The Dynamics of Cytokinin Changes after Grafting of Vegetative Apices on Flowering Rapeseed Plants

 ,
, 
 and
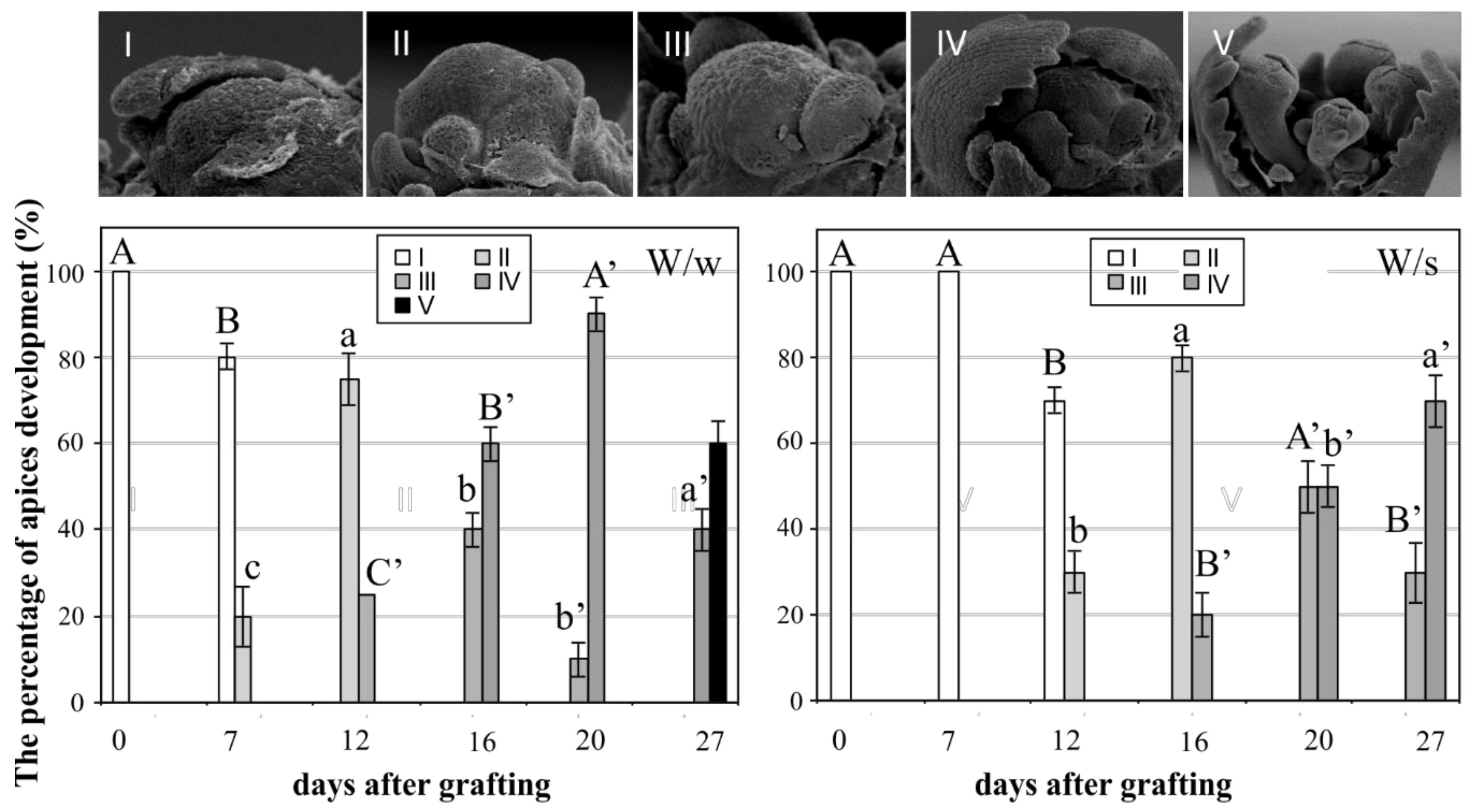
and 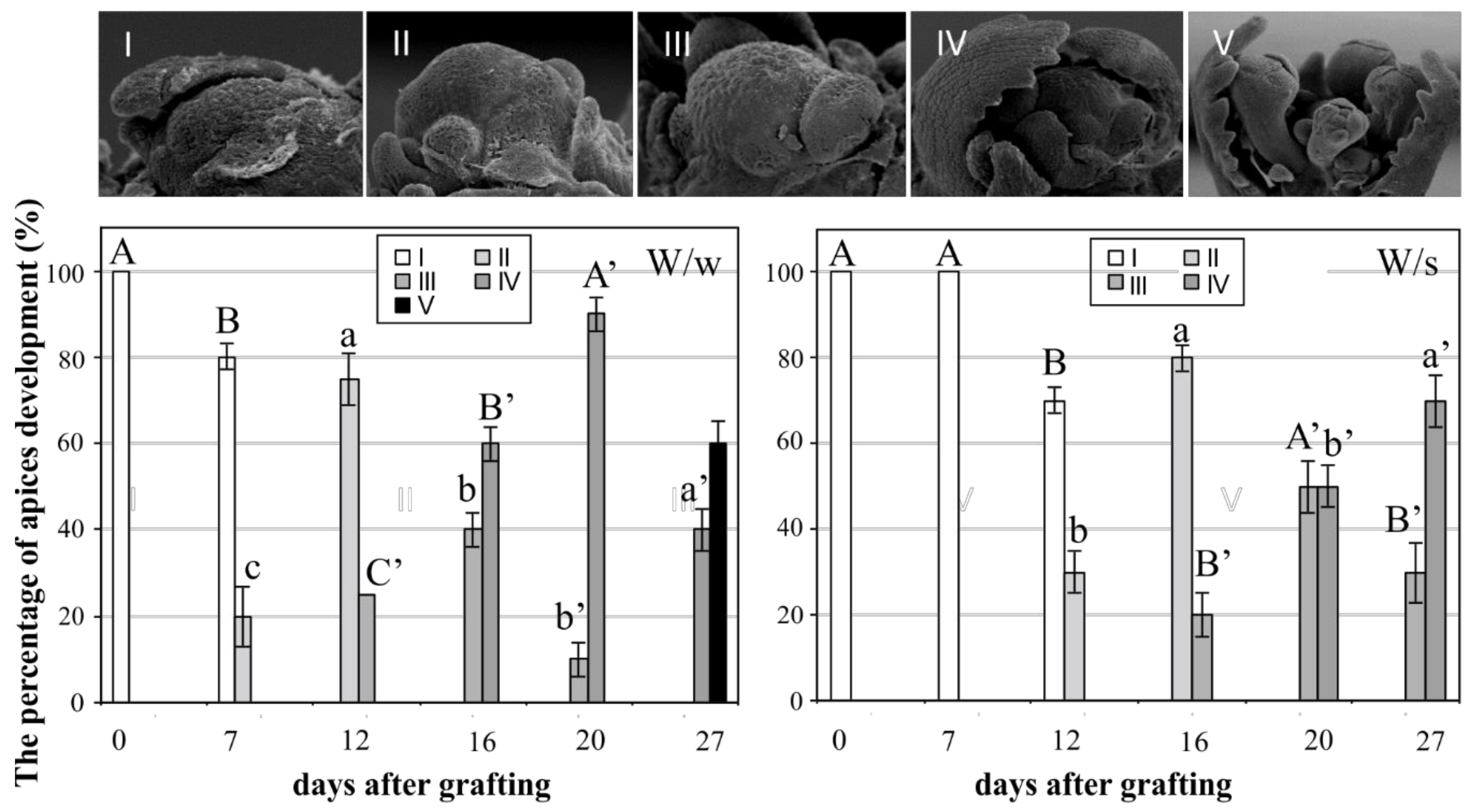{kind=link}
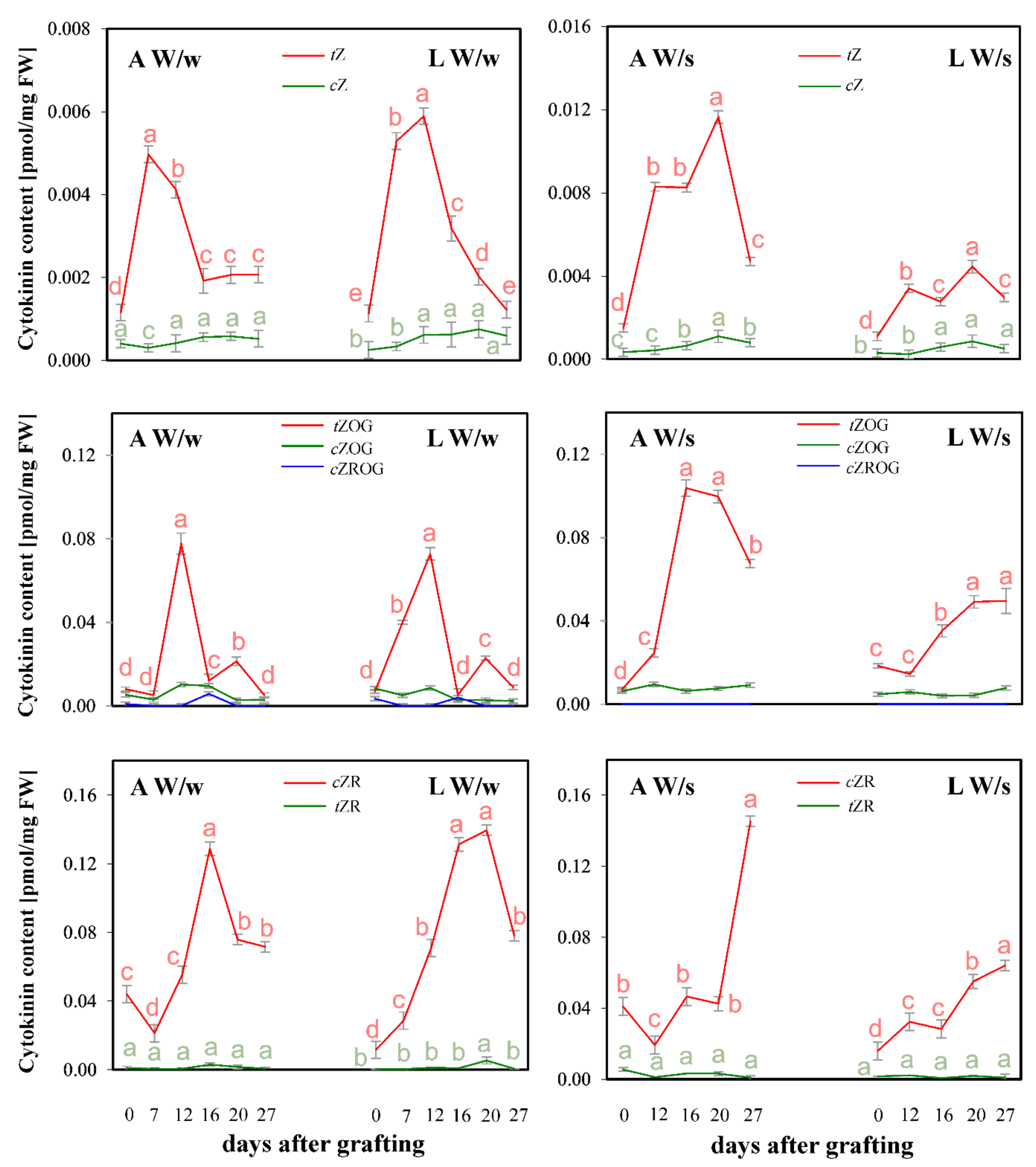{kind=link}
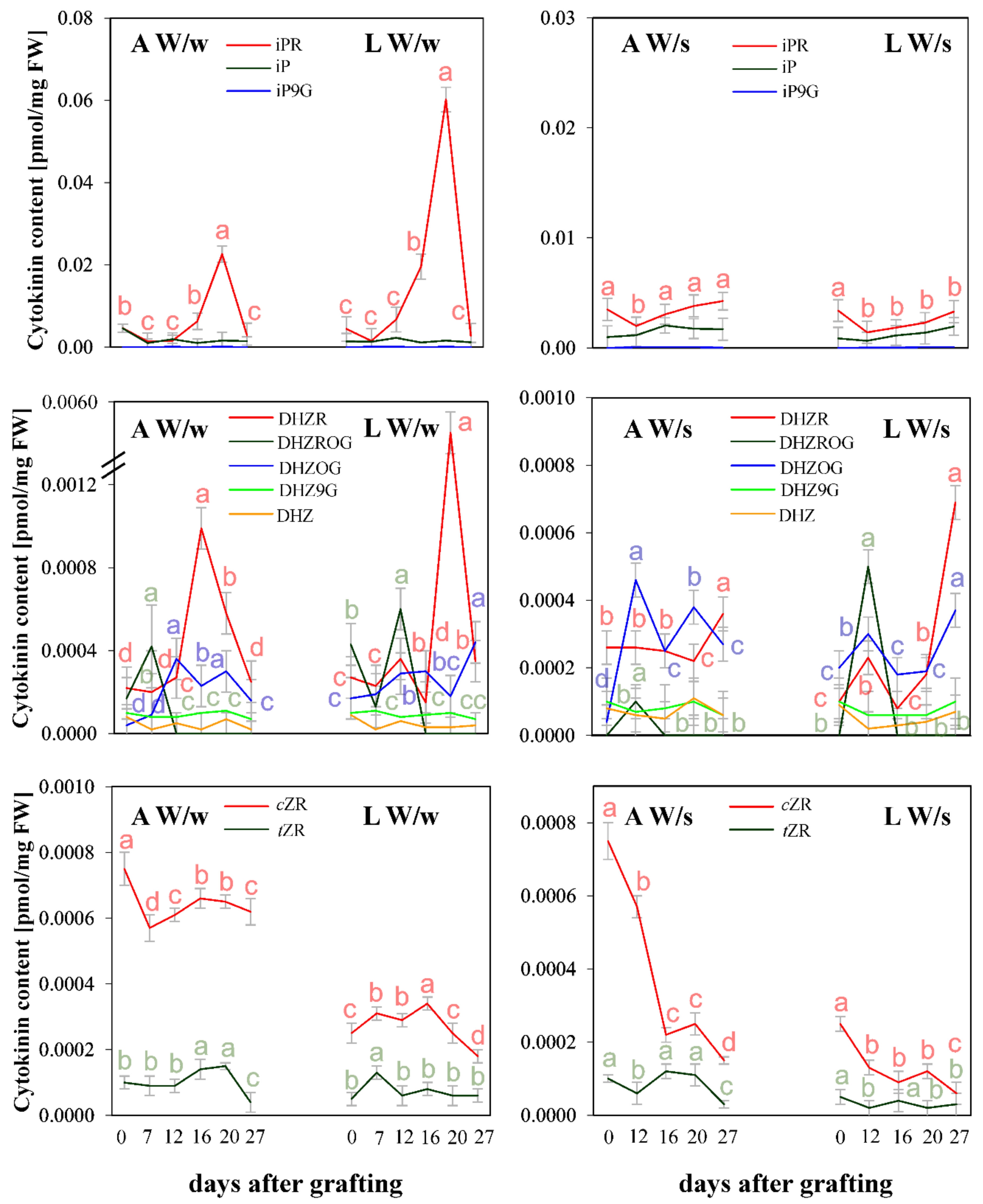{kind=link}
Abstract
:1. Introduction
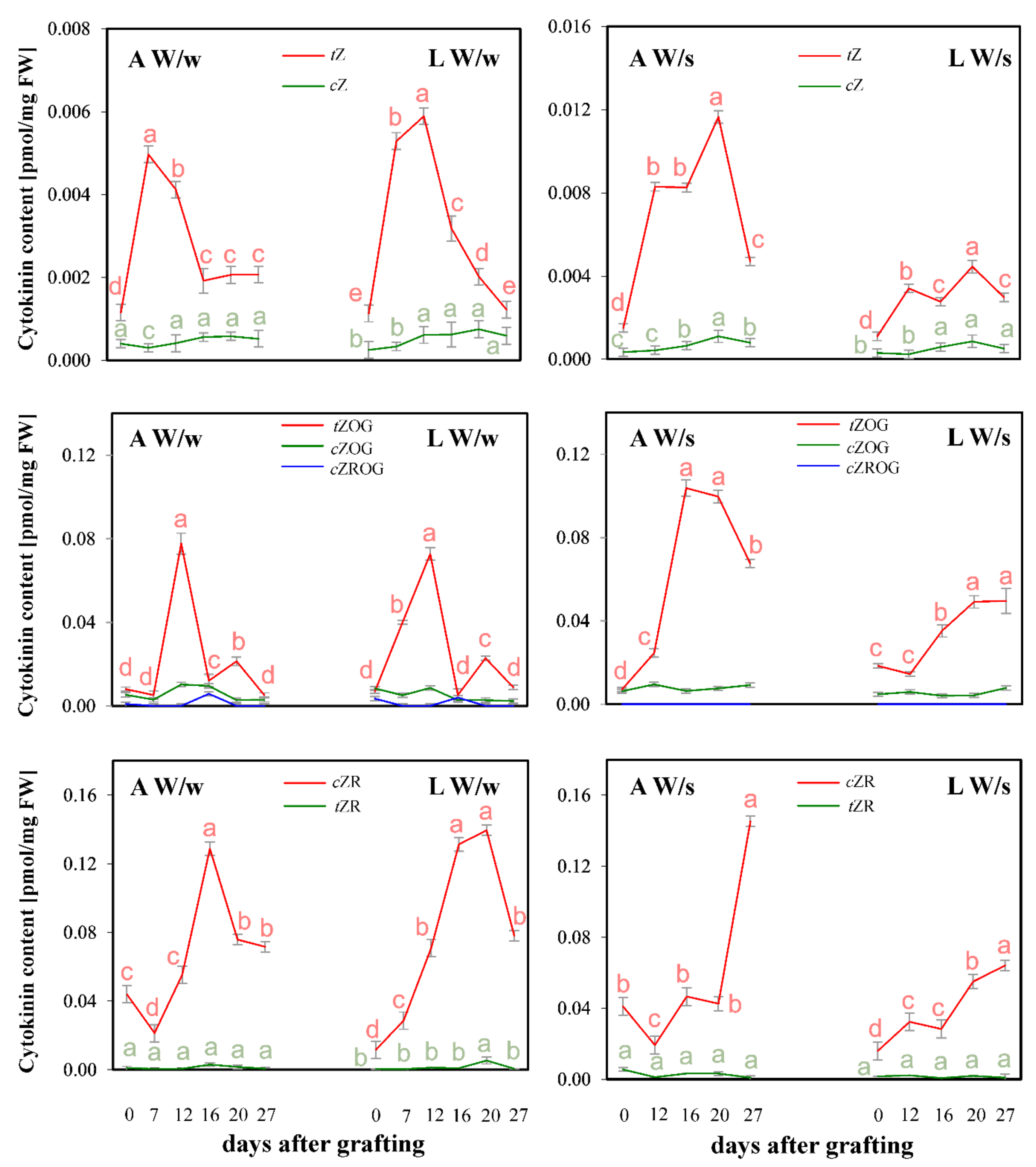
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material
3.2. Cytokinin Analysis by Ultra-High-Performance Liquid Chromatography-Electrospray Tandem Mass Spectrometry (UHPLC–ESI–MS/MS)
Author Contributions
Funding
Conflicts of Interest
References
- Poethig, R.S. Phase change and the regulation of developmental timing in plants. Science 2003, 301, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Doerner, P. Plant meristems: A merry-go-round of signals. Curr. Biol. 2003, 13, R368–R374. [Google Scholar] [CrossRef]
- Veit, B. Hormone mediated regulation of the shoot apical meristem. Plant Mol. Biol. 2009, 69, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef] [PubMed]
