Abstract
In plants, the Gretchen GH3 (GH3) protein is involved in free auxin (IAA) and amino acid conjugation, thus controlling auxin homeostasis. To date, many GH3 gene families have been identified from different plant species. However, the GH3 gene family in kiwifruit (Actinidia chinensis) has not been reported. In this study, 12 AcGH3 genes were identified, phylogenetic analysis of AtGH3 (Arabidopsis), SlGH3 (Solanum lycopersicum), and AcGH3 provided insights into various orthologous relationships among these proteins, which were categorized into three groups. Expression analysis of AcGH3 genes at different postharvest stages suggested limited or no role for most of the AcGH3 genes at the initiation of fruit ripening. AcGH3.1 was the only gene exhibiting ripening-associated expression. Further study showed that the expression of AcGH3.1 gene was induced by NAA (1-naphthylacetic acid, auxin analogue) and inhibited by 1-MCP (1-methylcyclopropene, ethylene receptor inhibitor), respectively. AcGH3.1 gene silencing inhibited gene expression and delayed fruit softening in kiwifruit. The results indicate that AcGH3.1 may play an important role in the softening process of fruits. Analysis of the AcGH3.1 promoter revealed the presence of many cis-elements related to hormones, light, and drought. The determination of GUS (β-Galactosidase) enzyme activity revealed that promoter activity increased strikingly upon abscisic acid (ABA), ethylene, or NAA treatment, and significantly decreased with salicylic acid (SA) treatment. The present study could help in the identification of GH3 genes and revelation of AcGH3.1 gene function during postharvest stages, which pave the way for further functional verification of the AcGH3.1 gene.
1. Introduction
Kiwifruits have long been called the ‘king of fruits’ due to its unique flavor and exceptional nutritional value. However, the fruit quality of kiwifruits not only declines rapidly after ripening, but also is not resistant to storage and transportation. The speed of softening after harvesting directly determines the shelf life, flavor and edible value of the fruit. Many studies have found that auxin (IAA) plays an important role in fruit ripening and aging. Exogenous IAA treatment of immature tomato fruit can delay fruit ripening and lead to parthenocarpy [1]. It is believed that IAA can delay the ripening of the climacteric fruit, and the fruit starts to fully mature due to the loss of hormone function of IAA [2].
Reduced IAA levels in fruits entering the mature stage can be achieved and maintained through changes in certain aspects of auxin metabolism [3,4]. Among them, IAA conjugation with amino acids is of particular interest because it is catalyzed by auxin induced proteins to provide a negative feedback loop to control auxin homeostasis [5]. Currently, various IAA-amino acid conjugates have been identified as endogenous compounds in a range of different plant species. In fruits such as tomatoes, grapes, pears, etc., the content of free IAA began to decrease before fruit ripening, which was accompanied by the increase of conjugated IAA (IAA-Asp) level, and it was believed that the ratio of free IAA/conjugated IAA determined the ripening of fruits, rather than the absolute amount of free IAA [1,6,7]. The conjugating reaction of free IAA to conjugated IAA is catalyzed by IAA amino acid synthase, which is encoded by the Gretchen Hagen 3 (GH3) gene. These proteins negatively regulate auxin homeostasis in plants [8]. Overexpression of OsGH3.1 resulted in reduced free IAA content and dwarfing of transgenic rice plants [9]. In addition, the identification of many IAA-amino acid conjugates as endogenous compounds in other different plant species has confirmed this mechanism in plants [10,11]. The first GH3 gene was isolated from soybean by differential screening after auxin treatment, and its transcription level was induced within 10 min after auxin treatment [8]. Since then, several members of the gene family have been identified in many plant species ranging from moss to gymnosperm to angiosperms [12,13,14].
Recent studies have shown that GH3 protein is involved in the growth and development of fleshy fruit, mainly by regulating the level of free IAA in the fruit to regulate fruit development and maturity. The role of the mature-associated GH3-1 protein in regulating endogenous IAA concentrations was investigated in grape berries and it was observed that this protein may be involved in establishing and maintaining low IAA concentrations in mature berries [15]. Further studies revealed that another GH3 protein, GH3-2, is also involved in the fruit ripening of grape berries [6]. In addition, differential expressions of two GH3 genes have also been reported during the development of the non-climacteric fruit longan. Among them, DlGH3.1 gene expression might be associated with pericarp growth, while DlGH3.2 accumulation is likely to be related to both pericarp growth and fruit ripening [16]. A CcGH3 gene was isolated from pepper, which was highly expressed in pericarp and placenta, and over-expressed in tomato accelerated fruit ripening, indicating that the CcGH3 gene was involved in fruit ripening [17]. Fifteen GH3 gene family members were isolated in a tomato, in which SlGH3-1 and SlGH3-2 showed mature-related expression and induction of auxin and ethylene. Therefore, this gene can be used to manipulate the maturation process in tomato fruit [18].
Kiwifruits are a climacteric fruit that softens quickly in the short term after harvest, which has huge limitations on its market value. IAA inhibits the ripening of kiwifruit, but the mechanism is not clear. Our study provides comprehensive information on the GH3 gene from kiwifruit, and analyzed the expression patterns during the postharvest process. We also analyzed the promoter activity of the AcGH3.1 gene and laid the foundation for studying its upstream regulation.
