Role of Salicylic Acid and Components of the Phenylpropanoid Pathway in Basal and Cultivar-Related Resistance of Oilseed Rape (Brassica napus) to Verticillium longisporum


Abstract
1. Introduction
2. Results
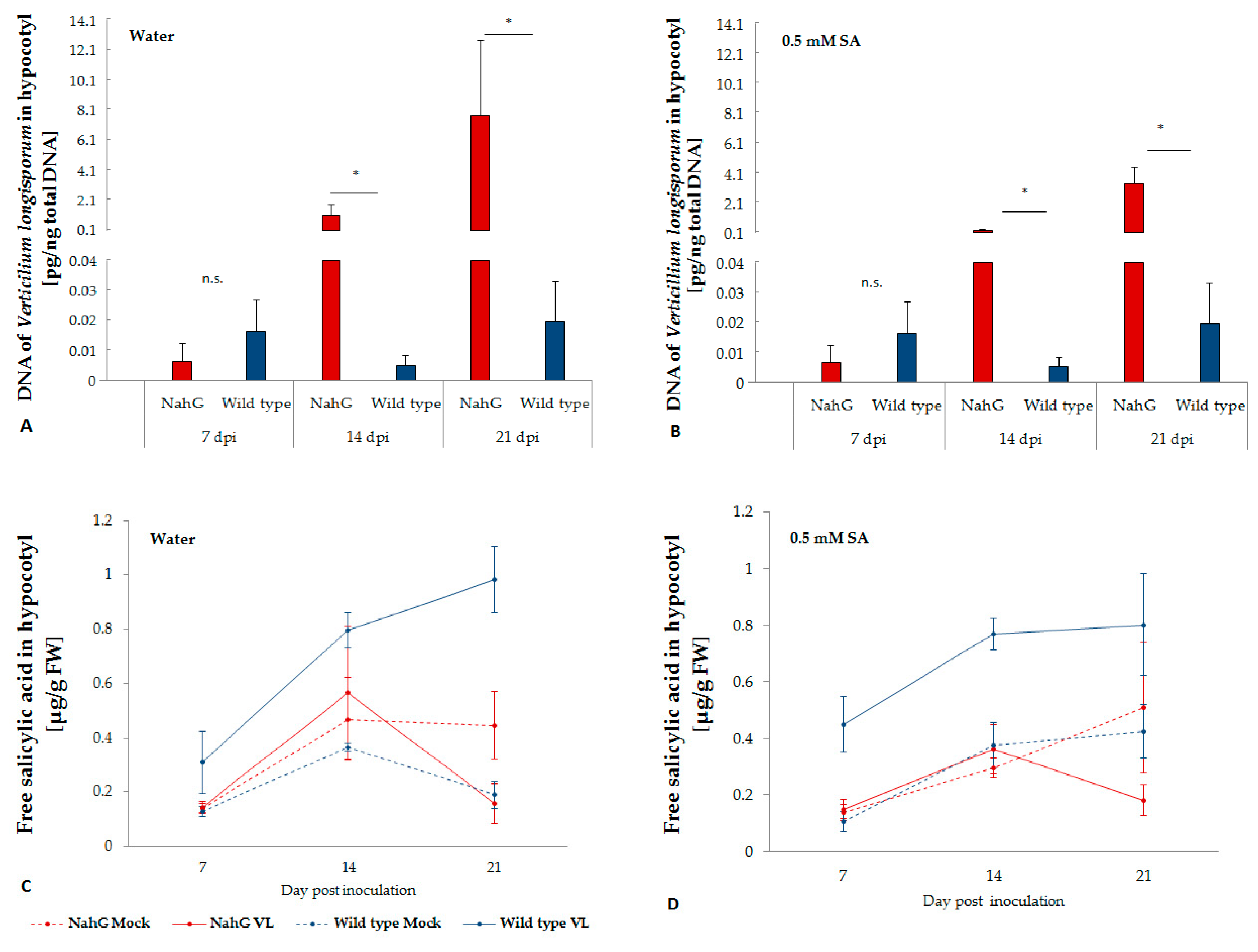
2.1. V. longisporum Disease Development and Plant Colonization in NahG Transformed Oilseed Rape
2.2. Endogenous SA in the Hypocotyl of Wild Type and NahG Transformant Plants
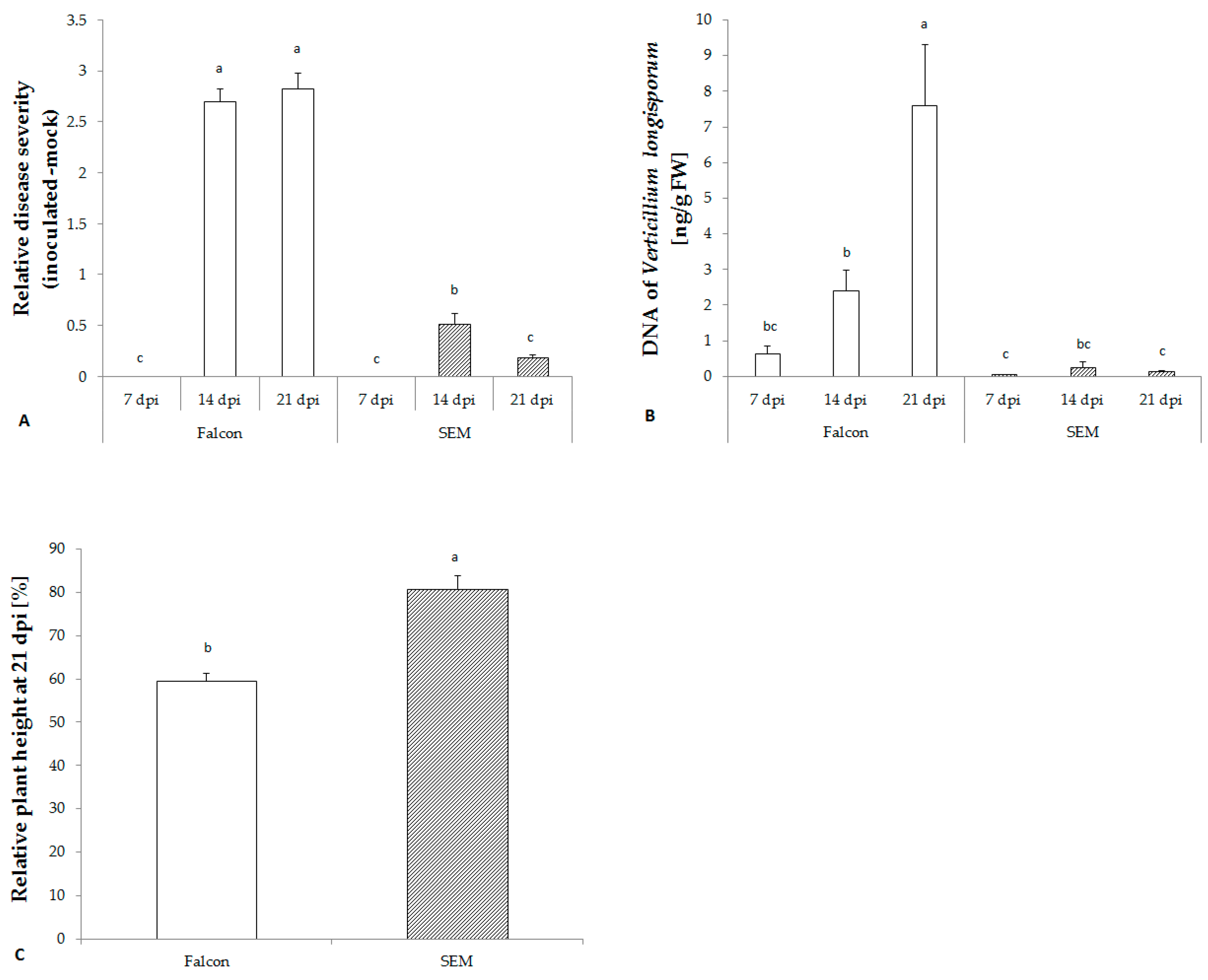
2.3. V. longisporum Disease Development in Resistant and Susceptible Cultivars
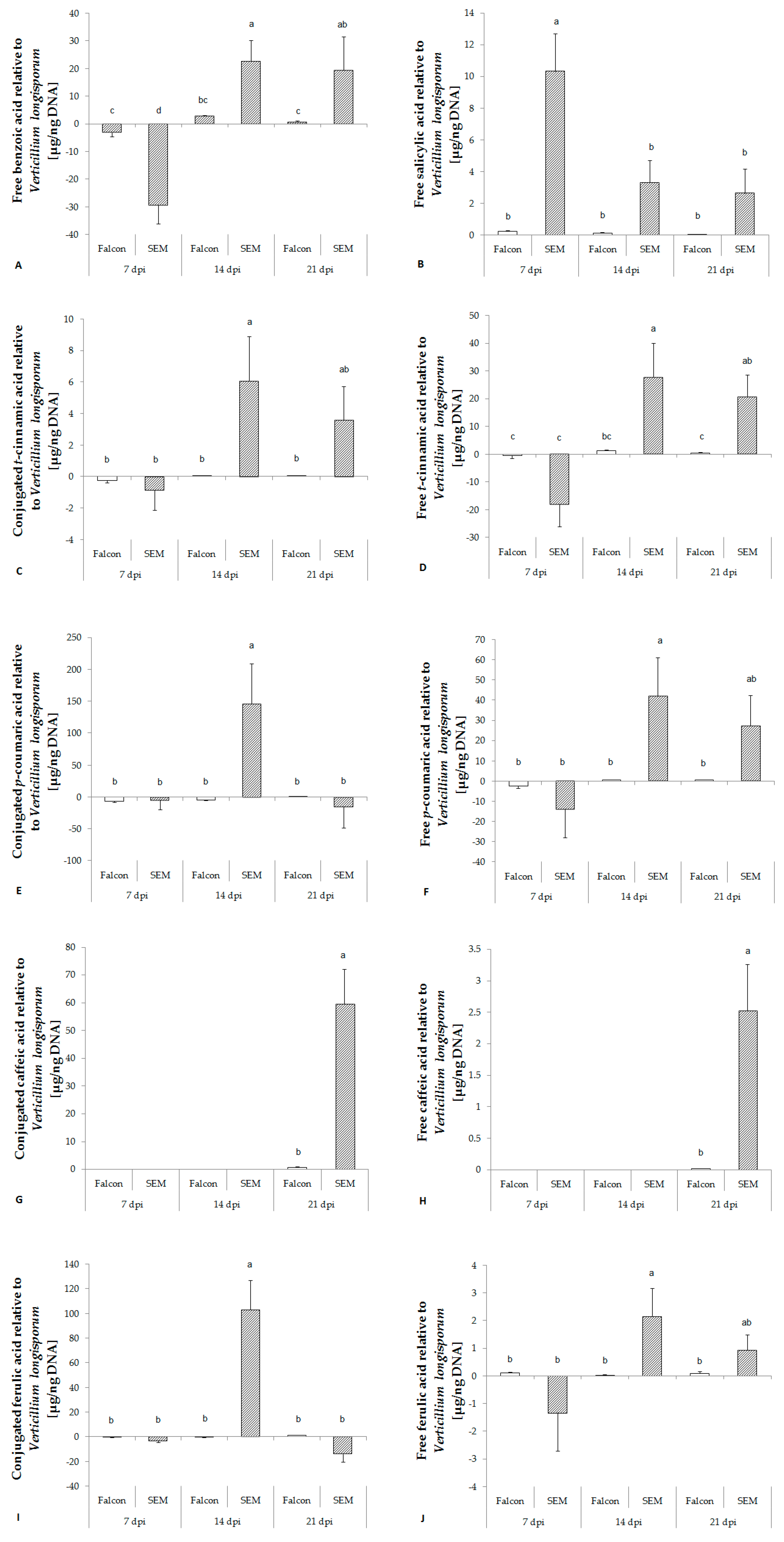
2.4. Changes in the SA Biosynthetic Pathway of B. napus Infected with V. longisporum
2.5. Changes in Phenolic Acid Levels related to Lignin Biosynthesis in Response to V. longisporum Infection
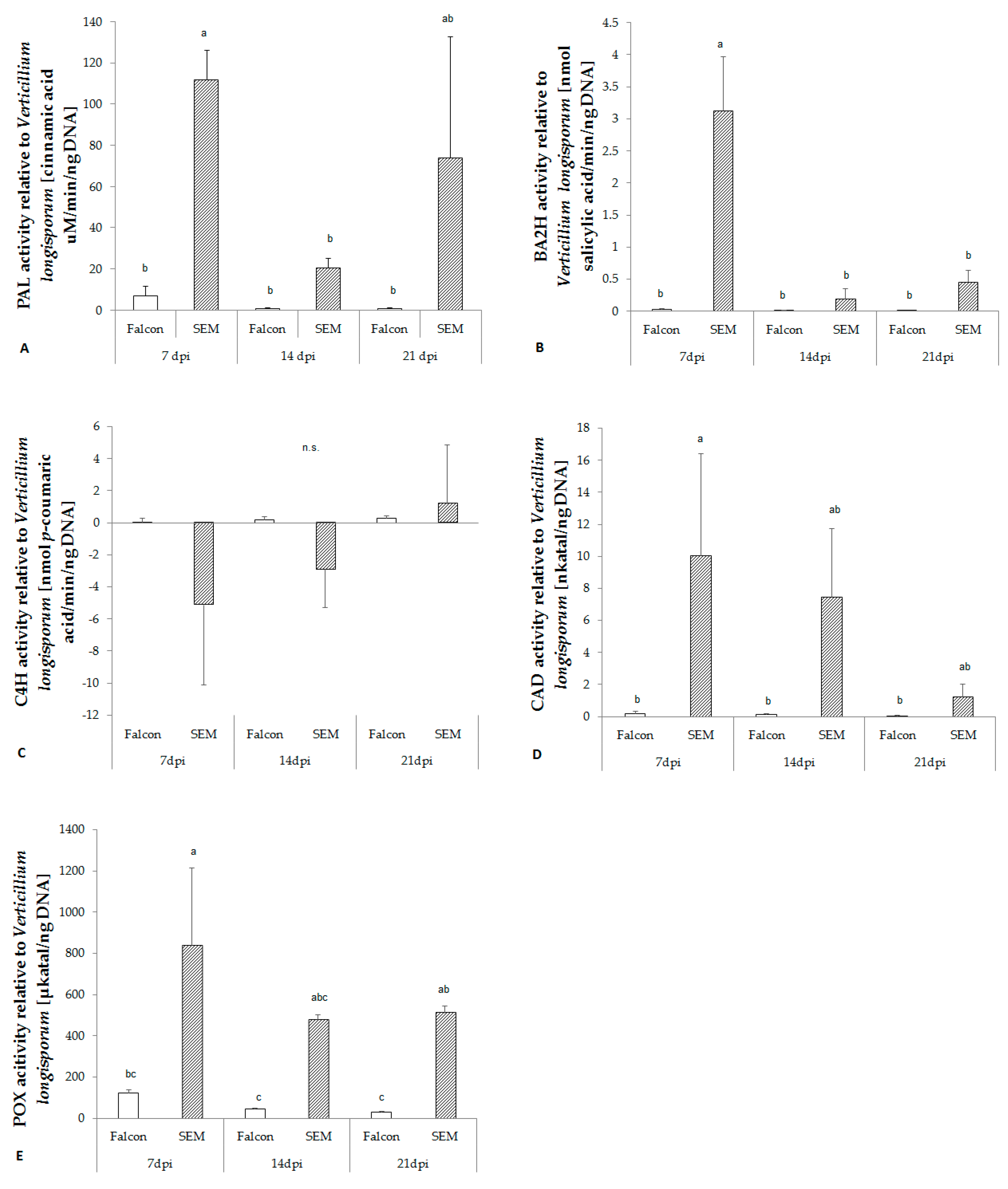
2.6. Activity of Key Enzymes in Lignin Biosynthetic Pathway
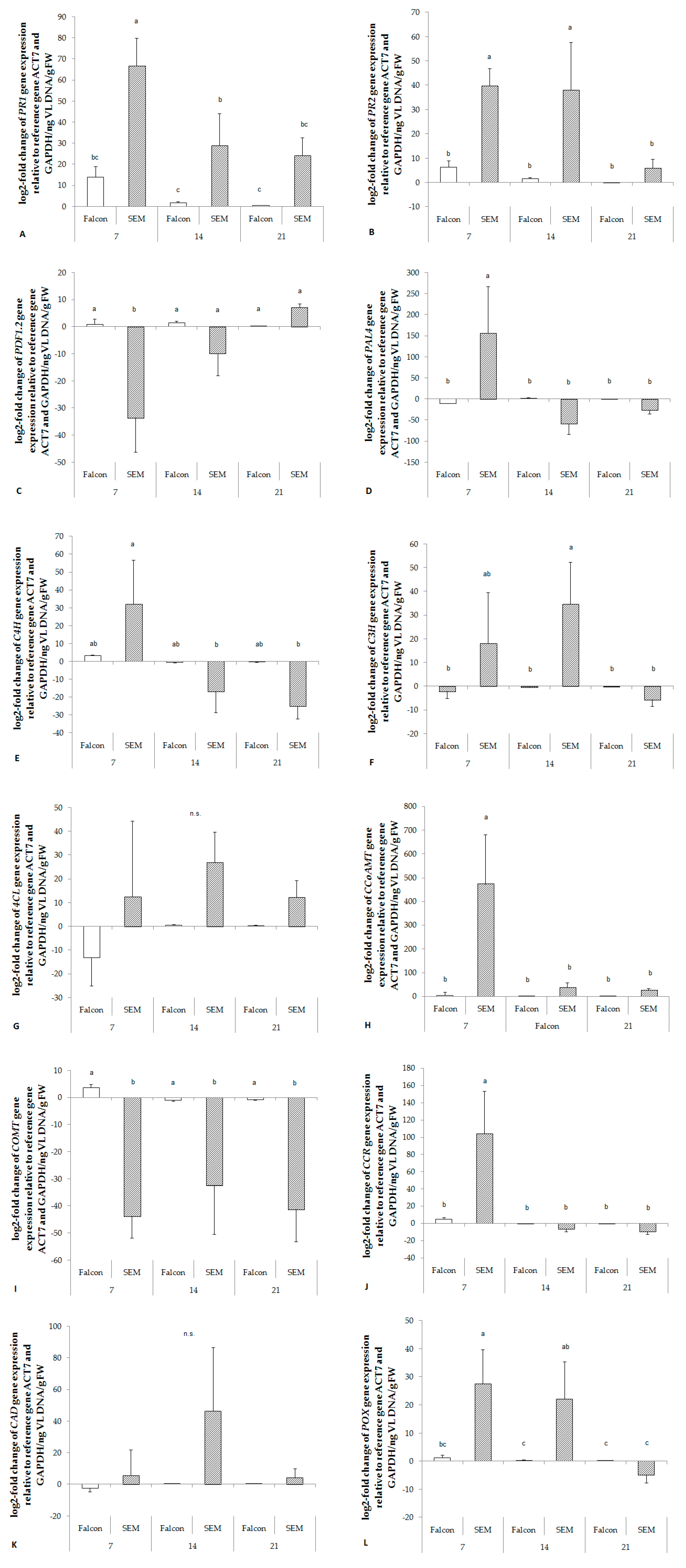
2.7. Regulation of Genes of Key Enzymes involved in Lignin Synthesis
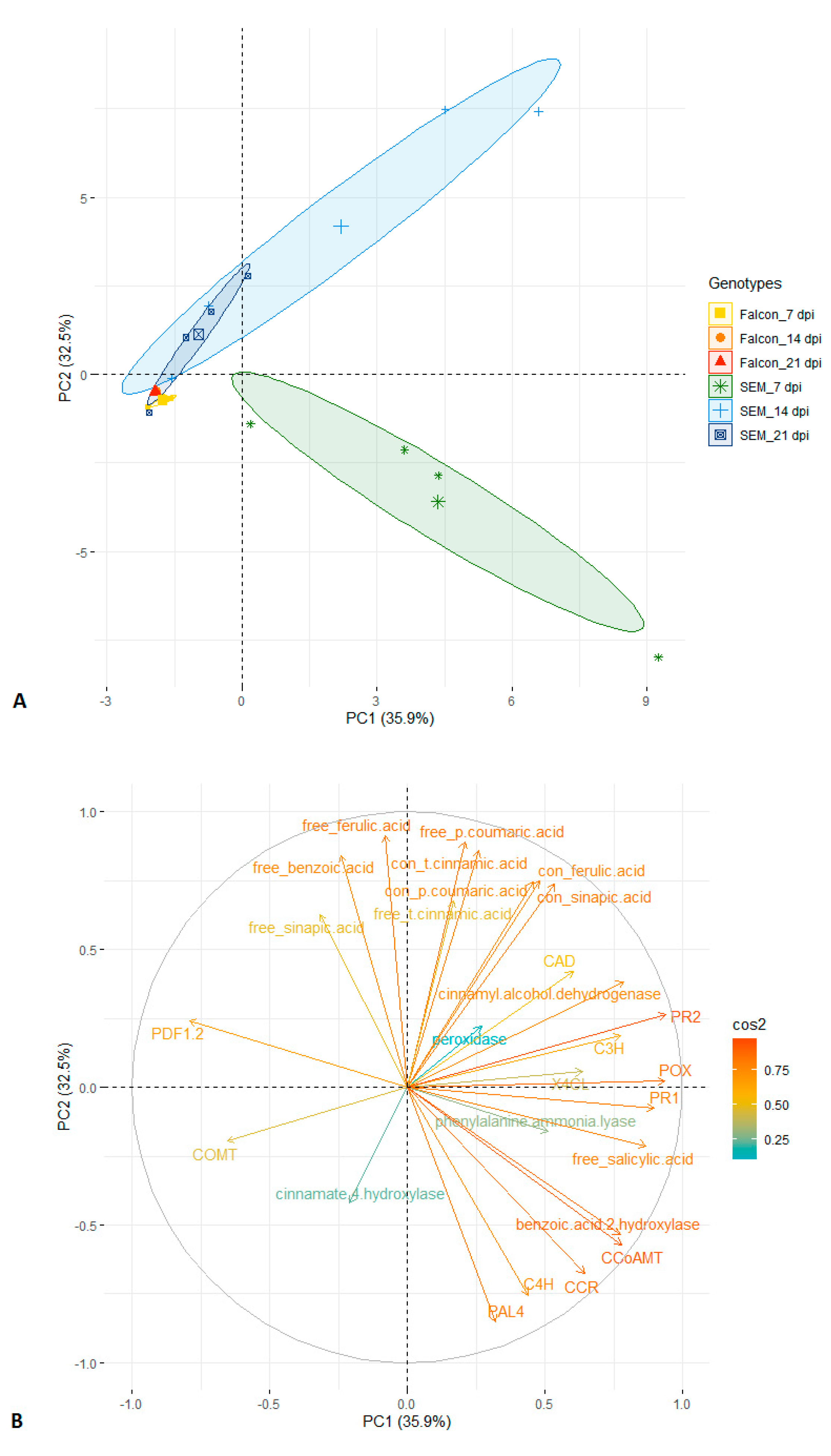
2.8. Principal Component Analysis
3. Discussion
3.1. SA Plays a Role in Basal Resistance of B. napus to V. longisporum
3.2. Role of SA and Phenolic Acids in Cultivar Resistance of B. napus to V. longisporum
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Cultivation
5.2. Treatments and Experimental Design
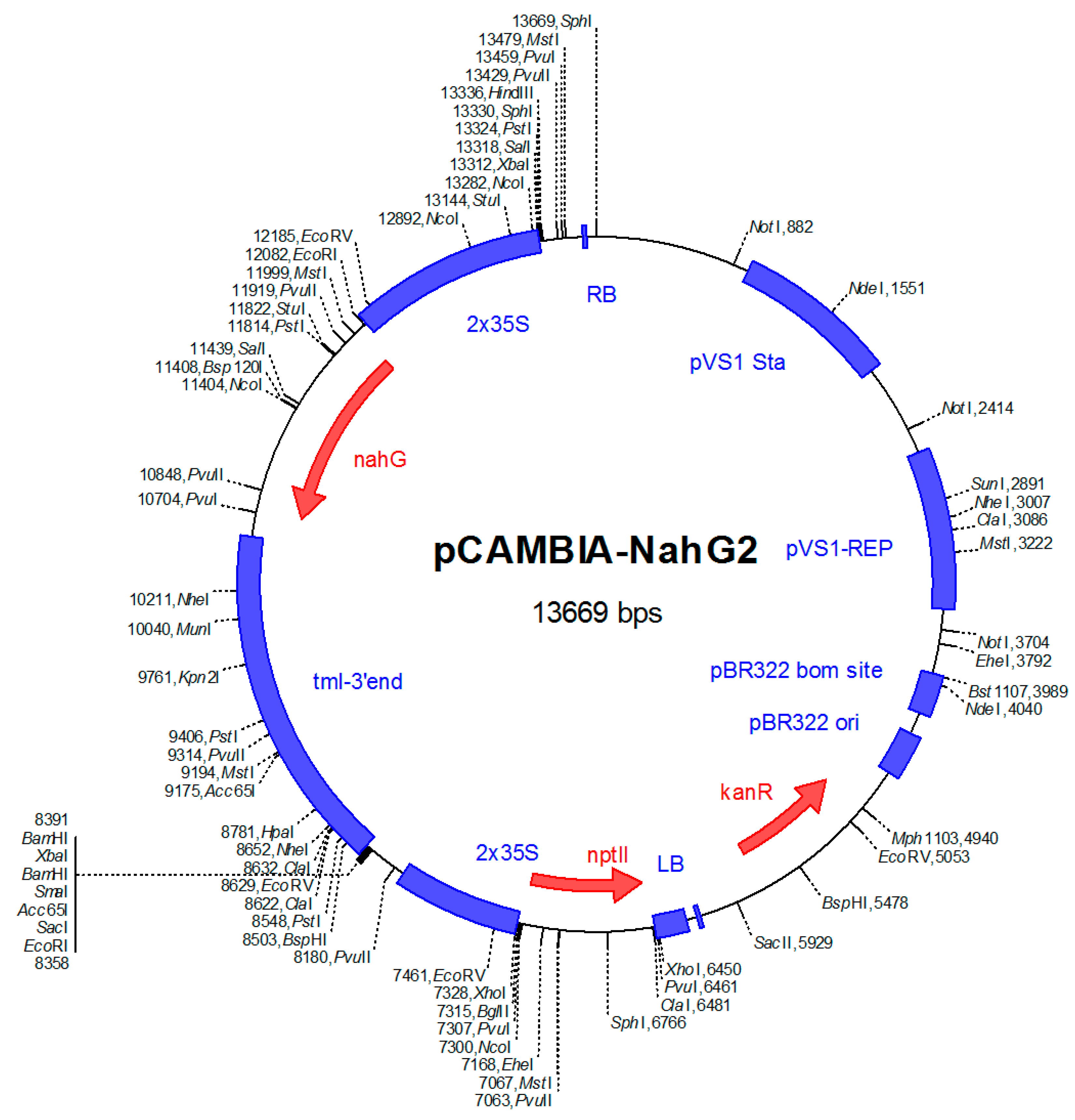
5.3. Production of Transgenic B. napus Expressing the NahG Gene
5.4. Exogenous Application of SA
5.5. Fungal Culture and Inoculation
5.6. Disease Assessment
5.7. Extraction and Quantification of Fungal DNA
5.8. Quantification of Endogenous SA
5.9. Quantification of Phenolic Acids in Hypocotyls
5.10. Enzyme Assays
5.11. Gene Expression
5.11.1. RNA Extraction and Synthesis of cDNA
5.11.2. Reverse Transcription Quantitative PCR (RT-qPCR)
5.12. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Inderbitzin, P.; Subbarao, K.V. Verticillium systematics and evolution: How confusion impedes Verticillium wilt management and how to resolve it. Phytopathology 2014, 104, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Steventon, L.A.; Fahleson, J.; Hu, Q.; Dixelius, C. Identification of the causal agent of Verticillium wilt of winter oilseed rape in Sweden, V. longisporum. Mycol. Res. 2002, 106, 570–578. [Google Scholar] [CrossRef]
