Unraveling Morphophysiological and Biochemical Responses of Triticum aestivum L. to Extreme pH: Coordinated Actions of Antioxidant Defense and Glyoxalase Systems

 ,
,  ,
, 

Abstract
1. Introduction
2. Results
2.1. Growth and Biomass Accumulation
2.2. Relative Water and Proline Content
2.3. Oxidative Stress Markers
2.4. Photosynthetic Pigment Contents
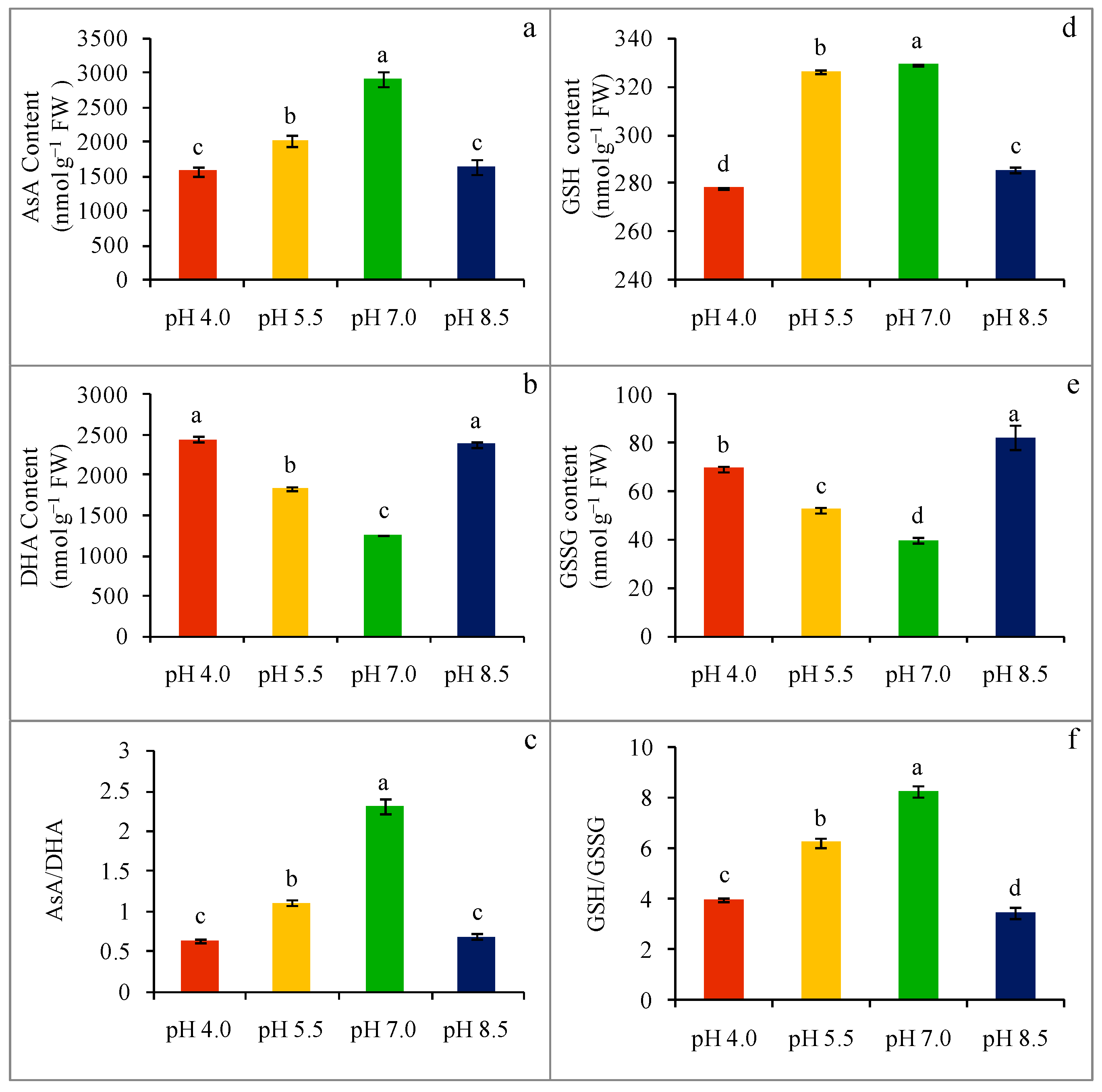
2.5. Nonenzymatic Antioxidant Content
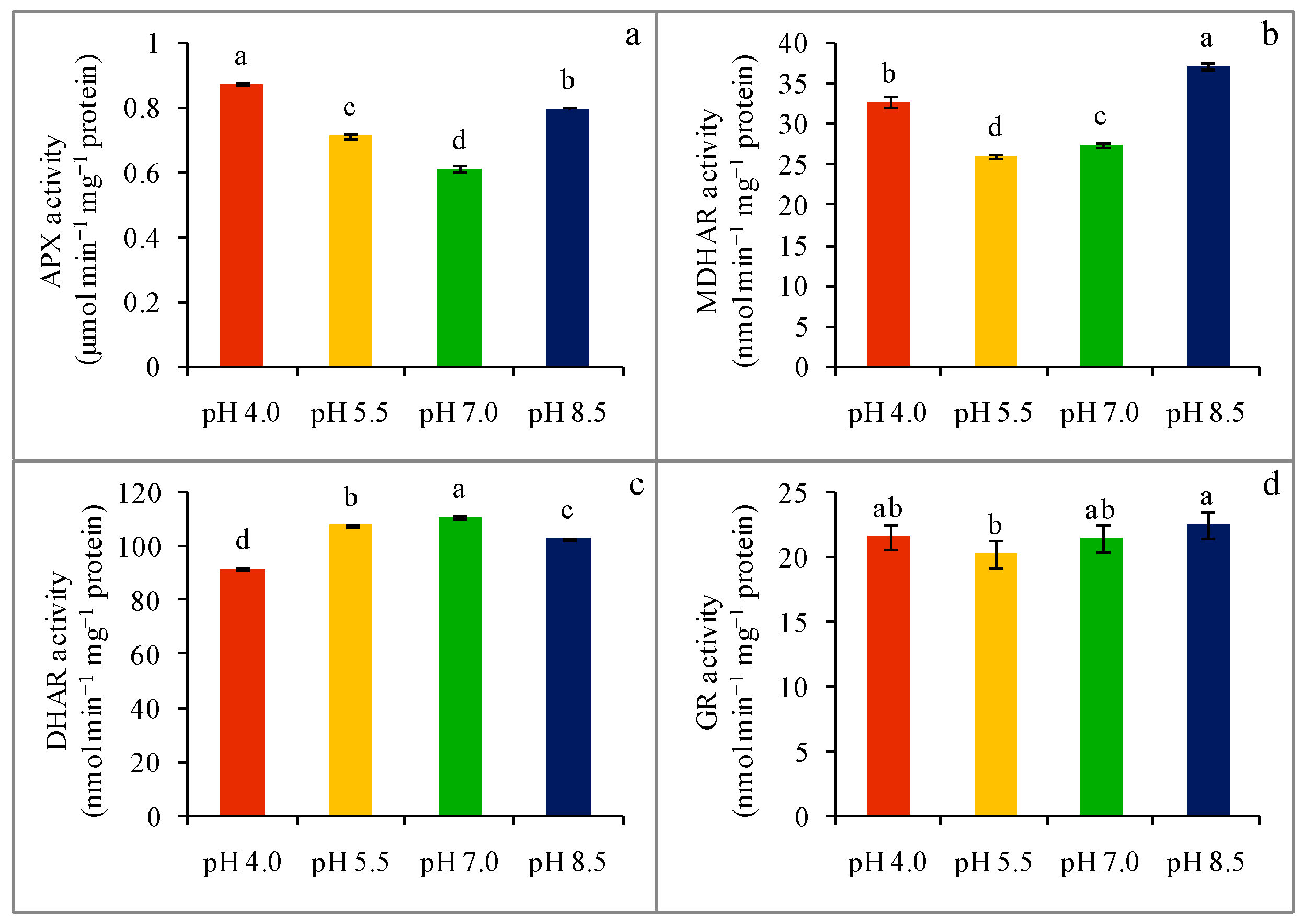
2.6. Enzyme Activity
2.7. Glyoxalase Enzymes Activity and Methylglyoxal Content
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Stress Treatments
4.2. Growth and Biomass Accumulation
4.3. Determination of Stress Markers
4.4. Photosynthetic Pigment Contents
4.5. Relative Water Content (RWC) and Proline Content
4.6. Nonenzymatic Antioxidant Assay
4.7. Protein Determination and Enzyme Activity Assay
4.8. Glyoxalase Activity Assay and Methylglyoxal Content
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fraire-Velázquez, S.; Balderas-Hernández, V.E. Abiotic stress in plants and metabolic responses. In Abiotic Stress-Plant Responses and Applications in Agriculture; Vahdati, K., Leslie, C., Eds.; InTech: London, UK, 2013; pp. 25–48. [Google Scholar]
- Merry, R.H. Acidity and alkalinity of soils. In Environmental and Ecological Chemistry-volume II; Sabljic, A., Ed.; Eolss Publications: Oxford, UK, 2009; pp. 115–131. [Google Scholar]
- Soil Survey Division Staff. Examination and Description of Soil Profiles. In Soil survey manual, US Department of Agriculture Handbook 18; Soil Survey Division Staff, Ed.; US Department of Agriculture: Washington DC, USA, 2017; pp. 83–230. [Google Scholar]
- Albert, S. The Kitchen Garden Grower’s Guide: A Practical Vegetable and Herb Garden Encyclopedia; Create Space Independent Pub: Seattle, WA, USA, 2008. [Google Scholar]
- Zheng, S.J. Crop production on acidic soils: overcoming aluminium toxicity and phosphorus deficiency. Ann. Bot. 2010, 106, 183–184. [Google Scholar] [CrossRef] [PubMed]
- Shavrukov, Y.; Hirai, Y. Good and bad protons: genetic aspects of acidity stress responses in plants. J. Exp. Bot. 2015, 67, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, R.E.; Duncan, R.R. Sorghum seedling growth as influenced by H+, Ca2+, and Mn2+ concentrations. J. Plant Nutr. 1989, 12, 1379–1394. [Google Scholar] [CrossRef]
- Yang, C.W.; Wang, P.; Li, C.Y.; Shi, D.C.; Wang, D.L. Comparison of effects of salt and alkali stresses on the growth and photosynthesis of wheat. Photosynthetica 2008, 46, 107–114. [Google Scholar] [CrossRef]