2. Results
2.1. Identification and Phylogenetic Analysis of AcGH3 Family Genes in Kiwifruit
To identify the GH3 family genes in kiwifruit, BLAST searches of the kiwifruit genome database (http://bioinfo.bti.cornell.edu/cgi-bin/kiwi/blast.cgi) were performed using the GH3 of the Arabidopsis and tomato proteins as a query sequence. A total of 12 GH3 genome DNA sequences were identified in A. chinensis; these genes were named as AcGH3.1-AcGH3.12. The names, IDs, open reading frame (ORF) lengths, position and deduced polypeptides of AcGH3 gene are shown in Table 1. The open reading frame (ORF) length of the AcGH3 gene varied from 1437 bp (AcGH3.1) to 2163 bp (AcGH3.5) and encoded 478–720aa peptides with a predicted molecular weight range of 54.29–81.26 kD. The theoretical pI ranged from 5.28 (AcGH3.4) to 8.53 (AcGH3.2) (Table 1). Except that AcGH3.5 and AcGH3.10 are distributed on chromosome 23, the other 10 GH3 members are distributed on different chromosomes. In addition, the chromosomal location of AcGH3.7 is unknown (Table 1).

Table 1.
The GH3 gene family in kiwifruit.
To investigate the relationship of the GH3 proteins, the full-length protein sequences of the 19 AtGH3s, 15 SlGH3s, and 12 AcGH3s were used to build the phylogenetic tree (Figure 1). The results indicated that the GH3 gene family could be grouped into three major subfamilies, I, II, and III. AcGH3.10–AcGH3.12 were grouped into subfamily I, the similarity of the three AcGH3 proteins is 43.9–93.2%. AcGH3.1–AcGH3.9 were grouped into subfamily III, the similarity of the nine AcGH3 proteins is between 35.9–93.5% (Figure 1; Table 2). No AcGH3 belonged to subfamily III. All 12 AcGH3s were distributed into 3 sister pairs of paralogous GH3s (AcGH3.2/AcGH3.3 (93.5%), AcGH3.6/AcGH3.7 (90.2%), AcGH3.11/AcGH3.12 (93.2%)), which had very strong bootstrap support (≥99), while the remaining AcGH3 were not matched (Figure 1; Table 2).
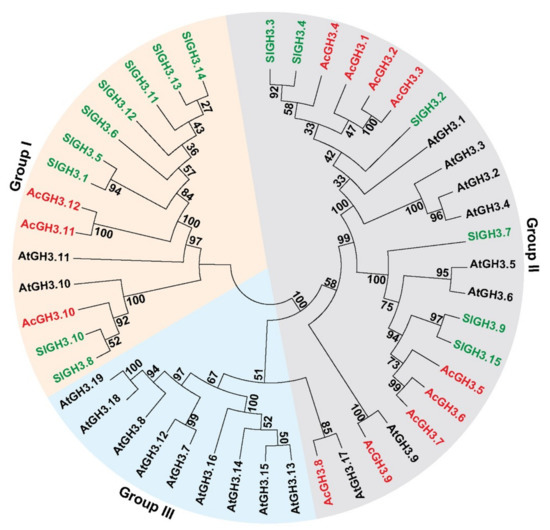
Figure 1.
Phylogenetic tree derived from the amino-acid sequences of 12 AcGH3 genes in kiwifruit and 15 SlGH3 genes in tomato, and 19 AtGH3 genes in Arabidopsis. All the GH3 genes are divided into three groups (I–III), and numbers near branches represent bootstrap values. The red, green and black fonts indicate GH3s from kiwifruit, tomato, and Arabidopsis, respectively.
Table 2.
Pairwise comparison of amino acid identity (%) of 12 AcGH3 proteins.
2.2. Expression Analysis of AcGH3 Gene Family
In order to clarify the function of the AcGH3 gene after harvest, we analyzed the changes of expression levels in seven different postharvest stages. In group I, AcGH3.10 is hardly expressed, AcGH3.11 and AcGH3.12 were the main expression members, the similarity between them is extremely high, and the expression trend is relatively consistent. They showed a decrease in expression level on day 0–9, followed by a rise (Figure 2). In group II, the CT values of AcGH3.4 and AcGH3.5 > 36, which hardly expressed. While AcGH3.2, AcGH3.3, AcGH3.6–AcGH3.9 showed very low expression levels in the fruit postharvest period. AcGH3.1 was the only gene exhibiting ripening-associated expression, its expression increased significantly, peaked at 15 days, and then declined at 18 days (Figure 2). Therefore, we suspect that AcGH3.1 may be related to the ripening process of the kiwifruit. We tested the response of AcGH3.1 gene expression to NAA (auxin analogue) and 1-MCP (ethylene receptor inhibitor), it was found that NAA can significantly promote the expression of AcGH3.1, while 1-MCP inhibits the increase of its expression (Figure 3).