- Gladders, P.; Smith, J.A.; Kirkpatrick, L.; Clewes, E.; Grant, C.; Barbara, D.; Barnes, A.V.; Lane, C.R. First record of Verticillium wilt (Verticillium longisporum) in winter oilseed rape in the UK. New Dis. Rep. 2011, 23, 8. [Google Scholar] [CrossRef]
- Zeise, K.; von Tiedemann, A. Host specialization among vegetative compatibility groups of Verticillium dahliae in relation to Verticillium longisporum. J. Phytopathol. 2002, 150, 112–119. [Google Scholar] [CrossRef]
- Canadian Food Inspection Agency. Pest Risk Management Document: Verticillium longisporum (Verticillium stripe). Available online: http://www.inspection.gc.ca/plants/plant-pests-invasive-species/directives/risk-management/rmd-17-01/eng/1487004855251/1487004951480 (accessed on 13 November 2018).
- Depotter, J.R.L.; Deketelaere, S.; Inderbitzin, P.; von Tiedemann, A.; Höfte, M.; Subbarao, K.V.; Wood, T.A.; Thomma, B.P.H.J. Verticillium longisporum, the invisible threat to oilseed rape and other brassicaceous plant hosts. Mol. Plant. Pathol. 2016, 17, 1004–1016. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S. Longevity of the Verticillium wilt fungus in the laboratory and field. Phytopathology 1955, 455, 180–181. [Google Scholar]
- Karapapa, V.K.; Bainbridge, B.W.; Heale, J.B. Morphological and molecular characterization of Verticillium longisporum comb, nov., pathogenic to oilseed rape. Mycol. Res. 1997, 101, 1281–1294. [Google Scholar] [CrossRef]
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity, pathogenicity, and management of verticillium species. Annu. Rev. Phytopathol. 2009, 47, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, Y.; Bouazizi, E.; Triki, M. Inductions of defense response in olive plants against Verticillium dahliae through application of salicylic acid as abiotic inducer. J. Adv. Biol. Biotechnol. 2016, 5, 1–9. [Google Scholar] [CrossRef]
- Maruri-López, I.; Aviles-Baltazar, N.Y.; Buchala, A.; Serrano, M. Intra and extracellular journey of the phytohormone salicylic acid. Front. Plant. Sci. 2019, 10, 423. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Spence, M.P.; Bobokalonova, A.; Carballo, V.; Glinkerman, C.M.; Pluskal, T.; Shen, A.; Weng, J.-K. PBS3 and EPS1 complete salicylic acid biosynthesis from isochorismate in Arabidopsis. bioRxiv 2019, 601948. [Google Scholar] [CrossRef]
- Achuo, E.A.; Audenaert, K.; Meziane, H.; Hofte, M. The salicylic acid-dependent defence pathway is effective against different pathogens in tomato and tobacco. Plant. Pathol. 2004, 53, 65–72. [Google Scholar] [CrossRef]
