Plant-Produced Anti-Enterovirus 71 (EV71) Monoclonal Antibody Efficiently Protects Mice Against EV71 Infection

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
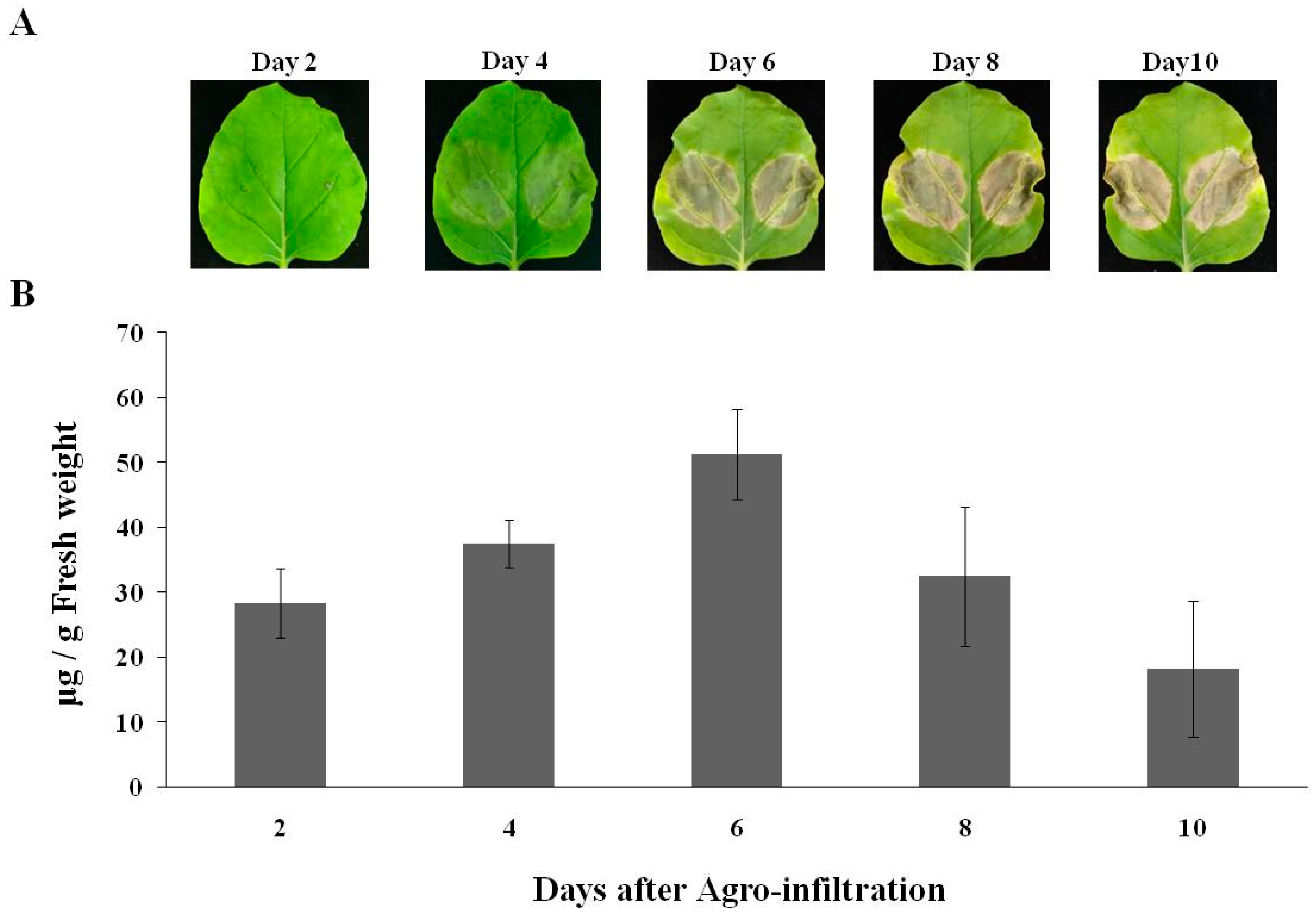
2.1. Expression of cD5 mAb in Nicotiana benthamiana
2.2. Purification of cD5 mAb from N. benthamiana Leaves
2.3. Plant-Produced cD5 mAb Retains Antigen-Binding Activity
2.4. Plant-Produced cD5 mAb Efficiently Neutralized EV71
2.5. Plant-Produced cD5 mAb Protected Recipient Mice Against Lethal EV71 Infection
3. Discussion
4. Materials and Methods
4.1. Gene Construct
4.2. Protein Expression, Extraction and Quantification
4.3. Purification
4.4. SDS-PAGE and Western Blot Analysis
4.5. Binding Assay
4.6. Neutralization Assay
4.7. In Vivo Protection Assay
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, Y.; Badaruddin, H.; Lee, V.J.; Cutter, J.; Cook, A.R. The effect of school closure on hand, foot, and mouth disease transmission in Singapore: A Modeling Approach. Am. J. Trop. Med. Hyg. 2018, 99, 1625–1632. [Google Scholar] [CrossRef]
- Hoang, M.T.V.; Nguyen, T.A.; Tran, T.T.; Vu, T.T.H.; Le, N.T.N.; Nguyen, T.H.N.; Le, T.H.N.; Nguyen, T.T.H.; Nguyen, T.H.; Le, N.T.N.; et al. Clinical and aetiological study of hand, foot and mouth disease in southern Vietnam, 2013-2015: Inpatients and outpatients. Int. J. Infect. Dis. 2019, 80, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ooi, M.H.; Wong, S.C.; Lewthwaite, P.; Cardosa, M.J.; Solomon, T. Clinical features, diagnosis, and management of enterovirus 71. Lancet Neurol. 2010, 9, 1097–1105. [Google Scholar] [CrossRef]
- Huang, C.C.; Liu, C.C.; Chang, Y.C.; Chen, C.Y.; Wang, S.T.; Yeh, T.F. Neurologic complications in children with enterovirus 71 infection. N. Engl. J. Med. 1999, 341, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Wang, J.; Liu, L.; Zhao, H.; Shi, H.; Zhang, Y.; Jiang, L.; Li, Q. Optimized development of a candidate strain of inactivated EV71 vaccine and analysis of its immunogenicity in rhesus monkeys. Hum. Vaccin. 2010, 6, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, Y.; Wang, J.; Zhao, H.; Jiang, L.; Che, Y.; Shi, H.; Li, R.; Mo, Z.; Huang, T.; et al. Study of the integrated immune response induced by an inactivated EV71 vaccine. PLoS ONE 2013, 8, e54451. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Liu, L.; Zhao, H.; Wang, J.; Liao, Y.; Zhang, X.; Na, R.; Liang, Y.; Wang, L.; Li, Q. Immunoprotection elicited by an enterovirus type 71 experimental inactivated vaccine in mice and rhesus monkeys. Vaccine 2011, 29, 6269–6275. [Google Scholar] [CrossRef] [PubMed]