- Yang, Y.; Qin, Y.; Xie, C.; Zhao, F.; Zhao, J.; Liu, D.; Chen, S.; Fuglsang, A.T.; Palmgren, M.G.; Schumaker, K.S.; et al. The Arabidopsis chaperone J3 regulates the plasma membrane H+-ATPase through interaction with the PKS5 kinase. Plant Cell 2010, 22, 1313–1332. [Google Scholar] [CrossRef] [PubMed]
- Joliot, P.; Johnson, G.N. Regulation of cyclic and linear electron flow in higher plants. Proc. Natl. Acad. Sci. USA 2011, 108, 13317–13322. [Google Scholar] [CrossRef]
- Song, H.; Xu, X.; Wang, H.; Tao, Y. Protein carbonylation in barley seedling roots caused by aluminum and proton toxicity is suppressed by salicylic acid. Russ. J. Plant Physiol. 2011, 58, 653–659. [Google Scholar] [CrossRef]
- Zhang, K.X.; Wen, T.; Dong, J.; Ma, F.W.; Bai, T.H.; Wang, K.; LI, C.Y. Comprehensive evaluation of tolerance to alkali stress by 17 genotypes of apple rootstocks. J. Integr. Agric. 2016, 15, 1499–1509. [Google Scholar] [CrossRef]
- Ivanov, Y.; Savochkin, Y.; Kuznetsov, V. Effect of mineral composition and medium pH on Scots pine tolerance to toxic effect of zinc ions. Russ. J. Plant Physiol. 2013, 60, 260–269. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Hossain, M.S.; Anee, T.I.; Parvin, K.; Fujita, M. Nitric oxide pretreatment enhances antioxidant defense and glyoxalase systems to confer PEG-induced oxidative stress in rapeseed. J. Plant Interact. 2017, 12, 323–331. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Exogenous selenium pretreatment protects rapeseed seedlings from cadmium-induced oxidative stress by upregulating antioxidant defense and methylglyoxal detoxification systems. Biol. Trace Elem. Res. 2012, 149, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.; Fujita, M. Hydrogen peroxide pretreatment mitigates cadmium-induced oxidative stress in Brassica napus L.: An intrinsic study on antioxidant defense and glyoxalase systems. Front. Plant Sci. 2017, 8, 115. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Coordinated actions of glyoxalase and antioxidant defense systems in conferring abiotic stress tolerance in plants. Int. J. Mol. Sci. 2017, 18, 200. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Suzuki, T.; Fujita, M. Polyamines-induced aluminum tolerance in mung bean: A study on antioxidant defense and methylglyoxal detoxification systems. Ecotoxicology 2017, 26, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Viveros, M.F.