapeptide Indels Are Associated with Reproductive Barriers
2.6. Ka/Ks Metric Lacks Sensitivity to Detect Evolutionary Changes in Closely-Related Taxa
2.7. Comparison of HD-AGPs from Nicotiana
2.8. Comparison of HD-AGPs from Solanum
2.9. Comparison of HD-AGPs from Petunia, Nicotiana, Solanum, and Capsicum
2.10. Association of XKPP Variants and IRBs in 15 Species-Pairs
2.11. HD-AGPs Exhibit Low within-Species Polymorphism
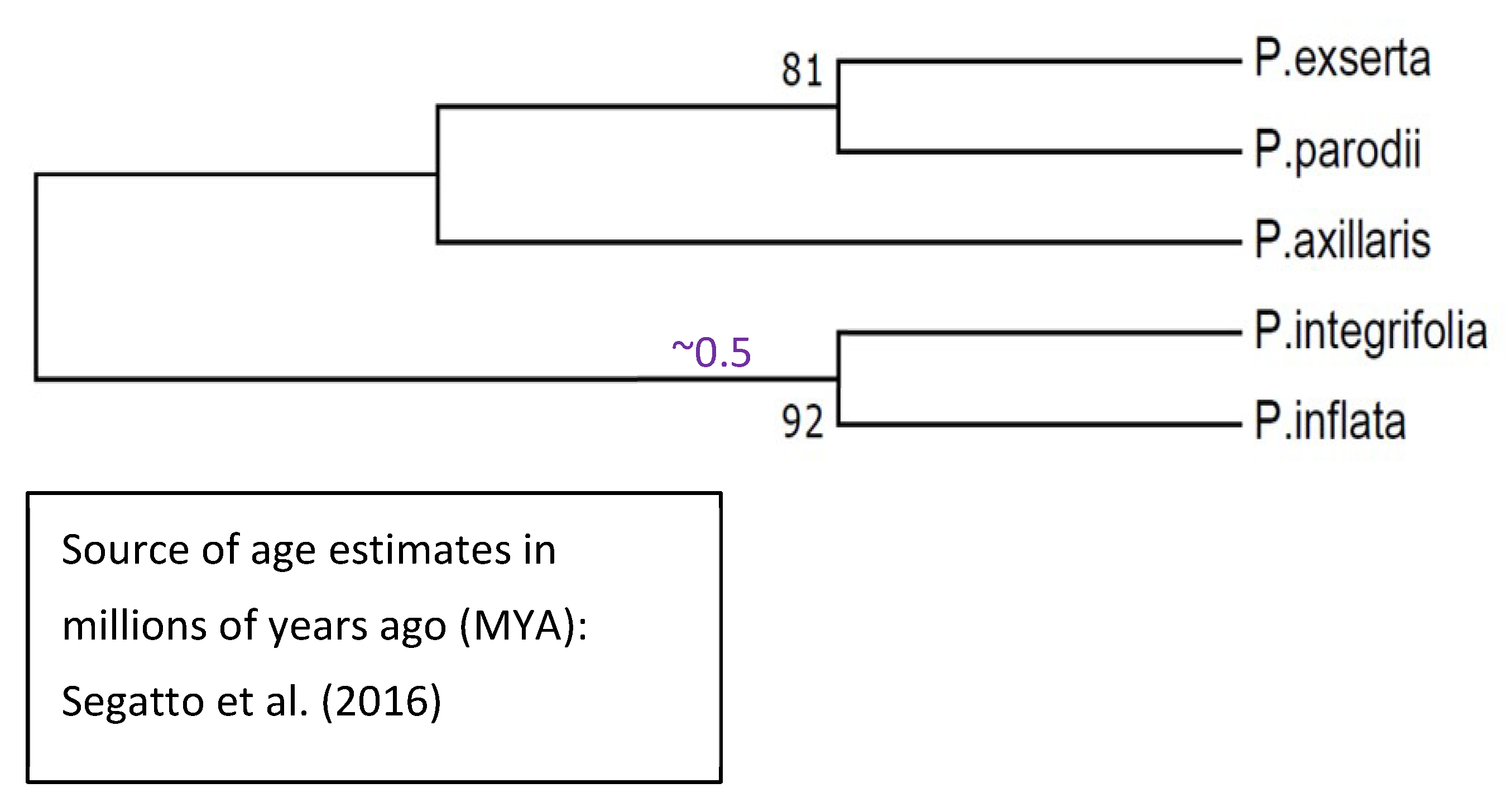
2.12. The HD-AGP Gene Tree Is Broadly Consistent With Well-Known Phylogenies
3. Discussion
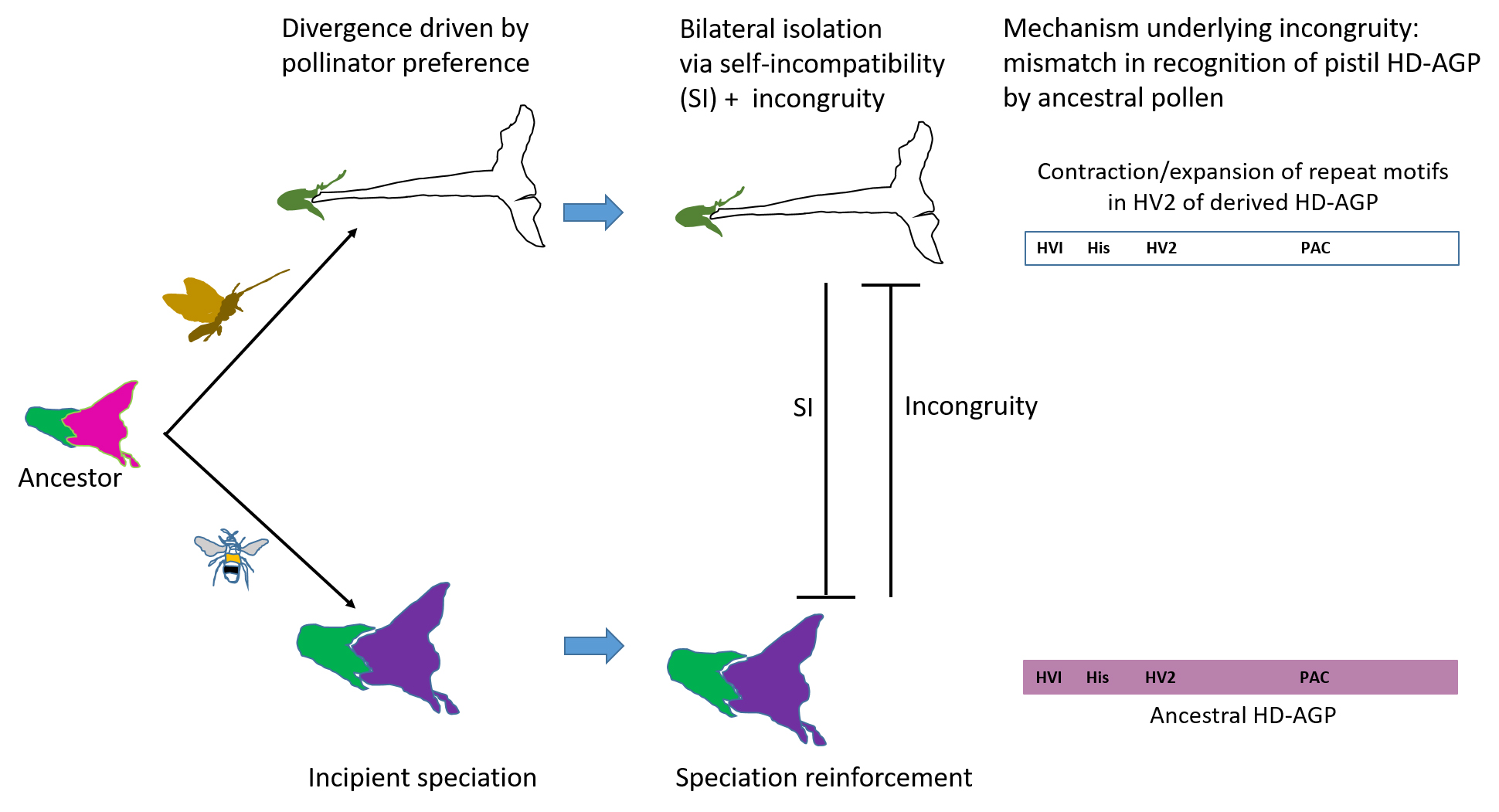
3.1. The Hypothesis: Sequence Divergence in HD-AGPs Generates Incongruity
3.2. HD-AGPs Are Identical or Near-Identical in Cross-Hybridizing Petunia Species
3.3. Tetrapeptide Indels in Petunia HD-AGPs Co-Vary with Interspecific Incongruity
3.4. Incongruity in Nicotiana Too Is Linked with Variation in XKPP Repeat Motifs
3.5. Incongruity in Most Solanum Species-Pairs Is also Associated with Variation in XKPP Repeat Motifs
3.6. There Is Positive Selection on HV1 and HV2, with Purifying Selection on Other HD-AGP Domains
3.7. Variation in XKPP Motifs Co-Varies with the Presence of Reproductive Barriers
3.8. Reinforcement of Sympatric Speciation by Postmating Prezygotic IRBS: A Model
4. Materials and Methods
4.1. Plants
4.2. Time Course of Pollen Tube Growth Following Conspecific and Heterospecific Pollinations
4.3. RNA Isolation and DNA Sequencing
4.4. Sequence Analysis
4.5. Bioinformatics and Statistical Analysis
4.6. Evolutionary Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Baack, E.; Melo, M.C.; Rieseberg, L.H.; Ortiz-Barrientos, D. The origins of reproductive isolation in plants. New Phytol. 2015, 207, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Rieseberg, L.H.; Willis, J.H. Plant speciation. Science. 2007, 317, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, R. Reinforcement in plants. New Phytol. 2013, 197, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Dobzhansky, T. Speciation as a stage in evolutionary divergence. Am. Nat. 1940, 74, 312–321. [Google Scholar]
- Grant, V. Evidence for the selective origin of incompatibility barriers in the leafy-stemmed gilias. Proc. Natl. Acad. Sci. USA 1965, 54, 1567–1571. [Google Scholar] [CrossRef] [PubMed]
- Nosil, P.; Schluter, D. The genes underlying the process of speciation. Trends Ecol. Evol. 2011, 26, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Rieseberg, L.H.; Blackman, B.K. Speciation genes in plants. Ann. Bot. 2010, 106, 439–455. [Google Scholar] [CrossRef]
- Venail, J.; Dell’Olivo, A.; Kuhlemeier, C. Speciation genes in the genus Petunia. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 461–468. [Google Scholar] [CrossRef]