- Bek, E.J.; Hussain, K.M.; Phuektes, P.; Kok, C.C.; Gao, Q.; Cai, F.; Gao, Z.; McMinn, P.C. Formalin-inactivated vaccine provokes cross-protective immunity in a mouse model of human enterovirus 71 infection. Vaccine 2011, 29, 4829–4838. [Google Scholar] [CrossRef]
- Cai, Y.; Ku, Z.; Liu, Q.; Leng, Q.; Huang, Z. A combination vaccine comprising of inactivated enterovirus 71 and coxsackievirus A16 elicits balanced protective immunity against both viruses. Vaccine 2014, 32, 2406–2412. [Google Scholar] [CrossRef]
- Chiu, C.H.; Chu, C.; He, C.C.; Lin, T.Y. Protection of neonatal mice from lethal enterovirus 71 infection by maternal immunization with attenuated Salmonella enterica serovar Typhimurium expressing VP1 of enterovirus 71. Microbes Infect. 2006, 8, 1671–1678. [Google Scholar] [CrossRef]
- Arita, M.; Nagata, N.; Iwata, N.; Ami, Y.; Suzaki, Y.; Mizuta, K.; Iwasaki, T.; Sata, T.; Wakita, T.; Shimizu, H. An attenuated strain of enterovirus 71 belonging to genotype a showed a broad spectrum of antigenicity with attenuated neurovirulence in cynomolgus monkeys. J. Virol. 2007, 81, 9386–9395. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Ho, M.S.; Wu, J.C.; Chen, W.J.; Huang, J.H.; Chou, S.T.; Hu, Y.C. Immunization with virus-like particles of enterovirus 71 elicits potent immune responses and protects mice against lethal challenge. Vaccine 2008, 26, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.N.; Lin, Y.C.; Fann, C.; Liao, N.S.; Shih, S.R.; Ho, M.S. Protection against lethal enterovirus 71 infection in newborn mice by passive immunization with subunit VP1 vaccines and inactivated virus. Vaccine 2001, 20, 895–904. [Google Scholar] [CrossRef]
- Chung, C.Y.; Chen, C.Y.; Lin, S.Y.; Chung, Y.C.; Chiu, H.Y.; Chi, W.K.; Lin, Y.L.; Chiang, B.L.; Chen, W.J.; Hu, Y.C. Enterovirus 71 virus-like particle vaccine: Improved production conditions for enhanced yield. Vaccine 2010, 28, 6951–6957. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ku, Z.; Dai, W.; Chen, T.; Ye, X.; Zhang, C.; Zhang, Y.; Liu, Q.; Jin, X.; Huang, Z. A bivalent virus-like particle based vaccine induces a balanced antibody response against both enterovirus 71 and norovirus in mice. Vaccine 2015, 33, 5779–5785. [Google Scholar] [CrossRef]
- Foo, D.G.; Alonso, S.; Chow, V.T.; Poh, C.L. Passive protection against lethal enterovirus 71 infection in newborn mice by neutralizing antibodies elicited by a synthetic peptide. Microbes Infect. 2007, 9, 1299–1306. [Google Scholar] [CrossRef]
- Liu, J.N.; Wang, W.; Duo, J.Y.; Hao, Y.; Ma, C.M.; Li, W.B.; Lin, S.Z.; Gao, X.Z.; Liu, X.L.; Xu, Y.F.; et al. Combined peptides of human enterovirus 71 protect against virus infection in mice. Vaccine 2010, 28, 7444–7451. [Google Scholar] [CrossRef]
- Cao, R.; Han, J.; Qin, E.; Qin, C. [Mechanism of intravenous immunoglobulin therapy for severe hand-foot-mouth disease: A review]. Sheng Wu Gong Cheng Xue Bao 2011, 27, 712–716. [Google Scholar]