Á.; Inostroza-Blancheteau, C.; Timmermann, T.; González, M.; Arce-Johnson, P. Overexpression of GlyI and GlyII genes in transgenic tomato (Solanum lycopersicum Mill.) plants confers salt tolerance by decreasing oxidative stress. Mol. Biol. Rep. 2013, 40, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, J.A.; Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Hossain, M.S.; Fujita, M. Maleic acid assisted improvement of metal chelation and antioxidant metabolism confers chromium tolerance in Brassica juncea L. Ecotoxicol. Environ. Saf. 2017, 144, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Kochian, L.V.; Piñeros, M.A.; Liu, J.; Magalhães, J.V. Plant adaptation to acid soils: the molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef]
- Yang, C.; Xu, H.; Wang, L.; Liu, J.; Shi, D.; Wang, D. Comparative effects of salt stress and alkali-stress on the growth, photosynthesis, solute accumulation, and ion balance of barley plants. Photosynthetica 2009, 47, 79–86. [Google Scholar] [CrossRef]
- Wang, X.; Geng, S.; Ri, Y.J.; Cao, D.; Liu, J.; Shi, D.; Yang, C. Physiological responses and adaptive strategies of tomato plants to salt and alkali stresses. Sci. Hort. 2011, 130, 248–255. [Google Scholar] [CrossRef]
- Marschner, H. Mechanisms of adaptation of plants to acid soils. Plant Soil 1991, 134, 1–20. [Google Scholar] [CrossRef]
- Silva, C.M.S.; Zhang, C.; Habermann, G.; Delhaize, E.; Ryan, P.R. Does the major aluminium-resistance gene in wheat, TaALMT1, also confer tolerance to alkaline soils? Plant Soil 2018. [Google Scholar] [CrossRef]
- Gong, B.; Li, X.; Vanden Langenberg, K.M.; Wen, D.; Sun, S.; Wei, M.; Li, Y.; Yang, F.; Shi, Q.; Wang, X. Overexpression of S-adenosyl-l-methionine synthetase increased tomato tolerance to alkali stress through polyamine metabolism. Plant Biotechnol. J. 2014, 12, 694–708. [Google Scholar] [CrossRef] [PubMed]
- Pal’ove-Balang, P.; Čiamporová, M.; Zelinová, V.; Pavlovkin, J.; Gurinová, E.; Mistrík, I. Cellular responses of two Latin-American cultivars of Lotus corniculatus to low pH and Al stress. Cent. Eur. J. Biol. 2012, 7, 1046–1054. [Google Scholar]
- Zhang, Y.K.; Zhu, D.F.; Zhang, Y.P.; Chen, H.Z.; Xiang, J.; Lin, X.Q. Low pH-induced changes of antioxidant enzyme and ATPase activities in the roots of rice (Oryza sativa L.) seedlings. PloS ONE 2015, 10, e0116971. [Google Scholar] [CrossRef] [PubMed]