- Hoballah, M.E.; Gubitz, T.; Stuurman, J.; Broger, B.; Barone, M.; Mandel, T.; Dell’Olivo, A.; Arnold, M.; Kuhlemeier, C. Single Gene–Mediated Shift in Pollinator Attraction in Petunia. Plant Cell 2007, 19, 779–790. [Google Scholar] [CrossRef]
- Brosi, B.J. Pollinator specialization: From the individual to the community. New Phytol. 2016, 210, 1190–1194. [Google Scholar] [CrossRef]
- Dell’olivo, A.; Hoballah, M.E.; Gübitz, T.; Kuhlemeier, C. Isolation barriers between Petunia axillaris and Petunia integrifolia (Solanaceae). Evolution 2011, 65, 1979–1991. [Google Scholar] [CrossRef] [PubMed]
- Fujii, D.; Kubo, K.-I.; Takayama, S. Non-self- and self-recognition models in plant self-incompatibility. Nat. Plants 2016, 2, 16130. [Google Scholar] [CrossRef]
- Hancock, C.N.; Kondo, K.; Beecher, B.; McClure, B. The S-locus and unilateral incompatibility. Philos. Trans. R. Soc. Lond. B. 2003, 358, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- McClure, B.A.; Cruz-Garcia, F.; Beecher, B.S.; Sulaman, W. Factors affecting inter- and intra-specific pollen rejection in Nicotiana. Ann. Bot. 2000, 85, 113–123. [Google Scholar] [CrossRef]
- Jiménez-Durán, K.; McClure, B.; García-Campusano, F.; Rodríguez-Sotres, R.; Cisneros, J.; Busot, G.; Cruz-García, F. NaStEP: A Proteinase Inhibitor Essential to Self-Incompatibility and a Positive Regulator of HT-B Stability in Nicotiana alata Pollen Tubes. Plant Physiol. 2013, 161, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Baek, Y.S.; Covey, P.A.; Petersen, J.J.; Chetelat, R.T.; McClure, B.; Bedinger, P.A. Testing the SI × SC rule: Pollen-pistil interactions in interspecific crosses between members of the tomato clade (Solanum section Lycopersicon, Solanaceae). Am. J. Bot. 2015, 102, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Garcia, F.; Hancock, C.N.; McClure, B. S-RNase complexes and pollen rejection. J. Exp. Bot. 2003, 59, 123–130. [Google Scholar] [CrossRef]
- Tovar-Méndez, A.; Lu, L.; McClure, B. HT proteins contribute to S-RNase-independent pollen rejection in Solanum. Plant J. 2017, 89, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Baek, Y.D.; Royer, S.M.; Broz, A.K.; Covey, P.A.; López-Casado, G.; Nuñez, R.; Kear, P.J.; Bonierbale, M.; Orillo, M.; van der Knaap, E.; et al. Interspecific reproductive barriers between sympatric populations of wild tomato species (Solanum section Lycopersicon). Am. J. Bot. 2016, 103, 1964–1978. [Google Scholar] [CrossRef]
- Noyszewski1, A.K.; Liu, Y.-C.; Tamura, K.; Smith, A.G. Polymorphism and structure of style–specific arabinogalactan proteins as determinants of pollen tube growth in Nicotiana. BMC Evol. Biol. 2017, 17, 186. [Google Scholar] [CrossRef]