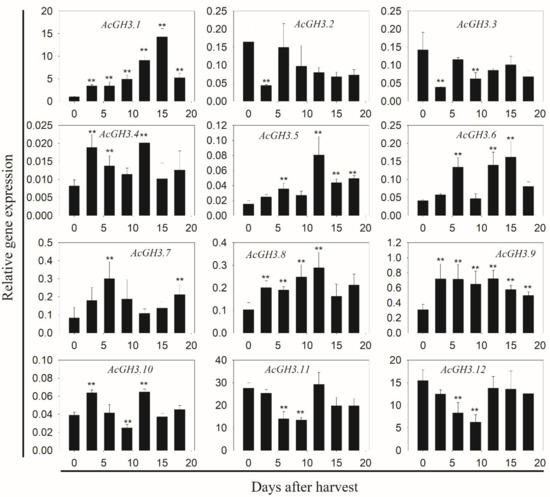
Figure 2.
q-PCR analysis of 12 AcGH3 genes in a total of seven postharvest stages. X-axis represents the different stages of postharvest. Y-axis represents the relative mRNA levels of genes. Expression of Actin was used as an internal control and to normalize the expression of AcGH3 genes. All values in the figure are relative to the AcGH3.1 value on day zero after harvest. Error bars show the standard error between three replicates performed. Asterisks indicate a significant difference relative to day zero after harvest according to a student’s t-test (** p < 0.01).
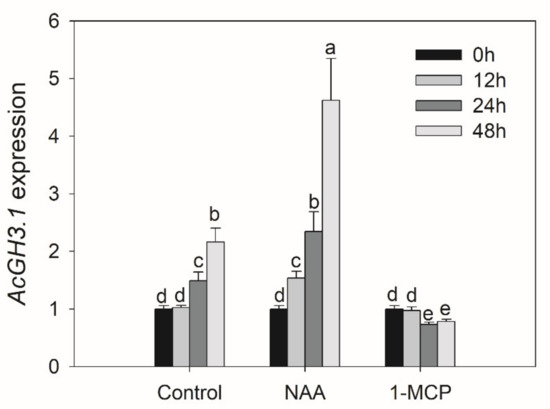
Figure 3.
Effects of NAA and 1-MCP treatment on AcGH3.1 gene expression in fruit after harvest. All values in the figure are relative to control value 0 h after harvest. Expression of Actin was used as an internal control and to normalize the expression of AcGH3.1 gene. Error bars show the standard error between three replicates performed. Bars with the same letter are not significantly different at the 0.01 level according to a student’s t-test.
2.3. Downregulation of the AcGH3.1 Gene Alter Fruit Maturation
To investigate the relationship of AcGH3.1 with fruit softening and ripening during postharvest, we generated the AcGH3.1-silenced construct by cloning the specific fragment (486 bp of the ORF) into the restriction site of Xba I and BamH I in the pTRV2 vector. Next, the purified plasmids of pTRV1, pTRV2, and pTRV2-AcGH3.1 were transformed into A. tumefaciens strain GV3101. The Agrobacterium strain was infiltrated into the kiwifruit pedicel about seven days before harvest. After this period, the fruits of pTRV1 + pTRV2-AcGH3.1 were collected. The expression level of AcGH3.1 was analyzed by qRT-PCR, its expression level in pTRV1+pTRV2-AcGH3.1 fruits was lower than that in control fruit throughout the storage period, and the decrease in firmness of pTRV1 + pTRV2-AcGH3.1 fruits was significantly inhibited (Figure 4). These results indicated that AcGH3.1 gene silencing delayed fruit softening.
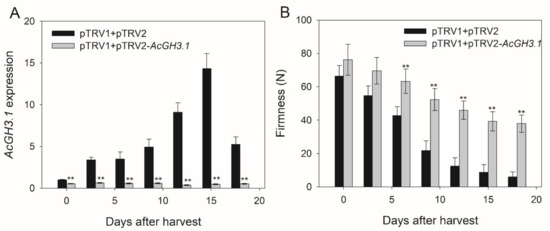
Figure 4.
Effects of AcGH3.1 silence on (A) AcGH3.1 expression and (B) firmness during kiwifruit postharvest storage. Expression of Actin was used as an internal control and to normalize the expression of AcGH3 genes. Error bars show the standard error between three replicates performed. Asterisks indicate a significant difference relative to pTRV1 + pTRV2 according to a student’s t-test (** p < 0.01).
2.4. Analysis of Cis-Acting Elements of AcGH3.1 Gene Promoter
PlantCARE was used to predict the upstream promoter elements of the gene and analyze the number of cis-acting elements related to hormones, light, and drought to further explore the functions of AcGH3.1. On the promoter of AcGH3.1, hormone-related action elements were relatively abundant, such as ABRE (abscisic acid (ABA) responsive element), ERE-motif (ethylene response factor binding site), TCA-element (salicylic acid (SA)-responsive element.), and TGA-element (auxin-responsive element) (Table 3). These plant hormones are reportedly involved in fruit ripening and softening, and can help us to better understand the transcriptional regulation of the AcGH3.1 gene.
Table 3.
Cis-elements were predicted in the promoter regions of AcGH3.1 related to hormone and abiotic stress in the kiwifruit.
Tobacco leaves are usually used to measure promoter activity. To further verify that AcGH3.1 expression was the above hormone inducible, fusion construct proAcGH3.1-GUS was transiently expressed in tobacco leaves and used to check the activities of the promoter (Figure 5A). After three days, we analyzed the effect of ABA, ethylene, SA, or NAA on proAcGH3.1-GUS, treated the transiently transformed tobacco leaves with ABA, ethylene, SA, or NAA, while tobacco leaves treated with H2O were used as controls. The determination of GUS enzyme activity revealed that GUS activity increased strikingly upon ABA, ethylene, or NAA treatment, and significantly decreased with SA treatment (Figure 5B). These observations suggested that AcGH3.1 expression is affected by ABA, ethylene, SA, or NAA, which might be closely related to fruit softening and ripening.