- Loake, G.; Grant, M. Salicylic acid in plant defence--the players and protagonists. Curr. Opin. Plant. Biol. 2007, 10, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Ratzinger, A.; Riediger, N.; von Tiedemann, A.; Karlovsky, P. Salicylic acid and salicylic acid glucoside in xylem sap of Brassica napus infected with Verticillium longisporum. J. Plant. Res. 2009, 122, 571–579. [Google Scholar] [CrossRef] [PubMed]
- König, S.; Feussner, K.; Kaever, A.; Landesfeind, M.; Thurow, C.; Karlovsky, P.; Gatz, C.; Polle, A.; Feussner, I. Soluble phenylpropanoids are involved in the defense response of Arabidopsis against Verticillium longisporum. New Phytol. 2014, 202, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Obermeier, C.; Hossain, M.A.; Snowdon, R.; Knüfer, J.; von Tiedemann, A.; Friedt, W. Genetic analysis of phenylpropanoid metabolites associated with resistance against Verticillium longisporum in Brassica napus. Mol. Breed. 2013, 31, 347–361. [Google Scholar] [CrossRef]
- Eynck, C.; Koopmann, B.; Karlovsky, P.; von Tiedemann, A. Internal resistance in winter oilseed rape inhibits systemic spread of the vascular pathogen Verticillium longisporum. Phytopathology 2009, 99, 802–811. [Google Scholar] [CrossRef] [PubMed]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Cardoza, Y.J.; Schmelz, E.A.; Raina, R.; Engelberth, J.; Tumlinson, J.H. Differential volatile emissions and salicylic acid levels from tobacco plants in response to different strains of Pseudomonas syringae. Planta 2003, 217, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Song, J.T.; Koo, Y.J.; Seo, H.S.; Kim, M.C.; Choi, Y.D.; Kim, J.H. Overexpression of AtSGT1, an Arabidopsis salicylic acid glucosyltransferase, leads to increased susceptibility to Pseudomonas syringae. Phytochemistry 2008, 69, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Koo, Y.J.; Kim, M.A.; Kim, E.H.; Song, J.T.; Jung, C.; Moon, J.-K.; Kim, J.-H.; Seo, H.S.; Song, S.I.; Kim, J.-K.; et al. Overexpression of salicylic acid carboxyl methyltransferase reduces salicylic acid-mediated pathogen resistance in Arabidopsis thaliana. Plant. Mol. Biol. 2007, 64, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Eynck, C.; Koopmann, B.; Grunewaldt-Stoecker, G.; Karlovsky, P.; von Tiedemann, A. Differential interactions of Verticillium longisporum and V. dahliae with Brassica napus detected with molecular and histological techniques. Eur. J. Plant. Pathol. 2007, 118, 259–274. [Google Scholar] [CrossRef]