- Cheung, A.Y. The pollen tube growth pathway: Its molecular and biochemical contributions and responses to pollination. Sex. Plant Reprod. 1996, 9, 330–336. [Google Scholar] [CrossRef]
- Twomey, M. Characterization of PhPRP1, a histidine domain arabinogalactan protein from Petunia hybrida pistils. J. Plant Physiol. 2013, 170, 1384–1388. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.M.; Pereira, L.G.; Coimbra, S. Arabinogalactan proteins: Rising attention from plant biologists. Plant Reprod. 2015, 28, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Turchetto, C.; Lima, J.S.; Rodrigues, D.M.; Bonatto, S.L.; Freitas, L.B. Pollen dispersal and breeding structure in a hawkmoth-pollinated Pampa grasslands species Petunia axillaris (Solanaceae). Ann. Bot. 2015, 115, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Lubliner, N.; Singh-Cundy, D.T.; Singh-Cundy, A. Characterization of the pollen growth transition (PGT) in self-incompatible Petunia inflata. Sex. Plant Reprod. 2003, 15, 243–253. [Google Scholar]
- Brooks, J.K. Histidine Domain-Arabinogalactan Proteins (HD-AGPs) in the Solanaceae. Master’s Thesis, Western Washington University, Bellingham, WA, USA, 2016. [Google Scholar]
- Bombarely, A.; Moser, M.; Amrad, A.; Bao, M.; Bapaume, L.; Barry, C.S.; Bliek, M.; Boersma, M.R.; Borghi, L.; Bruggmann, R.; et al. Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrid. Nat. Plants 2016, 2, 16074. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.Y.; May, B.; Kawata, E.; Gu, Q.; Wu, H.-M. Characterization of cDNAs for stylar transmitting tissue-specific proline-rich proteins in tobacco. Plant J. 1993, 3, 151–160. [Google Scholar] [CrossRef]
- Christoff, M. Cytological Studies in the Genus Nicotiana. Genetics 1928, 13, 233–277. [Google Scholar]
- Lee, C.B.; Page, L.E.; McClure, B.A.; Holtsford, T.P. Postpollination hybridization barriers in Nicotiana Section Alatae. Sex. Plant Reprod. 2008, 21, 183–195. [Google Scholar] [CrossRef]
- Liu, H.; Marubashi, W. Species Origin of Genomic Factors in Nicotiana nudicaulis Watson Controlling Hybrid Lethality in Interspecific Hybrids between N. nudicaulis Watson and N. tabacum L. PLoS ONE 2014, 9, e97004. [Google Scholar] [CrossRef]
- Grandillo, S.; Tanksley, S.D. Genetic analysis of RFLPs, GATA microsatellites and RAPDs in a cross between L. esculentum and L. pimpinellifolium. Theor. Appl. Genet. 1996, 92, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Chalivendra, S.C.; Lopez-Casado, G.; Kumar, A.; Kassenbrock, A.R.; Royer, S.; Tovar-Mèndez, A.; Covey, P.A.; Dempsey, L.A.; Randle, A.M.; Stack, S.M.; et al. Developmental onset of reproductive barriers and associated proteome changes in stigma/styles of Solanum pennellii. J. Exp. Bot. 2013, 64, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed]