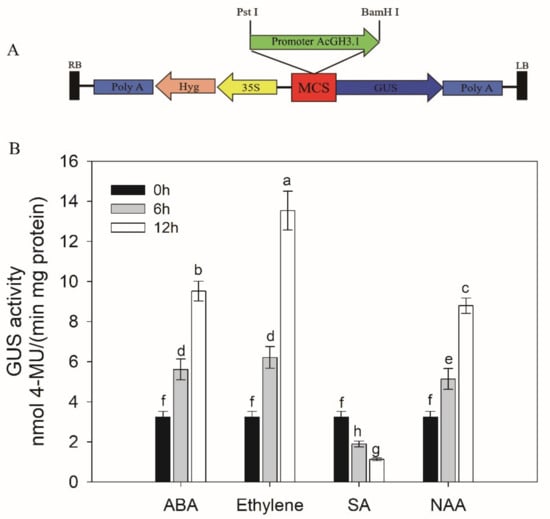
Figure 5.
(A) Schematic representation of AcGH3.1 promoter cloned in pCAMBIA1391 vector (promoter-less vector) at Pst I and BamH I sites for measuring GUS activity and field-infiltration; (B) Comparison of GUS activity determined in protein extracts (in vitro) treated with ABA, ethylene, SA, and NAA. Error bars show the standard error between three replicates performed. Bars with the same letter are not significantly different at the 0.01 level.
3. Discussion
As supported by an increasing amount of data, auxin plays an important role in the initiation of fruit ripening. The conjugation of IAA to amino acids, catalyzed by GH3 proteins, is an important aspect in the control of auxin homeostasis in plants [3,4,14]. It is necessary to study the transcription and function of members of this gene family, so that the information could be used for understand the auxin metabolism during fruit development and ripening. The first GH3 gene was isolated from soybean by differential screening after auxin treatment [8]. Nineteen GH3 genes were identified from Arabidopsis and 12 from rice [5,20]. Since then, several members of the gene family have been identified in many plant species, such as, grape, orange, tomato, papaya, etc. [15,18,21,22]. Here, 12 GH3 genes were identified from the kiwifruit genome. In the combined phylogenetic tree of Arabidopsis, tomato, and kiwifruit, all the GH3 proteins were found to cluster in three groups. AcGH3 is all grouped into subfamily I and II, and absence of any AcGH3 gene in group III. Presence of one sister pairing (AcGH3.9–AtGH3.9, 100%) in the combined phylogenetic tree, indicating a close relationship between the Arabidopsis and kiwifruit GH3 gene families, these genes might have descended from a common ancestor and can have conserved functions [23].
Gene expression analysis indicated that the GH3 gene has different expression patterns and function in different plants [16,24,25]. Two Arabidopsis GH3 proteins, DFL1 and YDK1, negatively regulate shoot bud cell elongation and lateral root formation, overexpressing GH3-2/YDK1 and GH3-6/DFL1 mutants showed a dwarfing phenotype consistent with these reduced levels of free IAA [26,27]. The AtGH3-9 gene is involved in the regulation of the primary root growth of Arabidopsis, the overexpressed strains of GH3-8 showed decreased IAA level and increased IAA-Asp level, abnormal development morphology and growth retardation [28]. Overexpression of a rice GH3 gene (GH3.13) related to drought adaptation showed a dwarfing phenotype with increased tiller number and leaf angle [29]. The expression levels of the ancestral species GH3 genes changed obviously after allopolyploidization in Brassica napus [30]. During fruit development and maturation, GH3 gene also showed different expression patterns. When IAA content was high, GH3-1 gene was highly expressed in grape flowers and young fruits [31,32], whereas relatively low in roots and leaves [15]. GH3.1/GH3-2 involved in the fruit ripening of grape berry [6,15]. CcGH3 expression in shoots, buds, sepals, petals, and mature pericarp and placenta could be induced by auxin [17]. In longan fruit, DlGH3.2 was believed to be involved in pericarp growth and fruit ripening [16]. A comprehensive expression profiling of auxin-related GH3 genes in tomato, the expression levels of SlGH3-1 and SlGH3-2 increased at the early stage of fruit ripening, indicating that they may be involved in the initiation of fruit ripening [18]. PpGH3-3 and PpGH3-4 were significantly up-regulated in the mature stage, suggesting that they may be involved in the ripening and softening of peach fruit [33]. PuGH3.1 plays an important role in the decrease of free IAA content in pear fruits, which is closely related to ethylene production and IAA content [7]. In our study, the expression of AcGH3.1 shared high correlation with fruit postharvest ripening, and was induced by NAA and inhibited by 1-MCP. After transient silencing of its expression, fruit ripening could be delayed, suggesting that the AcGH3.1 gene was involved in fruit ripening after harvest. This also provides a new idea for us to study the effect of IAA on the postharvest maturity of kiwifruit.