- Johansson, A.; Staal, J.; Dixelius, C. Early responses in the Arabidopsis-Verticillium longisporum pathosystem are dependent on NDR1, JA-and ET-associated signals via cytosolic NPR1 and RFO1. Mol. Plant. Microbe. Interact. 2006, 19, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X. Verticillium longisporum on Oilseed Rape (Brassica napus L.): Differential Roles of Salicylic Acid, Seed Transmission and Plant Colonization in Greenhouse and Field Conditions. Ph.D. Thesis, Georg-August University Göttingen, Göttingen, Germany, December 2018. [Google Scholar]
- Kuai, X.; MacLeod, B.J.; Després, C. Integrating data on the Arabidopsis NPR1/NPR3/NPR4 salicylic acid receptors; a differentiating argument. Front. Plant. Sci. 2015, 6, 235. [Google Scholar] [CrossRef] [PubMed]
- Camm, E.L.; Towers, G.H.N. Phenylalanine ammonia lyase. Phytochemistry 1973, 12, 961–973. [Google Scholar] [CrossRef]
- Miedes, E.; Vanholme, R.; Boerjan, W.; Molina, A. The role of the secondary cell wall in plant resistance to pathogens. Front. Plant. Sci. 2014, 5, 358. [Google Scholar] [CrossRef] [PubMed]
- Maury, S.; Delaunay, A.; Mesnard, F.; Crônier, D.; Chabbert, B.; Geoffroy, P.; Legrand, M. O-methyltransferase(s)-suppressed plants produce lower amounts of phenolic vir inducers and are less susceptible to Agrobacterium tumefaciens infection. Planta 2010, 232, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Reusche, M.; Thole, K.; Janz, D.; Truskina, J.; Rindfleisch, S.; Drübert, C.; Polle, A.; Lipka, V.; Teichmann, T. Verticillium infection triggers VASCULAR-RELATED NAC DOMAIN7-dependent de novo xylem formation and enhances drought tolerance in Arabidopsis. Plant. Cell 2012, 24, 3823–3837. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Song, T.; Zhang, X.; Yuan, H.; Su, L.; Li, W.; Xu, J.; Liu, S.; Chen, L.; Chen, T.; et al. Unconventionally secreted effectors of two filamentous pathogens target plant salicylate biosynthesis. Nat. Commun. 2014, 5, 4686. [Google Scholar] [CrossRef] [PubMed]
- Rahman, T.A.E.; Oirdi, M.E.; Gonzalez-Lamothe, R.; Bouarab, K. Necrotrophic pathogens use the salicylic acid signaling pathway to promote disease development in tomato. Mol. Plant. Microbe. Interact. 2012, 25, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Weier, D.; Hanke, C.; Eickelkamp, A.; Lühs, W.; Dettendorfer, J.; Schaffert, E.; Möllers, C.; Friedt, W.; Wolter, F.P.; Frentzen, M. Trierucoylglycerol biosynthesis in transgenic plants of rapeseed (Brassica napus L.). Lipid 1997, 99, 160–165. [Google Scholar] [CrossRef]