- Särkinen, T.; Bohs, L.; Olmstead, R.G.; Knapp, S. A phylogenetic framework for evolutionary study of the nightshades (Solanaceae): A dated 1000-tip tree. BMC Evol. Biol. 2013, 13, 214. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, J.J.; Dodsworth, S.; Chase, M.W. Time-calibrated phylogenetic trees establish a lag between polyploidisation and diversification in Nicotiana (Solanaceae). Plant Syst. Evol. 2017, 303, 1001–1012. [Google Scholar] [CrossRef]
- Segatto, A.L.A.; Reck-Kortmann, M.; Turchetto, C.; Freitas, L.B. Multiple markers, niche modelling, and bioregions analyses to evaluate the genetic diversity of a plant species complex. BMC Evol. Biol. 2017, 17, 234. [Google Scholar] [CrossRef] [PubMed]
- Lorenz-Lemke, A.P.; Togni, P.D.; Mäder, G.; Kriedt, R.A.; Stehmann, J.R.; Salzano, F.M.; Bonatto, S.L.; Freitas, L.B. Diversification of plant species in a subtropical region of eastern South American highlands: A phylogeographic perspective on native Petunia (Solanaceae). Mol. Ecol. 2010, 19, 5240–5251. [Google Scholar] [CrossRef] [PubMed]
- Kokubun, H.; Nakano, M.; Tsukamoto, T.; Watanabe, H.; Hashimoto, G.; Marchesi, E.; Bullrich, L.; Basualdo, I.L.; Kao, T.; Ando, T. Distribution of self-compatible and self-incompatible populations of Petunia axillaris (Solanaceae) outside Uruguay. J. Plant Res. 2006, 119, 419. [Google Scholar] [CrossRef]
- Reck-Kortmann, M.; Silva-Arias, G.A.; Segatto, A.L.; Mäder, G.; Bonatto, S.L.; de Freitas, L.B. Multilocus phylogeny reconstruction: New insights into the evolutionary history of the genus Petunia. Mol. Phylogenet. Evol. 2014, 81, 19–28. [Google Scholar] [CrossRef]
- Cheung, A.Y.; Wang, H.; Wu, H.-M. A floral transmitting tissue-specific glycoprotein attracts pollen tubes and stimulates their growth. Cell 1995, 82, 383–393. [Google Scholar] [CrossRef]
- Fernandez-Pozo, N.; Menda, N.; Edwards, J.D.; Saha, S.; Tecle, I.Y.; Strickler, S.R.; Bombarely, A.; Fisher-York, T.; Pujar, A.; Foerster, H.; et al. The Sol Genomics Network (SGN)—From genotype to phenotype to breeding. Nucleic Acids Res. 2015, 43, D1036–D1041. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, E.W.; Chase, M.W.; Knapp, S.; Litt, A.; Leitch, A.R.; Le Comber, S.C. Transgressive phenotypes and generalist pollination in the floral evolution of Nicotiana polyploids. Nat. Plants. 2016, 2, 16119. [Google Scholar] [CrossRef] [PubMed]
- Natarella, N.J.; Sink, K.C., Jr. Electrophoretic Analysis of Proteins and Peroxidases of Selected Petunia Species and Cultivars. Bot. Gazette 1975, 136, 20–26. [Google Scholar] [CrossRef]