- Long., A.; Zhang, J.; Yang, L.T.; Ye, X.; Lai, N.W.; Tan, L.L.; Lin, D.; Chen, L.S. Effects of ow pH on photosynthesis, related physiological parameters, and nutrient profiles of Citrus. Front Plant Sci. 2017, 8, 185. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.H.; Yang, C.W.; Yang, M.Y. Photosynthesis, photosystem II efficiency, amino acid metabolism and ion distribution in rice (Oryza sativa L.) in response to alkaline stress. Photosynthetica 2014, 52, 157–160. [Google Scholar] [CrossRef]
- Amist, N.; Singh, N.B. Responses of enzymes involved in proline biosynthesis and degradation in wheat seedlings under stress. Allelopathy J. 2017, 42, 195–205. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Alteration in chlorophylls and carotenoids in higher plants under abiotic stress condition. In Photosynthetic Pigments: Chemical Structure, Biological Function And Ecology; Golovko, T.K., Gruszecki, W.I., Prasad, M.N.V., Strzałka, K., Eds.; Komi Scientific Centre of the Ural Branch of the Russian Academy of Sciences: Syktyvkar, Russia, 2014; pp. 218–264. [Google Scholar]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase—mentor of abiotic stress tolerance in crop plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef]
- Martins, N.; Goncalves, S.; Romano, A. Metabolism and aluminum accumulation in Plantago almogravensis and P. algarbiensis in response to low pH and aluminum stress. Biol. Plant 2013, 57, 325–331. [Google Scholar] [CrossRef]
- Shi, Q.H.; Zhu, Z.J.; Li, J.; Qian, Q.Q. Combined effects of excess Mn and low pH on oxidative stress and antioxidant enzymes in cucumber roots. J. Integr. Agri. 2006, 5, 767–772. [Google Scholar]
- Mead, J.F. Free radical mechanisms of lipid damage and consequences for cellular membranes. In Free Radicals in Biology; Pryor, W.A., Ed.; Academic Press: New York, NY, USA, 1976; Volume 1, pp. 51–68. [Google Scholar]
- Xu, C.; Liu, Z.; Zhang, L.; Zhao, C.; Yuan, S.; Zhang, F. Organization of actin cytoskeleton during meiosis I in a wheat thermo-sensitive genic male sterile line. Protoplasma 2013, 250, 415–422. [Google Scholar] [CrossRef]
- Varga, B.; Janda, T.; László, E.; Veisz, O. Influence of abiotic stresses on the antioxidant enzyme activity of cereals. Acta Physiol. Plant 2012, 34, 849–858. [Google Scholar] [CrossRef]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Daur, I.; Nawaz, M.A.; Ahmad, M.Q.; Rana, I.A.; Atif, R.M.; Yang, S.H.; Chung, G. Redox and ionic homeostasis regulations against oxidative, salinity and drought stress in wheat (A Systems Biology Approach). Front. Genet. 2017, 8, 141. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Kushwaha, H.R.; Mustafiz, A.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Analysis of global gene expression profile of rice in response to methylglyoxal indicates its possible role as a stress signal molecule. Front. Plant Sci. 2015, 6, 682. [Google Scholar] [CrossRef]