- Hüsken, A.; Baumert, A.; Strack, D.; Becker, H.C.; Möllers, C.; Milkowski, C. Reduction of sinapate ester content in transgenic oilseed rape (Brassica napus) by dsRNAi-based Suppression of BnSGT1 Gene Expression. Mol. Breed. 2005, 16, 127–138. [Google Scholar] [CrossRef]
- Chung, Y.S.; Lee, N.R.; Cheon, C.L.; Song, E.S.; Lee, M.S.; Kim, Y.; Min, K.H. Molecular cloning of the nahG gene encoding salicylate hydroxylase from Pseudomonas fluorescens. Mol. Cells 2001, 11, 105–109. [Google Scholar] [PubMed]
- Zeise, K.; von Tiedemann, A. Morphological and physiological differentiation among vegetative compatibility groups of Verticillium dahliae in relation to V. longisporum. J. Phytopathol. 2001, 149, 469–475. [Google Scholar] [CrossRef]
- Eynck, C.; Koopmann, B.; von Tiedemann, A. Identification of Brassica accessions with enhanced resistance to Verticillium longisporum under controlled and field conditions. J. Plant. Dis. Protect. 2009, 116, 63–72. [Google Scholar] [CrossRef]
- Zheng, X.; Pfordt, A.; Khatri, L.; Eseola, A.B.; Wilch, A.; Koopmann, B.; Tiedemann, A. von. Contrasting patterns of colonization with Verticillium longisporum in winter and spring type oilseed rape (Brassica napus L.) in the field and greenhouse and the role of soil temperature. Plant. Dis. 2019, 103, 2090–2099. [Google Scholar] [CrossRef] [PubMed]
- Lopisso, D.T.; Knüfer, J.; Koopmann, B.; von Tiedemann, A. The vascular pathogen Verticillium longisporum does not affect water relations and plant responses to drought stress of its host, Brassica napus. Phytopathology 2017, 107, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Kamble, A.; Bhargava, S. ß-aminobutyric acid-induced resistance in Brassica juncea against the necrotrophic pathogen Alternaria brassicae. J. Phytopathol. 2007, 155, 152–158. [Google Scholar] [CrossRef]
- Ayaz, F.A.; Hayırlıoglu-Ayaz, S.; Alpay-Karaoglu, S.; Grúz, J.; Valentová, K.; Ulrichová, J.; Strnad, M. Phenolic acid contents of kale (Brassica oleraceae L. var. acephala DC.) extracts and their antioxidant and antibacterial activities. Food Chem. 2008, 107, 19–25. [Google Scholar] [CrossRef]