- Chen, S.; Matsubara, K.; Omori, T.; Kokubun, H. Phylogenetic analysis of the genus Petunia (Solanaceae) based on the sequence of the Hf1 gene. J. Plant Res. 2007, 120, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Berenschot, A.S.; Quecini, V. A reverse genetics approach identifies novel mutants in light responses and anthocyanin metabolism in Petunia. Physiol. Mol. Biol. Plants 2014, 20, 1–13. [Google Scholar] [CrossRef][Green Version]
- Klahre, U.; Gurba, A.; Hermann, K.; Saxenhofer, M.; Bossolini, E.; Guerin, P.M.; Kuhlemeier, C. Pollinator Choice in Petunia Depends on Two Major Genetic Loci for Floral Scent Production. Curr. Biol. 2011, 21, 730–739. [Google Scholar] [CrossRef]
- Mulcahy, G.B.; Mulcahy, D.L. The two phases of growth of Petunia hybrida (Hort. Vilm-Andz.) pollen tubes through compatible styles. J. Palynol. 1982, 18, 61–64. [Google Scholar]
- McCray, F.A. Compatibility of Certain Nicotiana Species. Genetics 1932, 17, 621–636. [Google Scholar]
- Bedinger, P.A.; Chetelat, R.T.; McClure, B.; Moyle, L.C.; Rose, J.K.; Stack, S.M.; van der Knaap, E.; Baek, Y.S.; Lopez-Casado, G.; Covey, P.A.; et al. Interspecific reproductive barriers in the tomato clade: Opportunities to decipher mechanisms of reproductive isolation. Sex. Plant Reprod. 2010, 24, 171–187. [Google Scholar] [CrossRef]
- Rick, C.M. Hybridization between Lycopersicon esculentum and Solanum pennellii: Phylogenetic and cytogenetic significance. Proc. Natl. Sci. Acad. USA 1960, 46, 78–82. [Google Scholar] [CrossRef]
- Hogenboom, N.G. Incongruity: Non-Functioning of Intercellular and Intracellular Partner Relationships in Encyclopedia of Plant Physiology; Linskens, H.F., Heslop-Harrison, J., Eds.; Springer: Berlin, Germany, 1984; pp. 640–654. [Google Scholar]
- Park, M.; Park, J.; Kim, S.; Kwon, J.-K.; Park, H.M.; Bae, I.H.; Yang, T.-J.; Lee, Y.H.; Kang, B.-C. Evolution of the large genome in Capsicum annuum occurred through accumulation of single-type long terminal repeat retrotransposons and their derivatives. Plant J. 2012, 69, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, X.; Li, Z.; Zhu, Q.; Tang, W.; Ge, S.; Luo, J. Nucleotide substitution pattern in rice paralogues: Implication for negative correlation between the synonymous substitution rate and codon usage bias. Gene 2006, 376, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Gore, J.M.; Ran, F.A.; Ornston, L.N. Deletion Mutations Caused by DNA Strand Slippage in Acinetobacter baylyi. Appl. Environ. Microbiol. 2006, 72, 5239–5245. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.L.; Church, D.M.; Lash, A.E.; Leipe, D.D.; Madden, T.L.; Pontius, J.U.; Schuler, G.D.; Schriml, L.M.; Tatusova, T.A.; Wagner, L.