- Hoque, T.S.; Hossain, M.A.; Mostofa, M.G.; Burritt, D.J.; Fujita, M.; Tran, L.S.P. Methylglyoxal: An emerging signaling molecule in plant abiotic stress responses and tolerance. Front. Plant Sci. 2016, 7, 1341. [Google Scholar] [CrossRef]
- Kaur, C.; Singla-Pareek, S.L.; Sopory, S.K. Glyoxalase and methylglyoxal as biomarkers for plant stress tolerance. Crit. Rev. Plant Sci. 2014, 33, 429–456. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photo peroxidation in isolated chloroplasts: I. kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide induced chilling tolerance in mung beans mediated through ABA-independent glutathione accumulation. Func. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef]
- Arnon, D.T. Copper enzymes in isolated chloroplasts polyphenal oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Doderer, A.; Kokkelink, I.; van der Veen, S.; Valk, B.; Schramk, A.; Douma, A. Purification and characterization of two lipoxygenase isoenzymes from germinating barley. Biochim. Biophys. Acta 1992, 112, 97–104. [Google Scholar] [CrossRef]
- El-Shabrawi, H.; Kumar, B.; Kaul, T.; Reddy, M.K.; Singla-Pareek, S.L.; Sopory, S.K. Redox homeostasis, antioxidant defense, and methylglyoxal detoxification as markers for salt tolerance in Pokkali rice. Protoplasma 2010, 245, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhuyan, M.H.M.B.; Oku, H.; Fujita, M. Exogenous nitric oxide pretreatment protects Brassica napus L. seedlings from paraquat toxicity through the modulation of antioxidant defense and glyoxalase systems. Plant Physiol. Biochem. 2018, 126, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dorr, A.J.M.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Hossain, M.A.; Nakano, Y.; Asada, K. Monodehydroascorbate reductase in spinach chloroplasts and its participation in the regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984, 25, 385–395. [Google Scholar]
- Principato, G.B.; Rosi, G.; Talesa, V.; Giovanni, E.; Uotila, L. Purification and characterization of two forms of glyoxalase II from the liver and brain of Wistar rats. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1987, 911, 349–355. [Google Scholar] [CrossRef]