- Mattila, P.; Kumpulainen, J. Determination of free and total phenolic acids in plant-derived foods by HPLC with diode-array detection. J. Agric. Food Chem. 2002, 50, 3660–3667. [Google Scholar] [CrossRef] [PubMed]
- Leon, J.; Yalpani, N.; Raskin, I.; Lawton, M.A. Induction of benzoic acid 2-hydroxylase in virus-inoculated tobacco. Plant. Physiol. 1993, 103, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Kamble, A.; Koopmann, B.; von Tiedemann, A. Induced resistance to Verticillium longisporum in Brassica napus by β-aminobutyric acid. Plant. Pathol. 2013, 62, 552–561. [Google Scholar] [CrossRef]
- Salvador, V.H.; Lima, R.B.; dos Santos, W.D.; Soares, A.R.; Böhm, P.A.F.; Marchiosi, R.; Ferrarese, M.d.L.L.; Ferrarese-Filho, O. Cinnamic acid increases lignin production and inhibits soybean root growth. PLoS ONE 2013, 8, e69105. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Alejo, N.; Gómez-Peralta, J.E. Activity of enzymes involved in capsaicin biosynthesis in callus tissue and fruits of chili pepper (Capsicum annuum L.). J. Plant. Physiol. 1993, 141, 147–152. [Google Scholar] [CrossRef]
- Phimchan, P.; Chanthai, S.; Bosland, P.W.; Techawongstien, S. Enzymatic changes in phenylalanine ammonia-lyase, cinnamic-4-hydroxylase, capsaicin synthase, and peroxidase activities in capsicum under drought stress. J. Agric. Food Chem. 2014, 62, 7057–7062. [Google Scholar] [CrossRef] [PubMed]
- Chabannes, M.; Barakate, A.; Lapierre, C.; Marita, J.M.; Ralph, J.; Pean, M.; Danoun, S.; Halpin, C.; Grima-Pettenati, J.; Boudet, A.M. Strong decrease in lignin content without significant alteration of plant development is induced by simultaneous down-regulation of cinnamoyl CoA reductase (CCR) and cinnamyl alcohol dehydrogenase (CAD) in tobacco plants. Plant. J. 2001, 28, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Mitra, A. Reinforcement of cell wall in roots of Lycopersicon esculentum through induction of phenolic compounds and lignin by elicitors. Physiol. Mol. Plant. Pathol. 2007, 71, 201–209. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 45. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Treatment | 0.5 mM SA Root Treatment | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NahG | Wild Type | NahG | Wild Type | |||||||
| Mock | VL | Mock | VL | Mock | VL | Mock | VL | |||
| Disease severity [1,2,3,4,5,6,7,8,9] | 7 dpi | 1.1 ± 0.07 a | 1.3 ± 0.12 a | 1.0 ± 0.00 a | 1.1 ± 0.05 a | 1.2 ± 0.09 a | 1.3 ± 0.10 a | 1.0 ± 0.00 a | 1.0 ± 0.00 a | |