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2002, 30, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Sánchez-DelBarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef] [PubMed]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486–487. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Self-Incompatibilibility (SI) or Self-Compatibility (SC) | Name of HD-AGP | Length in Basepairs (Number of Amino Acids) | Intron Length (bp) |
|---|---|---|---|---|
| Petunia axillaris axillaris | SC/SI | PaaPRP1 | 768 (256) | 1102 |
| Petunia axillaris parodii | SC | PapPRP1 | 768 (256) | 595 |
| Petunia exserta | SC | PePRP1 | 768 (256) | 593 |
| Petunia integrifolia | SI | PitPRP1 | 780 (260) | 604 |
| Petunia inflata | SI | PiPRP1 | 780 (260) | 601 |
| HD-AGP Sequence Attributes | ||||||||
|---|---|---|---|---|---|---|---|---|
| Species Pairs Compared | Name of HD-AGPs Compared | Reproductive Barrier | Overall Sequence Similarity: Nucleotide (Amino Acid) | Total # of Variant Residues | Total # of Variant Residues in HV1 | Total # of Variant Residues in HV2 | Total # of Variant Residues that Lie in XKPP Motifs | XKPP bias (# of Variant Residues in XKPP/Total # of Variant Residues X100) |
| P. axillaris axillaris vs. P. axillaris parodii | PaaPRP1 vs. PapPRP1 | None | 99.3% (99.6%) | 1 | 0 | 0 | 0 | 0 |
| P. axillaris axillaris vs. P. exserta | PaaPRP1 vs. PePRP1 | None | 99.5% (100%) | 0 | 0 | 0 | 0 | 0 |
| P. axillaris axillaris vs. P. integrifolia | PaaPRP1 vs. PitPRP1 | Bilateral IRB | 97.6% (97.3%) | 7 | 0 | 6 | 4 | 57% |
| P. axillaris axillaris vs. P. inflata | PaaPRP1 vs. PiPRP1 | Bilateral IRB | 97% (96.9%) | 8 | 0 | 6 | 4 | 50% |
| P. integrifolia vs. P. inflata | PitPRP1 vs. PiPRP1 | None | 99.23% (98.9%) | 3 | 0 | 0 | 0 | 0 |
| Species Compared | Reproductive Barrier | Sequence 1 | Sequence 2 | XKPP Indels in HV2 | Ka/Ks Ratio | |||
|---|---|---|---|---|---|---|---|---|
| HV1 | His | HV2 | PAC | |||||
| P. axillaris axillaris vs. P axillaris parodii | None | PaaPRP1 | PapPRP1 | 0 | 0.0000 | 0.0000 | 0.0000 | 0.6730 |
| P. axillaris axillaris vs. P. exserta | None | PaaPRP1 | PePRP1 | 0 | 0.0000 | 0.0000 | 0.0000 | Ka-Ks > 0 |
| P. axillaris axillaris vs. P. integrifolia | SI-related IRB + incongruity | PaaPRP1 | PitPRP1 | 1 | 0.0000 | 0.0000 | Ka-Ks > 0 | 0.2210 |
| P. axillaris axillaris vs. P. inflata | SI-related IRB + incongruity | PaaPRP1 | PiPRP1 | 1 | 0.0000 | 0.0000 | Ka-Ks > 0 | 0.9946 |
| P. axillaris parodii vs. P. exserta | None | PapPRP1 | PePRP1 | 0 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| P. axillaris parodii vs. P. integrifolia | SI-related IRB + incongruity | PapPRP1 | PitPRP1 | 1 | 0.0000 | 0.0000 | Ka-Ks > 0 | 0.2346 |