- Elgundi, Z.; Reslan, M.; Cruz, E.; Sifniotis, V.; Kayser, V. The state-of-play and future of antibody therapeutics. Adv. Drug Deliv. Rev. 2017, 122, 2–19. [Google Scholar] [CrossRef]
- Lim, X.F.; Jia, Q.; Khong, W.X.; Yan, B.; Premanand, B.; Alonso, S.; Chow, V.T.; Kwang, J. Characterization of an isotype-dependent monoclonal antibody against linear neutralizing epitope effective for prophylaxis of enterovirus 71 infection. PLoS ONE 2012, 7, e29751. [Google Scholar] [CrossRef]
- Kiener, T.K.; Jia, Q.; Meng, T.; Chow, V.T.; Kwang, J. A novel universal neutralizing monoclonal antibody against enterovirus 71 that targets the highly conserved “knob” region of VP3 protein. PLoS Negl. Trop. Dis. 2014, 8, e2895. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.H.; Luo, Y.J.; Wu, X.Y.; Si, B.Y.; Lin, L.; Zhu, Q.Y. Monoclonal antibody induced with inactived EV71-Hn2 virus protects mice against lethal EV71-Hn2 virus infection. Virol. J. 2010, 7, 106. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Chou, A.H.; Lien, S.P.; Lin, H.Y.; Liu, S.J.; Chang, J.Y.; Guo, M.S.; Chow, Y.H.; Yang, W.S.; Chang, K.H.; et al. Identification and characterization of a cross-neutralization epitope of Enterovirus 71. Vaccine 2011, 29, 4362–4372. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.Q.; Ma, J.; Xu, L.J.; Li, Y.X.; Zhao, H.; Han, J.F.; Tao, J.; Li, X.F.; Zhu, S.Y.; Qin, E.D.; et al. Generation and characterization of a protective mouse monoclonal antibody against human enterovirus 71. Appl. Microbiol. Biotechnol. 2015, 99, 7663–7671. [Google Scholar] [CrossRef]
- Lin, T.; Xianyu, L.; Lyu, S. Monoclonal neutralizing antibodies against EV71 screened from mice immunized with yeast-produced virus-like particles. Virol. Sin. 2015, 30, 208–213. [Google Scholar] [CrossRef]
- Ku, Z.; Ye, X.; Shi, J.; Wang, X.; Liu, Q.; Huang, Z. Single Neutralizing Monoclonal Antibodies Targeting the VP1 GH Loop of Enterovirus 71 Inhibit both Virus Attachment and Internalization during Viral Entry. J. Virol. 2015, 89, 12084–12095. [Google Scholar] [CrossRef]
- Ye, X.; Fan, C.; Ku, Z.; Zuo, T.; Kong, L.; Zhang, C.; Shi, J.; Liu, Q.; Chen, T.; Zhang, Y.; et al. Structural Basis for Recognition of Human Enterovirus 71 by a Bivalent Broadly Neutralizing Monoclonal Antibody. PLoS Pathog. 2016, 12, e1005454. [Google Scholar] [CrossRef]
- Chen, Q.; Davis, K.R. The potential of plants as a system for the development and production of human biologics. F1000Reserch 2016, 5. [Google Scholar] [CrossRef]