Further studies have shown that the expression of the GH3 gene is affected by many other factors. Several endogenous and exogenous factors are currently known, including auxin (IAA), abscisic acid (ABA), ethylene, brassinolide (BL), gibberellin (GA), jasmonic acid (JA), salicylic acid (SA), light and biotic/abiotic stress can regulate the expression of GH3 gene in plants [5,6,9,14,15,20,24,26,29,34,35]. The positive regulation of auxin is a common feature of group II GH3 genes [5], and the presence of a core sequence (TGTCTC) of many auxin response elements (AuxRE) on the GH3-1 gene promoter illustrates this problem [36]. GH3-1 expression in grapes can be induced by ABA or sucrose treatment, and the induction was amplified by the combination of the both, meanwhile, the presence of multiple ABA response elements (ABRE-like) in the GH3-1 promoter region was predicted [15]. ABA and ethylene promote the expression of DlGH3.2 during the ripening stage of longan fruits [16]. In peppers and pears, the expression level of GH3 gene is strongly induced by ethylene, but is inhibited by 1-MCP [7,17]. Some GH3 genes, such as PBS3 (also known as AtGH3.12) and AtGH3.5, play key roles in regulating salicylic acid metabolism and inducing defense responses, suggesting that these genes can be regulated by other plant growth substances [35,37]. Transcriptional levels of GH3.8 in rice can be rapidly induced by salicylic acid and jasmonic acid treatment [28]. In addition, AtGH3.5 expression is induced by abiotic stresses such as cold, salt, drought and ABA [38], and cadmium was found to induce the up-regulation of some GH3 genes in mustard [39]. Several types of transcription factors such as ARF (auxin response factor), bZIP (basic region-leucine zipper) and R2R3 type MYB (v-myb avian myeloblastosis viral oncogene homolog) have also been reported to effect the expression of the GH3 gene in different plants [40,41,42]. When analyzing the AcGH3.1 promoter sequence, we found that there are many cis-elements related to plant hormones in the promoter sequence, such as ABRE (abscisic acid responsive element), ERE-motif (ethylene response factor binding site), TCA-element (salicylic acid-responsive element.), and TGA-element (auxin-responsive element), which have been proved to play an important role in fruit ripening [43,44,45,46]. In addition, we found that the activity of the promoter of the AcGH3.1 gene is affected by these exogenous hormones, which laid the foundation for our study of its upstream regulation.
4. Materials and Methods
4.1. Plant Materials and Treatments
Kiwifruits (A. chinensis, Donghong) were collected at the commercially mature stage from the Institute of Kiwifruit Research in Fengxin County, Jiangxi Province, China. Fruit samples were divided into three groups; each treatment contained three biological replicates, each consisting of approximately 200 fruits. The first group was treated with 50 mg/L 1-naphthylacetic acid (NAA), the second group was treated with 1 μL/L 1-methylcyclopropene (1-MCP), and the third group was treated with air as a control. All fruits were enclosed in 0.02-mm thick polyethylene film bags to maintain relative humidity at nearly 95%, and then stored at 20 °C. At each sampling point, 3 replicates of 4 fruits each were collected from each treatment. The outer pericarp (without skin or seeds) of control fruits was separated at 0, 12, 24, 48 h, and 3, 6, 9, 12, 15, 18 d of storage, rapidly frozen in liquid nitrogen and kept at −80 °C until use.
4.2. Identification of the GH3 Gene from Kiwifruit Genome and Phylogenetic Analysis
Candidate genes encoding GH3 were retrieved by BLASTP searching against the kiwifruit genome database (http://bioinfo.bti.cornell.edu/cgi-bin/kiwi/home.cgi), using tomato/Arabidopsis GH3 proteins as queries. Phylogenetic analysis was conducted using the ClustalW in software MEGA 7.0 (Arizona State University, Tempe, AZ, USA), and an unrooted phylogenetic tree of the GH3 gene families was constructed using the software MEGA 7.0 [47]. Evolutionary history was inferred using the neighbor-joining method with 1000 replicates.
4.3. RNA Extraction and qRT-PCR
Total RNA was extracted using a plant RNA extraction kit (Huayueyang, Beijing, China). DNA contamination in the isolated RNA was digested by incubation with DNase I (Takara, Dalian, China) for 30 min at 37 °C before cDNA was synthesized; 1 μg RNA was used for cDNA synthesis using the Verso cDNA kit (Takara, Dalian, China); qRT-PCR was performed using a CFX96 Touch Real-time PCR (Polymerase Chain Reaction) instrument (Bio-RAD, Hercules, CA, USA) with 2 μL cDNA, 1 X TB Green Master Mix (5 μL) (Takara, Dalian, China), and 1 μM each of two gene-specific primers (Table S1), with a final volume of 10 μL in water. The thermal cycle scheme is as follows: 30 s at 95 °C, 40 cycles of 5 s at 95 °C, and 34 s at 60 °C. The Actin gene was used as internal reference. The relative expression of gene was calculated by the comparative cycle threshold method (△△Ct). Three biological replicates and three technical replicates were performed to verify the accuracy of expression data.