- Amasino, R. Seasonal and developmental timing of flowering. Plant J. 2010, 61, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef]
- Bernier, G.; Kinet, J.-M.; Sachs, R.M. The Physiology of Flowering, Volume II. Transition to Reproductive Growth; CRC Press: Boca Raton, FL, USA, 1981; pp. 97–104. [Google Scholar]
- Bernier, G.; Corbesier, L.; Perilleux, C. The flowering process: On the track of controlling factors in Sinapis alba. Russ. J. Plant Physiol. 2002, 49, 445–450. [Google Scholar] [CrossRef]
- Jacqmard, A.; Detry, N.; Dewitte, W.; van Onckelen, H.; Bernier, G. In situ localisation of cytokinins in the shoot apical meristem of Sinapis alba at floral transition. Planta 2002, 214, 970–973. [Google Scholar] [CrossRef] [PubMed]
- Corbesier, L.; Prinsen, E.; Jacqmard, A.; Lejeune, P.; Van Onckelen, H.; Périlleux, C.; Bernier, G. Cytokinin levels in leaves, leaf exudate and shoot apical meristem of Arabidopsis thaliana during floral transition. J. Exp. Bot. 2003, 54, 2511–2517. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; Chickarmane, V.S.; Ohno, C.; Meyerowitz, E.M. Multiple feedback loops through cytokinin signaling control stem cell number within the Arabidopsis shoot meristem. Proc. Natl. Acad. Sci. USA 2009, 106, 16529–16534. [Google Scholar] [CrossRef]
- Ormenese, S.; Bernier, G.; Périlleux, C. Cytokinin application to the shoot apical meristem of Sinapis alba enhances secondary plasmodesmata formation. Planta 2006, 224, 1481–1484. [Google Scholar] [CrossRef] [PubMed]
- D’Aloia, M.; Bonhomme, D.; Bouché, F.; Tamseddak, K.; Ormenese, S.; Torti, S.; Coupland, G.; Périlleux, C. Cytokinin promotes flowering of Arabidopsis via transcriptional activation of the FT paralogue TSF. Plant J. 2011, 65, 972–979. [Google Scholar] [CrossRef]
- Bernier, G. My favourite flowering image: The role of cytokinin as a flowering signal. J. Exp. Bot. 2013, 64, 5795–5799. [Google Scholar] [CrossRef] [PubMed]