- Lai, H.; Engle, M.; Fuchs, A.; Keller, T.; Johnson, S.; Gorlatov, S.; Diamond, M.S.; Chen, Q. Monoclonal antibody produced in plants efficiently treats West Nile virus infection in mice. Proc. Natl. Acad. Sci. USA 2010, 107, 2419–2424. [Google Scholar] [CrossRef]
- Huang, Z.; Phoolcharoen, W.; Lai, H.; Piensook, K.; Cardineau, G.; Zeitlin, L.; Whaley, K.J.; Arntzen, C.J.; Mason, H.S.; Chen, Q. High-level rapid production of full-size monoclonal antibodies in plants by a single-vector DNA replicon system. Biotechnol. Bioeng. 2010, 106, 9–17. [Google Scholar] [CrossRef]
- Both, L.; van Dolleweerd, C.; Wright, E.; Banyard, A.C.; Bulmer-Thomas, B.; Selden, D.; Altmann, F.; Fooks, A.R.; Ma, J.K. Production, characterization, and antigen specificity of recombinant 62-71-3, a candidate monoclonal antibody for rabies prophylaxis in humans. FASEB J. 2013, 27, 2055–2065. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.Y.; Wang, Y.; Bian, L.; Xu, M.; Liang, Z. EV71 vaccine, a new tool to control outbreaks of hand, foot and mouth disease (HFMD). Expert Rev. Vaccines 2016, 15, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Templar, A.; Marsh, D.; Nesbeth, D.N. A synthetic biology standard for Chinese Hamster Ovary cell genome monitoring and contaminant detection by polymerase chain reaction. Springerplus 2016, 5, 1510. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q. Plant-made vaccines against West Nile virus are potent, safe, and economically feasible. Biotechnol. J. 2015, 10, 671–680. [Google Scholar] [CrossRef]
- Nandi, S.; Kwong, A.T.; Holtz, B.R.; Erwin, R.L.; Marcel, S.; McDonald, K.A. Techno-economic analysis of a transient plant-based platform for monoclonal antibody production. MAbs 2016, 8, 1456–1466. [Google Scholar] [CrossRef]
- Chen, Q.; Lai, H.; Hurtado, J.; Stahnke, J.; Leuzinger, K.; Dent, M. Agroinfiltration as an Effective and Scalable Strategy of Gene Delivery for Production of Pharmaceutical Proteins. Adv. Tech. Biol. Med. 2013, 1. [Google Scholar] [CrossRef]
- Weintraub, J.A.; Hilton, J.F.; White, J.M.; Hoover, C.I.; Wycoff, K.L.; Yu, L.; Larrick, J.W.; Featherstone, J.D. Clinical trial of a plant-derived antibody on recolonization of mutans streptococci. Caries Res. 2005, 39, 241–250. [Google Scholar] [CrossRef]
- Ma, J.K.; Drossard, J.; Lewis, D.; Altmann, F.; Boyle, J.; Christou, P.; Cole, T.; Dale, P.; van Dolleweerd, C.J.; Isitt, V.; et al. Regulatory approval and a first-in-human phase I clinical trial of a monoclonal antibody produced in transgenic tobacco plants. Plant Biotechnol. J. 2015, 13, 1106–1120. [Google Scholar] [CrossRef]
- Tuse, D.; Ku, N.; Bendandi, M.; Becerra, C.; Collins, R., Jr.; Langford, N.; Sancho, S.I.; Lopez-Diaz de Cerio, A.; Pastor, F.; Kandzia, R.; et al. Clinical Safety and Immunogenicity of Tumor-Targeted, Plant-Made Id-KLH Conjugate Vaccines for Follicular Lymphoma. Biomed. Res. Int. 2015, 2015, 648143. [Google Scholar] [CrossRef]