| P. axillaris parodii vs. P. inflata | SI-related IRB + incongruity | PapPRP1 | PiPRP1 | 1 | 0.0000 | 0.0000 | Ka-Ks > 0 | 0.4373 |
| P. exserta vs. P. integrifolia | SI-related IRB + incongruity | PePRP1 | PitPRP1 | 1 | 0.0000 | 0.0000 | Ka-Ks > 0 | 0.3304 |
| P. exserta vs. P. inflata | SI-related IRB + incongruity | PePRP1 | PiPRP1 | 1 | 0.0000 | 0.0000 | Ka-Ks > 0 | 0.6595 |
| P. integrifolia vs. P. inflata | None | PitPRP1 | PiPRP1 | 0 | 0.0000 | 0.0000 | 0.0000 | 0.5931 |
| Species | Name of HD-AGPs Compared | Section, Haploid Number | SI: Self-Incompatible; SC: Self-Compatible | Breeding Behavior | Length of HD-AGP (# of Amino Acids) | Total # of XKPP Motifs |
|---|---|---|---|---|---|---|
| Nicotiana alata | NaPRP4 | Alatae, n = 9 | Mostly SI | Rejects most heterospecific pollen via SI | 255 | 7 |
| Nicotiana sylvestris | NsPRP1 = TTS1 | Sylvestres, n = 12 | SC | N. alata or N. paniculata pollen set seed | 257 | 7 |
| Nicotiana paniculata | NpPRP1 | Paniculatae, n = 12 | Mostly SC | N.alata or N. sylvestris pollen set seed | 255 | 8 |
| Nicotiana tomentosiformis | NtPRP1 = TTS2 | Tomentosae, n = 12 | SC | Hybridizes with N. obtusifolia, but IRB with other 3 | 261 | 9 |
| Nicotiana obtusifolia | NobPRP1 | Trigonophyllae N = 12 | SC | Hybridizes with N. tomentosiformis, but IRB with other 3 | 263 | 9 |
| Species Compared | Seq 1 | Seq 2 | Reproductive Barrier | Percent Identity: Nucleotide (Amino Acid) | Number of XKPP Variants |
|---|---|---|---|---|---|
| N. alata vs. N. sylvestris | NaPRP4 | NsPRP1 | SI-related IRB + incongruity | 95 (96) | 2 |
| N. alata vs. N. paniculata | NaPRP4 | NpPRP1 | None | 94 (95) | 1 |
| N. alata vs. N. tomentosiformis | NaPRP4 | NtoPRP1 | SI-related IRB + incongruity | 91 (92) | 1 |
| N. alata vs. N.obtusifolia | NaPRP4 | NobPRP1 | SI-related IRB + incongruity | 90 (92) | 1 |
| N. sylvestris vs. N. paniculata | NsPRP1 | NpPRP1 | None | 94 (93) | 0 |
| N. sylvestris vs. N. tomentosiformis | NsPRP1 | NtoPRP1 | Bilateral incongruity | 89 (88) | 1 |
| N. sylvestris vs. N.obtusifolia | NsPRP1 | NobPRP1 | Bilateral incongruity | 88 (93) | 1 |
| N. paniculata vs. N. tomentosiformis | NpPRP1 | NtoPRP1 | Bilateral incongruity | 92 (95) | 1 |
| N. paniculata vs. N.obtusifolia | NpPRP1 | NobPRP1 | Bilateral incongruity | 94 (94) | 1 |
| N. tomentosiformis vs. N.obtusifolia | NtoPRP1 | NobPRP1 | None | 95 (95) | 0 |
| Species Compared (SI/SC) | Seq 1 | Seq 2 | Reproductive Barrier | Percent Identity: Nucleotide (Amino Acid) | XKPP Indels (Number) |
|---|---|---|---|---|---|
| S. lycopersicum (SC) vs. S. pimpinellifolium (SC) | SlPRP1 | SppPRP1 | None | 99.9 (100) | 0 |