| 14 dpi | 1.2 ± 0.09 b | 4.3 ± 0.20 a | 1.1 ± 0.05 b | 1.2 ± 0.08 b | 1.2 ± 0.09 b | 3.2 ± 0.34 a | 1.0 ± 0.00 b | 1.3 ± 0.00 b | ||
| 21 dpi | 1.4 ± 0.15 b | 5.5 ± 0.15 a | 1.2 ± 0.08 b | 1.6 ± 0.18 b | 1.4 ± 0.15 b | 4.9 ± 0.18 a | 1.2 ± 0.09 b | 1.6 ± 0.20 b | ||
| Plant height [cm] | 22.5 ± 1.3 c | 6.7 ± 0.4 d | 34.6 ± 2.8 a | 23.2 ± 2.0 c | 24.5 ± 1.5 bc | 7.4 ± 0.4 d | 31.5 ± 2.7 ab | 21.0 ± 1.4 c | ||
| Dry weight [mg] | 7 dpi | root | 28 ± 1.1 a | 30 ± 4.7 a | 24 ± 3.2 ab | 31 ± 1.6 a | 23 ± 1.9 ab | 17 ± 0.9 b | 28 ± 1.5 a | 26 ± 3.8 ab |
| shoot | 134 ± 5.1 ab | 136 ± 18.7 ab | 136 ± 9.3 ab | 153 ± 6.9 a | 115 ± 7.7 b | 122 ± 15.9 ab | 134 ± 6.8 ab | 150 ± 6.3 ab | ||
| 14 dpi | root | 74 ± 9.5 a | 64 ± 9.8 a | 70 ± 16.5 a | 63 ± 9.4 a | 61 ± 6.6 a | 57 ± 7.0 a | 87 ± 18.6 a | 63 ± 4.3 a | |
| shoot | 375 ± 64.2 a | 309 ± 29.0 a | 400 ± 51.8 a | 342 ± 9.4 a | 296±23.7 a | 277 ± 25.6 a | 409 ± 18.6 a | 339 ± 4.3 a | ||
| 21 dpi | root | 132 ± 11.1 b | 70 ± 4.8 c | 170 ± 8.3 a | 128 ± 10.9 b | 116±10.4 b | 55 ± 5.2 c | 140 ± 14.9 ab | 131 ± 10.3 b | |
| shoot | 662 ± 89.7 a | 383 ± 80.6 b | 1144 ± 192.2 a | 847 ± 192.2 a | 743±142.4 a | 380 ± 88.1 b | 1000 ± 156.4 a | 684 ± 60.8 a | ||
| Score | Symptom Development |
|---|---|
| 1 | No symptoms |
| 2 | Weak symptoms on the oldest leaf (yellowing, black veins) |
| 3 | Weak symptoms on the next younger leaves |
| 4 | About 50% of the leaves have symptoms |
| 5 | More than 50% of the leaves have symptoms |
| 6 | Up to 50% of the leaves are dead |
| 7 | More than 50% of the leaves are dead |
| 8 | Only apex is still alive |
| 9 | The plant is dead |
| Step | qPCR for Quantification of V. longisporum | RT-qPCR for Gene Expression |
|---|---|---|
| Initial Denaturation | 95 °C, 4 min | 95 °C, 4 min |
| Denaturation | 95 °C, 10 s | 95 °C, 10 s |
| Annealing | 60 °C, 15 s | 64.8 °C, 15 s |
| Extension | 72 °C, 15 s | 72 °C, 15 s |
| Repeat Times | 40 cycles | 40 cycles |
| Melting Curve Analysis | 55 to 95 °C | 60 to 95 °C |
| Gene | NCBI Accession | Primer Sequence | PCR Efficiency [%] | Reference | |
|---|---|---|---|---|---|
| 4CL | XM_013895971.1 | F | ACGCCGAGATGAAAATCATC | 106.2 | This study |
| R | CCGTCTTTGTCAATGGTCTC | ||||
| ACT7 | NM_001316079.1 | F | GCTGACCGTATGAGCAAAG | 73.6 | Wang et al. 2014 |
| R | AAGATGGATGGACCCGAC | ||||
| C3H | XM_013879044.1 | F | AGACCAGAGAGGTTCTTGGA | 119.4 | This study |
| R | CGAGTCCAGGGTTTTCAGAC | ||||