- Li, Y.; Yu, J.; Qi, X.; Yan, H. Monoclonal antibody against EV71 3D(pol) inhibits the polymerase activity of RdRp and virus replication. BMC Immunol. 2019, 20, 6. [Google Scholar] [CrossRef]
- Thanongsaksrikul, J.; Srimanote, P.; Tongtawe, P.; Glab-Ampai, K.; Malik, A.A.; Supasorn, O.; Chiawwit, P.; Poovorawan, Y.; Chaicumpa, W. Identification and production of mouse scFv to specific epitope of enterovirus-71 virion protein-2 (VP2). Arch. Virol. 2018, 163, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Ng, Q.; Chin, W.; Meng, T.; Chow, V.T.K.; Wang, C.I.; Kwang, J.; He, F. Effective in vivo therapeutic IgG antibody against VP3 of enterovirus 71 with receptor-competing activity. Sci. Rep. 2017, 7, 46402. [Google Scholar] [CrossRef] [PubMed]
- Ku, Z.; Shi, J.; Liu, Q.; Huang, Z. Development of murine monoclonal antibodies with potent neutralization effects on enterovirus 71. J. Virol. Methods 2012, 186, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Ku, Z.; Ye, X.; Huang, X.; Cai, Y.; Liu, Q.; Li, Y.; Su, Z.; Huang, Z. Neutralizing antibodies induced by recombinant virus-like particles of enterovirus 71 genotype C4 inhibit infection at pre- and post-attachment steps. PLoS ONE 2013, 8, e57601. [Google Scholar] [CrossRef]
- Zhang, C.; Ku, Z.; Liu, Q.; Wang, X.; Chen, T.; Ye, X.; Li, D.; Jin, X.; Huang, Z. High-yield production of recombinant virus-like particles of enterovirus 71 in Pichia pastoris and their protective efficacy against oral viral challenge in mice. Vaccine 2015, 33, 2335–2341. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rattanapisit, K.; Chao, Z.; Siriwattananon, K.; Huang, Z.; Phoolcharoen, W. Plant-Produced Anti-Enterovirus 71 (EV71) Monoclonal Antibody Efficiently Protects Mice Against EV71 Infection. Plants 2019, 8, 560. https://doi.org/10.3390/plants8120560
Rattanapisit K, Chao Z, Siriwattananon K, Huang Z, Phoolcharoen W. Plant-Produced Anti-Enterovirus 71 (EV71) Monoclonal Antibody Efficiently Protects Mice Against EV71 Infection. Plants. 2019; 8(12):560. https://doi.org/10.3390/plants8120560
Chicago/Turabian StyleRattanapisit, Kaewta, Zhang Chao, Konlavat Siriwattananon, Zhong Huang, and Waranyoo Phoolcharoen. 2019. "Plant-Produced Anti-Enterovirus 71 (EV71) Monoclonal Antibody Efficiently Protects Mice Against EV71 Infection" Plants 8, no. 12: 560. https://doi.org/10.3390/plants8120560
APA StyleRattanapisit, K., Chao, Z., Siriwattananon, K., Huang, Z., & Phoolcharoen, W. (2019). Plant-Produced Anti-Enterovirus 71 (EV71) Monoclonal Antibody Efficiently Protects Mice Against EV71 Infection. Plants, 8(12), 560. https://doi.org/10.3390/plants8120560