- Wild, R.; Ooi, L.; Srikanth, V.; Münch, G. A quick, convenient and economical method for the reliable determination of methylglyoxal in millimolar concentrations: The N-acetyl-l-cysteine assay. Anal. Bioanal. Chem. 2012, 403, 2577–2581. [Google Scholar] [CrossRef] [PubMed]
- Addinsoft, XLSTAT V. 2017.1.01: Data Analysis and Statistics Software for Microsoft Excel. Available online: https://www.xlstat.com/en/ (accessed on 11 September 2018).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Root Length (cm) | Shoot FW (g plant−1) | Root FW (g plant−1) | Shoot DW (g plant−1) | Root DW (%) | Leaf RWC (%) | Pro (nmol g−1FW) |
|---|---|---|---|---|---|---|---|---|
| pH 4.0 | 10.32 ± 0.23c | 5.34 ± 0.04c | 0.98 ± 0.003b | 0.17 ± 0.001d | 0.15 ± 0.002b | 0.05 ± 0.001c | 82.31 ± 0.53c | 5.53 ± 0.09b |
| pH 5.5 | 13.17 ± 0.37b | 6.18 ± 0.08b | 1.06 ± 0.056a | 0.21 ± 0.002b | 0.16 ± 0.002a | 0.07 ± 0.002b | 88.67 ± 1.23b | 1.08 ± 0.04c |
| pH 7.0 | 15.33 ± 0.57a | 6.52 ± 0.10a | 1.12 ± 0.014a | 0.24 ± 0.001a | 0.16 ± 0.002a | 0.07 ± 0.001a | 94.31 ± 0.44a | 0.30 ± 0.01d |
| pH 8.5 | 10.52 ± 0.13c | 5.32 ± 0.02c | 0.93 ± 0.003b | 0.18 ± 0.001c | 0.15 ± 0.001b | 0.05 ± 0.001c | 81.12 ± 0.21c | 6.57 ± 0.10a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhuyan, M.H.M.B.; Hasanuzzaman, M.; Mahmud, J.A.; Hossain, M.S.; Bhuiyan, T.F.; Fujita, M. Unraveling Morphophysiological and Biochemical Responses of Triticum aestivum L. to Extreme pH: Coordinated Actions of Antioxidant Defense and Glyoxalase Systems. Plants 2019, 8, 24. https://doi.org/10.3390/plants8010024
Bhuyan MHMB, Hasanuzzaman M, Mahmud JA, Hossain MS, Bhuiyan TF, Fujita M. Unraveling Morphophysiological and Biochemical Responses of Triticum aestivum L. to Extreme pH: Coordinated Actions of Antioxidant Defense and Glyoxalase Systems. Plants. 2019; 8(1):24. https://doi.org/10.3390/plants8010024
Chicago/Turabian StyleBhuyan, M. H. M. Borhannuddin, Mirza Hasanuzzaman, Jubayer Al Mahmud, Md. Shahadat Hossain, Tasnim Farha Bhuiyan, and Masayuki Fujita. 2019. "Unraveling Morphophysiological and Biochemical Responses of Triticum aestivum L. to Extreme pH: Coordinated Actions of Antioxidant Defense and Glyoxalase Systems" Plants 8, no. 1: 24. https://doi.org/10.3390/plants8010024
APA StyleBhuyan, M. H. M. B., Hasanuzzaman, M., Mahmud, J. A., Hossain, M. S., Bhuiyan, T. F., & Fujita, M. (2019). Unraveling Morphophysiological and Biochemical Responses of Triticum aestivum L. to Extreme pH: Coordinated Actions of Antioxidant Defense and Glyoxalase Systems. Plants, 8(1), 24. https://doi.org/10.3390/plants8010024