4.4. AcGH3.1-Silenced Kiwifruit
A 486 bp fragment of the AcGH3.1 gene was amplified by PCR from kiwifruit cDNA sources using gene specific primes GCTCTAGATTGAACCGCAATGGAACAGT (forward) and CGGGATCCTGATCCGATCCCAATTCTCAC (reverse) (Xba I and BamH I sites are underlined). The product of PCR was cloned into pTRV2 (posttranscriptional Tobacco Rattle Virus) at Xba I and BamH I restriction sites to form pTRV2-AcGH3.1. The purified plasmids of pTRV1, pTRV2, and pTRV2-AcGH3.1 were transformed into Agrobacterium tumefaciens strain GV3101. Finally, A. tumefacines cells were resuspended with infiltration media (10 mM MgCl2, 10 mM MES (2-Morpholinoethanesulfonic Acid monohydrate), 200 μM AS (Acetosyringon)), adjusted to OD600 > 1.0. The liquid culture contained pTRV1 and pTRV2 (as a control) or pTRV1 and pTRV2-AcGH3.1 in a 1:1 ratio added and was cultured at 28 °C for 3 h. After this time, the liquid culture was infiltrated into the fruit pedicel approximately 7 d before harvest, after which, the AcGH3.1-silenced fruits were collected. Fruit firmness was measured as previously described [48], it was measured on opposite sides of each fruit after peel removal (1 mm thick), using a fruit texture analyzer (FTA, model GS, Güss Manufacturing Ltd., Strand, South Africa) with a 10-mm probe. Data were recorded as Newton (N) and fruit firmness was expressed as the mean of 15–20 fruits.
4.5. Cis-Elements Analysis
To investigate cis-elements in AcGH3.1 gene promoter region, sequence of the 1287 kb region upstream of the start codons (ATG) was extracted from the kiwifruit genome database, and the gene promoter elements were predicted and analyzed using the PlantCARE online site (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) [49].
4.6. Promoter Activity Assay
The putative promoter region of AcGH3.1, a 1287 bp PCR fragment upstream of the start codon ATG, was amplified from kiwifruit genomic DNA using the primers AACTGCAGCTTTTATATATATATATATAT (forward) and CGGGATCCATATGAAGTGGTCCCCAATT (reverse) (Pst I and BamH I sites are underlined). The PCR product was digested with Pst I and BamH I was cloned in front of the GUS gene in the pCAMBIA1391 vector (promoter less vector), yielding the construct proAcGH3.1-GUS. The AcGH3.1 promoter cloned in pCAMBIA1391 was transformed in Agrobacterium tumefaciens (GV3101). Then, A. tumefaciens containing the proAcGH3.1-GUS construct standing 3 h at room temperature. 500 μL agrobacterium was injected into the abaxial surface of the fully expanded tobacco leaves with a 1 mL syringe. After 3 days, different treatments were performed on the leaves. Agrobacterium-infected leaves were incubated at room temperature for 0, 6 or 12 h in petri dishes containing 10 μM abscisic acid (ABA), 10 μM ethephon, 10 Μm salicylic acid (SA) or 10 μM NAA, respectively. GUS staining was carried out as previously described [50].
Supplementary Materials
The following are available online at https://www.mdpi.com/2223-7747/8/11/473/s1, Table S1: Primers for qRT-PCR.
Author Contributions
Z.G. designed and conducted the experiments, analyzed the data, accomplished pictures and wrote the manuscript. N.S., L.F., Y.F. contributed to the design of the experiments. J.C. conceived and designed the experiments, and finalized the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by the Natural Science Foundation Project of Jiangxi Education Department (NO. GJJ13255), National Natural Science Foundation of China (NO. 31760598), and Natural Science Foundation and Advantage Innovation Team Project of Jiangxi Province (NO. 20171BAB214031).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Fang, L.; Zhang, S.; Sross, G. Role of abscisic acid and indole-3-acetic acid in kiwifruit ripening. Acta Hortic. Sin. 1999, 26, 81–86. [Google Scholar]
- Delker, C.; Raschke, A.; Quint, M. Auxin dynamics: The dazzling complexity of a small molecule’s message. Planta 2008, 227, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef]
- Staswick, P.E.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 2005, 17, 616–627. [Google Scholar] [CrossRef]
- Boettcher, C.; Boss, P.K.; Davies, C. Acyl substrate preferences of an IAA-amido synthetase account for variations in grape (Vitis vinifera L.) berry ripening caused by different auxinic compounds indicating the importance of auxin conjugation in plant development. J. Exp. Bot. 2011, 62, 4267–4280. [Google Scholar] [CrossRef]
- Yue, P.; Wang, Y.; Bu, H.; Li, X.; Yuan, H.; Wang, A. Ethylene promotes IAA reduction through PuERFs-activated PuGH3.1 during fruit ripening in pear (Pyrus ussuriensis). Postharvest Biol. Technol. 2019, 157, 110955. [Google Scholar] [CrossRef]
- Hagen, G.; Kleinschmidt, A.; Guilfoyle, T. Auxin-regulated gene-expression in intact soybean hypocotyl and excised hypocotyl sections. Planta 1984, 162, 147–153. [Google Scholar] [CrossRef]