| C4H | XM_013888134.1 | F | GTATGTGCCGTTTGGTGTTG | 73.8 | This study |
| R | GGACCTTGGCTTCATTACGA | ||||
| CAD | XM_013817405.1 | F | GGTGGCTTCGCTGACACTAT | 70.2 | This study |
| R | TCACACCCATGTGTCCAACT | ||||
| CCoAMT | XM_013799238.1 | F | TTCAAGGCAGCACACGATAG | 127.3 | This study |
| R | TGCCATACTTGTGGACCGTA | ||||
| CCR | XM_013836581.1 | F | TCCGCTAAGACTTACGCTAATC | 74.8 | This study |
| R | CCTCGTAGACCAGCACATGG | ||||
| COMT | XM_013793239.1 | F | CCGGAAAAAGGGAAAGTGATC | 123 | This study |
| R | TCACATCGAATAAAACCTGACC | ||||
| GAPDH | XM_013856115.1 | F | CGCTTCCTTCAACATCATTCCCA | 95.6 | Alkooranee et al. 2015 |
| R | TCAGATTCCTCCTTGATAGCCTT | ||||
| PAL4 | XM_013817346.1 | F | GGCACGGACAGTTATGGAGT | 96.8 | This study |
| R | GCCGACTTAGGTAGCGTGAG | ||||
| PDF1.2 | XM_013862352.1 | F | ATCACCCTTCTCTTCGCTGCTCTC | 109.4 | Wu et al. 2016 |
| R | CATACTCCTGACCATGTCCCACTAG | ||||
| POX | XM_013786965.1 | F | CTCTCTGGGGGTCACACATT | 82.3 | This study |
| R | TGTCGAAAACCGTAGGGGTA | ||||
| PR1 | XM_013877950.1 | F | AAAGCTACGCCGACCGACTACGAG | 108.8 | Alkooranee et al. 2015 |
| R | CCAGAAAAGTCGGCGCTACTCCA | ||||
| PR2 | AF229403.1 | F | GTACGCTCTGTTCAAACCGACCC | 109.1 | Alkooranee et al. 2015 |
| R | TTTCCAACGATCCTCCGCCTGA | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Koopmann, B.; von Tiedemann, A. Role of Salicylic Acid and Components of the Phenylpropanoid Pathway in Basal and Cultivar-Related Resistance of Oilseed Rape (Brassica napus) to Verticillium longisporum. Plants 2019, 8, 491. https://doi.org/10.3390/plants8110491
Zheng X, Koopmann B, von Tiedemann A. Role of Salicylic Acid and Components of the Phenylpropanoid Pathway in Basal and Cultivar-Related Resistance of Oilseed Rape (Brassica napus) to Verticillium longisporum. Plants. 2019; 8(11):491. https://doi.org/10.3390/plants8110491
Chicago/Turabian StyleZheng, Xiaorong, Birger Koopmann, and Andreas von Tiedemann. 2019. "Role of Salicylic Acid and Components of the Phenylpropanoid Pathway in Basal and Cultivar-Related Resistance of Oilseed Rape (Brassica napus) to Verticillium longisporum" Plants 8, no. 11: 491. https://doi.org/10.3390/plants8110491
APA StyleZheng, X., Koopmann, B., & von Tiedemann, A. (2019). Role of Salicylic Acid and Components of the Phenylpropanoid Pathway in Basal and Cultivar-Related Resistance of Oilseed Rape (Brassica napus) to Verticillium longisporum. Plants, 8(11), 491. https://doi.org/10.3390/plants8110491