| S. lycopersicum (SC) vs. S. pennellii (SI) | SlPRP1 | SpPRP1 | SI-related IRB + incongruity? | 97.1 (97.6) | 1 |
| S. lycopersicum (SC) vs. S. lycopersicoides (SI) | SlPRP4 | SlyPRP1 | SI-related IRB + incongruity | 94.4 (94.5) | 2 |
| S. pimpinellifolium (SC) vs. S. pennellii (SI) | SppPRP1 | SpPRP1 | SI-related IRB + incongruity? | 97.1 (97.6) | 1 |
| S. pimpinellifolium (SC) vs. S. lycopersicoides (SI) | SppPRP1 | SlyPRP1 | SI-related IRB + incongruity | 94.5 (94.5) | 2 |
| S. pennellii (SI) vs. S. lycopersicoides (SI) | SpPRP1 | SlyPRP1 | Bilateral IRB: presumed SI-related | 95.6 (96.4) | 2 |
| 8 | Ka/Ks Ratio | ||||||
|---|---|---|---|---|---|---|---|
| Species Compared | Seq 1 | Seq 2 | Percent Identity: Nucleotide (Amino Acid) | HV1 | His | HV2 | PAC |
| Petunia axillaris vs. Nicotiana alata | PaPRP1 | NaPRP4 | 83 (72) | 1.5007 | 0.1556 | 0.7213 | 0.3914 |
| Petunia axillaris vs. Capsicum annuum | PaPRP1 | CaPRP1 | 78 (75) | 1.6617 | 0.1009 | 1.1805 | 0.3419 |
| Petunia axillaris vs. Solanum lycopersicum | PaPRP1 | SlPRP1 | 75 (65) | 0.8780 | 0.0972 | 1.6127 | 0.2808 |
| Petunia axillaris vs. Solanum tuberosum | PaPRP1 | StuPRP1 | 77 (67) | 1.6635 | 0.0972 | 1.7705 | 0.2540 |
| Nicotiana alata vs. Capsicum annuum | NaPRP4 | CaPRP1 | 80 (79) | 1.9018 | 0.2060 | 0.4827 | 0.2924 |
| Nicotiana alata vs. Solanum lycopersicum | NaPRP4 | SlPRP1 | 77 (69) | 1.1220 | 0.0972 | 0.1643 | 0.1971 |
| Nicotiana alata vs. Solanum tuberosum | NaPRP4 | StuPRP1 | 78 (71) | 2.2642 | 0.0972 | 0.3550 | 0.2039 |
| Capsicum annuum vs. Solanum tuberosum | CaPRP1 | StuPRP1 | 79 (71) | Ka-Ks > 0 | 0.3292 | 1.1589 | 0.2091 |
| Capsicum annuum vs. Solanum lycopersicum | CaPRP1 | SlPRP1 | 73 (64) | 0.06875 | 0.3292 | 0.7712 | 0.2716 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Callaway, T.D.; Singh-Cundy, A. HD-AGPs as Speciation Genes: Positive Selection on a Proline-Rich Domain in Non-Hybridizing Species of Petunia, Solanum, and Nicotiana. Plants 2019, 8, 211. https://doi.org/10.3390/plants8070211
Callaway TD, Singh-Cundy A. HD-AGPs as Speciation Genes: Positive Selection on a Proline-Rich Domain in Non-Hybridizing Species of Petunia, Solanum, and Nicotiana. Plants. 2019; 8(7):211. https://doi.org/10.3390/plants8070211
Chicago/Turabian StyleCallaway, Tara D., and Anu Singh-Cundy. 2019. "HD-AGPs as Speciation Genes: Positive Selection on a Proline-Rich Domain in Non-Hybridizing Species of Petunia, Solanum, and Nicotiana" Plants 8, no. 7: 211. https://doi.org/10.3390/plants8070211
APA StyleCallaway, T. D., & Singh-Cundy, A. (2019). HD-AGPs as Speciation Genes: Positive Selection on a Proline-Rich Domain in Non-Hybridizing Species of Petunia, Solanum, and Nicotiana. Plants, 8(7), 211. https://doi.org/10.3390/plants8070211