- Domingo, C.; Andres, F.; Tharreau, D.; Iglesias, D.J.; Talon, M. Constitutive expression of OsGH3.1 reduces auxin content and enhances defense response and resistance to a fungal pathogen in rice. Mol. Plant 2009, 22, 201–210. [Google Scholar]
- Ludwig-Mueller, J. Auxin conjugates: Their role for plant development and in the evolution of land plants. J. Exp. Bot. 2011, 62, 1757–1773. [Google Scholar] [CrossRef]
- Staswick, P. Plant hormone conjugation. Plant Signal. Behav. 2009, 4, 757–759. [Google Scholar] [CrossRef] [PubMed]
- Okrent, R.A.; Wildermuth, M.C. Evolutionary history of the GH3 family of acyl adenylases in rosids. Plant Mol. Biol. 2011, 76, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Terol, J.; Domingo, C.; Talon, M. The GH3 family in plants: Genome wide analysis in rice and evolutionary history based on EST analysis. Gene 2006, 371, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Bai, Y.; Shen, C.; Wu, Y.; Zhang, S.; Jiang, D.; Guilfoyle, T.J.; Chen, M.; Qi, Y. Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genomics 2010, 10, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Boettcher, C.; Keyzers, R.A.; Boss, P.K.; Davies, C. Sequestration of auxin by the indole-3-acetic acid-amido synthetase GH3-1 in grape berry (Vitis vinifera L.) and the proposed role of auxin conjugation during ripening. J. Exp. Bot. 2010, 61, 3615–3625. [Google Scholar] [CrossRef] [PubMed]
- Kuang, J.; Zhang, Y.; Chen, J.; Chen, Q.; Jiang, Y.; Lin, H.; Xu, S.; Lu, W. Two GH3 genes from longan are differentially regulated during fruit growth and development. Gene 2011, 485, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.D.; Kang, B.C.; Jiang, H.; Moore, S.L.; Li, H.X.; Watkins, C.B.; Setter, T.L.; Jahn, M.M. A GH3-like gene, CcGH3, isolated from Capsicum chinense L. fruit is regulated by auxin and ethylene. Plant Mol. Biol. 2005, 58, 447–464. [Google Scholar] [CrossRef]
- Kumar, R.; Agarwal, P.; Tyagi, A.K.; Sharma, A.K. Genome-wide investigation and expression analysis suggest diverse roles of auxin-responsive GH3 genes during development and response to different stimuli in tomato (Solanum lycopersicum). Mol. Genet. Genomics 2012, 287, 221–235. [Google Scholar] [CrossRef]
- Panu, A.; Manohar, J.; Konstantin, A.; Delphine, B.; Gabor, C.; Edouard, d.C.; Séverine, D.; Volker, F.; Arnaud, F.; Elisabeth, G.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, E597–W603. [Google Scholar]
- Jain, M.; Kaur, N.; Tyagi, A.K.; Khurana, J.P. The auxin-responsive GH3 gene family in rice (Oryza sativa). Funct. Integr. Genomics 2006, 6, 36–46. [Google Scholar] [CrossRef]
- Liu, K.; Wang, J.; Li, H.; Zhong, J.; Feng, S.; Pan, Y.; Yuan, C. Identification, expression and iaa-amide synthetase activity analysis of Gretchen Hagen 3 in Papaya fruit (Carica papaya L.) during postharvest process. Front. Plant Sci. 2016, 7, 1555. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Pang, S.; Ma, Y.; Deng, L.; He, S.; Yi, S.; Lv, Q.; Zheng, Y. The ARF, AUX/IAA and GH3 gene families in citrus: Genome-wide identification and expression analysis during fruitlet drop from abscission zone A. Mol. Genet. Genomics 2015, 290, 2089–2105. [Google Scholar] [CrossRef] [PubMed]
- Rahul, K.; Tyagi, A.K.; Sharma, A.K. Genome-wide analysis of auxin response factor (ARF) gene family from tomato and analysis of their role in flower and fruit development. Mol. Genet. Genomics 2011, 285, 245–260. [Google Scholar]
- Khan, S.; Stone, J.M. Arabidopsis thaliana GH3.9 in auxin and jasmonate cross talk. Plant Signal. Behav. 2007, 2, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Tomoyuki, T.; Miki, N.; Akie, I.; Katsushi, M.; Minami, M. DFL2, a new member of the Arabidopsis GH3 gene family, is involved in red light-specific hypocotyl elongation. Plant Cell Physiol. 2003, 44, 1071–1080. [Google Scholar]
- Nakazawa, M.; Yabe, N.; Ichikawa, T.; Yamamoto, Y.Y.; Yoshizumi, T.; Hasunuma, K.; Matsui, M. DFL1, an auxin-responsive GH3 gene homologue, negatively regulates shoot cell elongation and lateral root formation, and positively regulates the light response of hypocotyl length. Plant J. 2001, 25, 213–221. [Google Scholar] [CrossRef]
- Tomoyuki, T.; Miki, N.; Akie, I.; Mika, K.; Takanari, I.; Naoki, T.; Hiroaki, S.; Katsushi, M.; Minami, M. ydk1-D, an auxin-responsive GH3 mutant that is involved in hypocotyl and root elongation. Plant J. 2010, 37, 471–483. [Google Scholar]
- Ding, X.; Cao, Y.; Huang, L.; Zhao, J.; Xu, C.; Li, X.; Wang, S. Activation of the indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes salicylate- and jasmonate-independent basal immunity in rice. Plant Cell 2008, 20, 228–240. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Cao, J.; Zhang, Y.; Zhang, S.; Xia, Y.; Sun, D.; Sun, Y. Altered architecture and enhanced drought tolerance in rice via the down-regulation of indole-3-acetic acid by TLD1/OsGH3.13 activation. Plant Physiol. 2009, 151, 1889–1901. [Google Scholar] [CrossRef]