- Tarkowská, D.; Filek, M.; Biesaga-Kościelniak, J.; Marciñska, I.; Macháčková, I.; Krekule, J.; Strnad, M. Cytokinins in shoot apices of Brassica napus plants during vernalization. Plant Sci. 2012, 187, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Gajdošová, S.; Spíchal, L.; Kamínek, M.; Hoyerová, K.; Novák, O.; Dobrev, P.I.; Galuszka, P.; Klíma, P.; Gaudinová, A.; Žižková, E.; et al. Distribution, biological activities, metabolism, and the conceivable function of cis-zeatin-type cytokinins in plants. J. Exp. Bot. 2011, 62, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Schmülling, T. New insights into the functions of cytokinins in plant development. J. Plant Growth Regul. 2002, 21, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Köllmer, I.; Bartrina, I.; Manns, I.; Holst, K.; Schmülling, T. New insights into the biology of cytokinin degradation. Plant Biol. 2006, 8, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Ayre, B.G.; Turgeon, R. Graft transmission of a floral stimulant derived from CONSTANS. Plant Physiol. 2004, 135, 2271–2278. [Google Scholar] [CrossRef] [PubMed]
- Filek, M.; Biesaga-Koścelniak, J.; Marcińska, I.; Krekule, J.; Macháčková, I.; Dubert, F. The effects of electric current on flowering of grafted scions of non-vernalized winter rape. Biol. Plant. 2003, 46, 625–628. [Google Scholar] [CrossRef]
- Filek, M.; Biesaga-Kosćielniak, J.; Marcińska, I.; Macháčková, I.; Dubert, F. Electric current affects the rate of development in isolated apical parts of rape in vitro. Biol. Plant. 2006, 50, 465–468. [Google Scholar] [CrossRef]
- Bieleski, R.L. The problem of halting enzyme action when extracting plant tissues. Anal. Biochem. 1964, 9, 431–442. [Google Scholar] [CrossRef]
- Faiss, M.; Zalubilova, J.; Strnad, M.; Schmülling, T. Conditional transgenic expression of the ipt gene indicates a function for cytokinins in paracrine signaling in whole tobacco plants. Plant J. 1997, 12, 401–415. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarkowská, D.; Filek, M.; Krekule, J.; Biesaga-Kościelniak, J.; Marciñska, I.; Popielarska-Konieczna, M.; Strnad, M. The Dynamics of Cytokinin Changes after Grafting of Vegetative Apices on Flowering Rapeseed Plants. Plants 2019, 8, 78. https://doi.org/10.3390/plants8040078
Tarkowská D, Filek M, Krekule J, Biesaga-Kościelniak J, Marciñska I, Popielarska-Konieczna M, Strnad M. The Dynamics of Cytokinin Changes after Grafting of Vegetative Apices on Flowering Rapeseed Plants. Plants. 2019; 8(4):78. https://doi.org/10.3390/plants8040078
Chicago/Turabian StyleTarkowská, Danuše, Maria Filek, Jan Krekule, Jolanta Biesaga-Kościelniak, Izabela Marciñska, Marzena Popielarska-Konieczna, and Miroslav Strnad. 2019. "The Dynamics of Cytokinin Changes after Grafting of Vegetative Apices on Flowering Rapeseed Plants" Plants 8, no. 4: 78. https://doi.org/10.3390/plants8040078
APA StyleTarkowská, D., Filek, M., Krekule, J., Biesaga-Kościelniak, J., Marciñska, I., Popielarska-Konieczna, M., & Strnad, M. (2019). The Dynamics of Cytokinin Changes after Grafting of Vegetative Apices on Flowering Rapeseed Plants. Plants, 8(4), 78. https://doi.org/10.3390/plants8040078