- Wang, R.; Li, M.; Wu, X.; Wang, J. The Gene Structure and Expression Level Changes of the GH3 Gene Family in Brassica napus Relative to Its Diploid Ancestors. Genes 2019, 10, 58. [Google Scholar] [CrossRef]
- Deytieux-Belleau, C.; Gagne, S.; L’Hyvernay, A.; Doneche, B.; Geny, L. Possible roles of both abscisic acid and indol-acetic acid in controlling grape berry ripening process. J. Int. Sci. Vigne Vin 2007, 41, 141–148. [Google Scholar] [CrossRef]
- Zhang, X.R.; Luo, G.G.; Wang, R.H.; Wang, J.; Himelrick, D.G. Growth and developmental responses of seeded and seedless grape berries to shoot girdling. J. Am. Soc. Hortic. Sci. 2003, 128, 316–323. [Google Scholar] [CrossRef]
- Zeng, W.; Pan, L.; Niu, L.; Lu, Z.; Cui, G.; Wang, Z. Bioinformatics Analysis and Expression of the Nectarine Indole-3-aceticacid- amido Synthase(GH3)Gene Family During Fruit Development. Acta Hortic. Sin. 2015, 42, 833–842. [Google Scholar]
- Jagadeeswaran, G.; Raina, S.; Acharya, B.R.; Maqbool, S.B.; Mosher, S.L.; Appel, H.M.; Schultz, J.C.; Klessig, D.F.; Raina, R. Arabidopsis GH3-LIKE DEFENSE GENE 1 is required for accumulation of salicylic acid, activation of defense responses and resistance to pseudomonas syringae. Plant J. 2007, 51, 234–246. [Google Scholar] [CrossRef]
- Zhang, Z.; Qun, L.; Zhimiao, L.; Staswick, P.E.; Muyang, W.; Ying, Z.; Zuhua, H. Dual regulation role of GH3.5 in salicylic acid and auxin signaling during Arabidopsis-pseudomonas syringae interaction. Plant Physiol. 2007, 145, 450–464. [Google Scholar] [CrossRef]
- Ulmasov, T.; Liu, Z.B.; Hagen, G.; Guilfoyle, T.J. Composite structure of auxin response elements. Plant Cell 1995, 7, 1611–1623. [Google Scholar]
- Nobuta, K.; Okrent, R.A.; Stoutemyer, M.; Rodibaugh, N.; Kempema, L.; Wildermuth, M.C.; Innes, R.W. The GH3 acyl adenylase family member PBS3 regulates salicylic acid-dependent defense responses in Arabidopsis. Plant Physiol. 2007, 144, 1144–1156. [Google Scholar] [CrossRef]
- Park, J.E.; Park, J.Y.; Kim, Y.S.; Staswick, P.E.; Jeon, J.; Yun, J.; Kim, S.Y.; Kim, J.; Lee, Y.H.; Park, C.M. GH3-mediated auxin homeostasis links growth regulation with stress adaptation response in Arabidopsis. J. Biol. Chem. 2007, 282, 10036–10046. [Google Scholar] [CrossRef]
- Minglin, L.; Zhang, Y.X.; Chai, T.Y. Identification of genes up-regulated in response to Cd exposure, in Brassica juncea L. Gene 2005, 363, 151–158. [Google Scholar] [CrossRef]
- Heinekamp, T.; Strathmann, A.; Kuhlmann, M.; Froissard, M.; Muller, A.; Perrot-Rechenmann, C.; Droge-Laser, W. The tobacco bZIP transcription factor BZI-1 binds the GH3 promoter in vivo and modulates auxin-induced transcription. Plant J. 2004, 38, 298–309. [Google Scholar] [CrossRef]
- Seo, P.J.; Park, C. Auxin homeostasis during lateral root development under drought condition. Plant Signal. Behav. 2009, 4, 1002–1004. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Muto, H.; Higuchi, K.; Matamura, T.; Tatematsu, K.; Koshiba, T.; Yamamoto, K.T. Disruption and overexpression of auxin response factor 8 gene of Arabidopsis affect hypocotyl elongation and root growth habit, indicating its possible involvement in auxin homeostasis in light condition. Plant J. 2004, 40, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Jiang, Z.; Zhang, L.; Tan, D.; Wei, Y.; Yuan, H.; Li, T.; Wang, A. Apple (Malus domestica) MdERF2 negatively affects ethylene biosynthesis during fruit ripening by suppressing MdACS1 transcription. Plant J. 2016, 88, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Mou, W.; Li, D.; Luo, Z.; Li, L.; Mao, L.; Ying, T. SlAREB1 transcriptional activation of NOR is involved in abscisic acid-modulated ethylene biosynthesis during tomato fruit ripening. Plant Sci. 2018, 276, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Sagar, M.; Chervin, C.; Mila, I.; Hao, Y.; Roustan, J.; Benichou, M.; Gibon, Y.; Biais, B.; Maury, P.; Latche, A.; et al. SlARF4, an Auxin Response Factor Involved in the Control of Sugar Metabolism during Tomato Fruit Development. Plant Physiol. 2013, 161, 1362–1374. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.S.; Zhang, S.L.; Ferguson, I. The role of salicylic acid in postharvest ripening of kiwifruit. Postharvest Biol. Technol. 2003, 28, 67–74. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, T.; Qin, G.; Li, B.; Tian, S. Synergistic action of antioxidative systems contributes to the alleviation of senescence in kiwifruit. Postharvest Biol. Technol. 2016, 111, 15–24. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions—Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher-plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).