Genetic, Phenotypic, and Commercial Characterization of an Almond Collection from Sardinia



Abstract
:1. Introduction
2. Results
2.1. Flowering-Related Traits
2.2. Nut- and Kernel-Related Traits
2.3. Kernel and Oil Content Composition
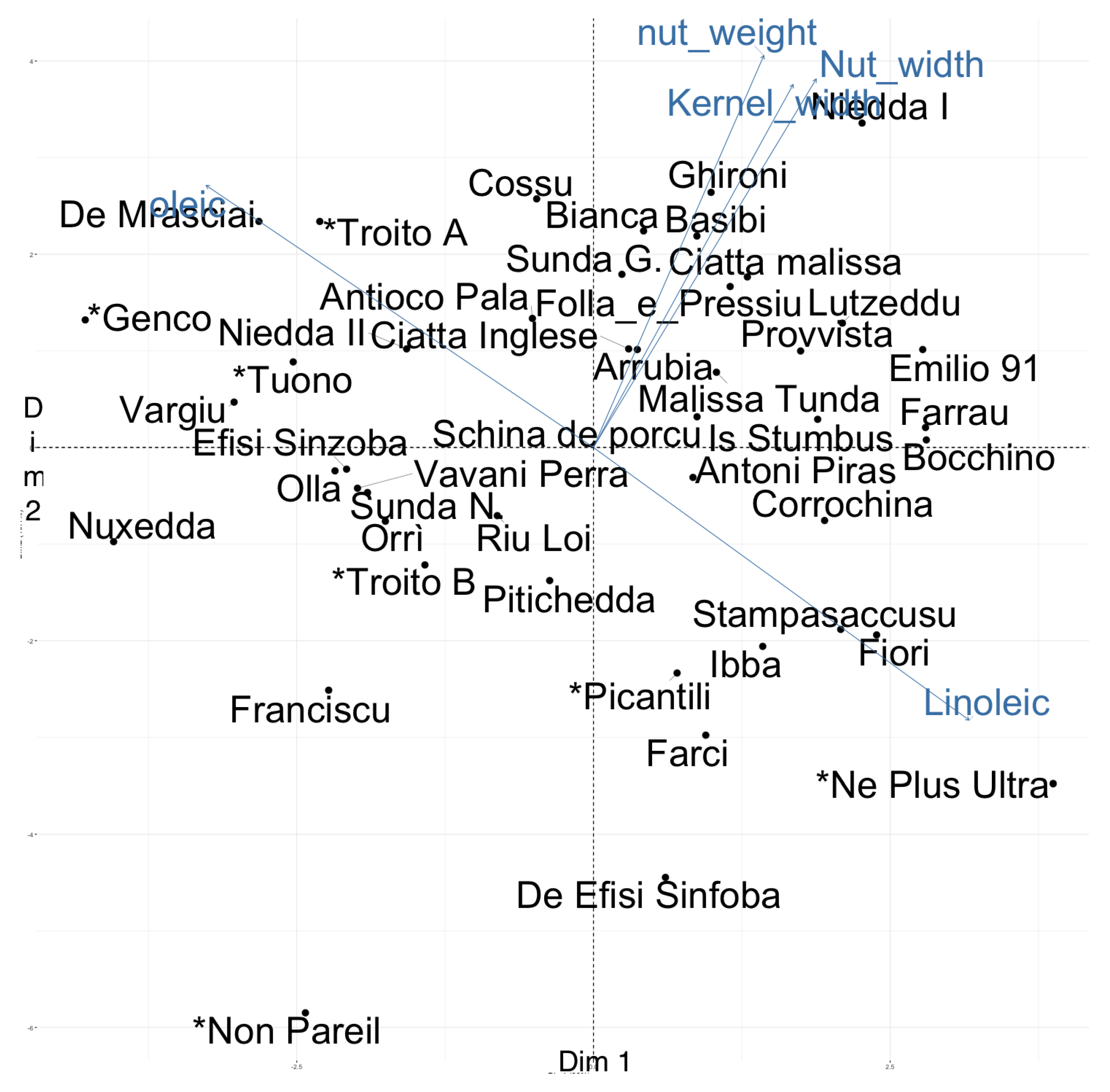
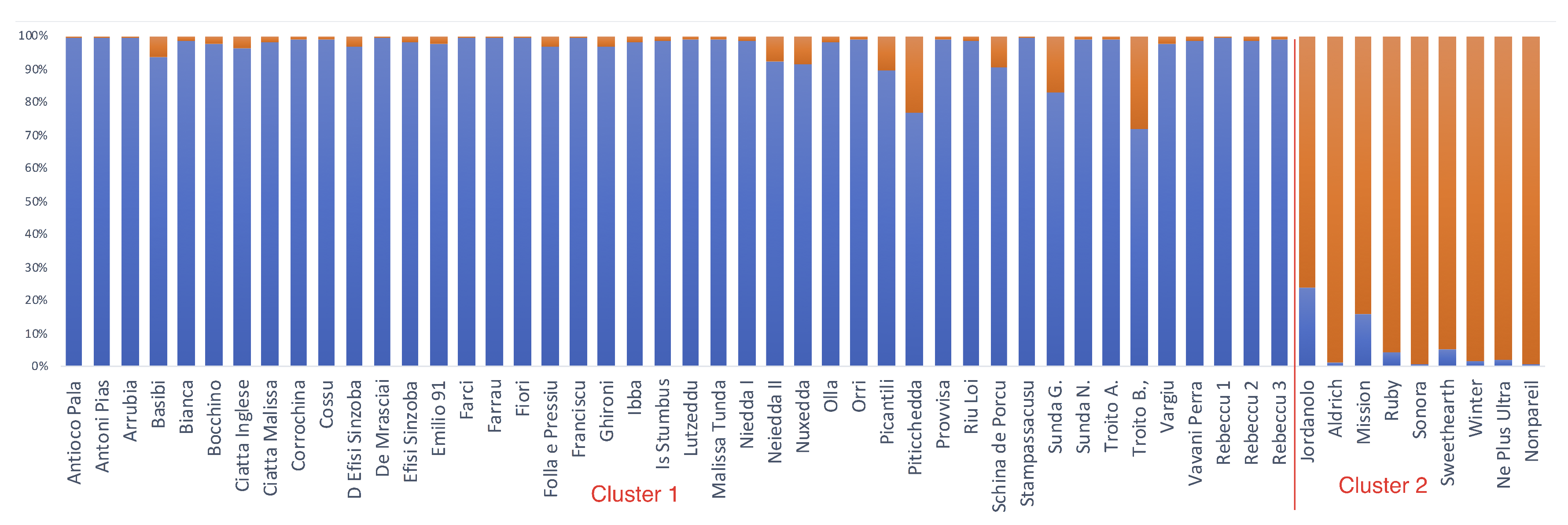
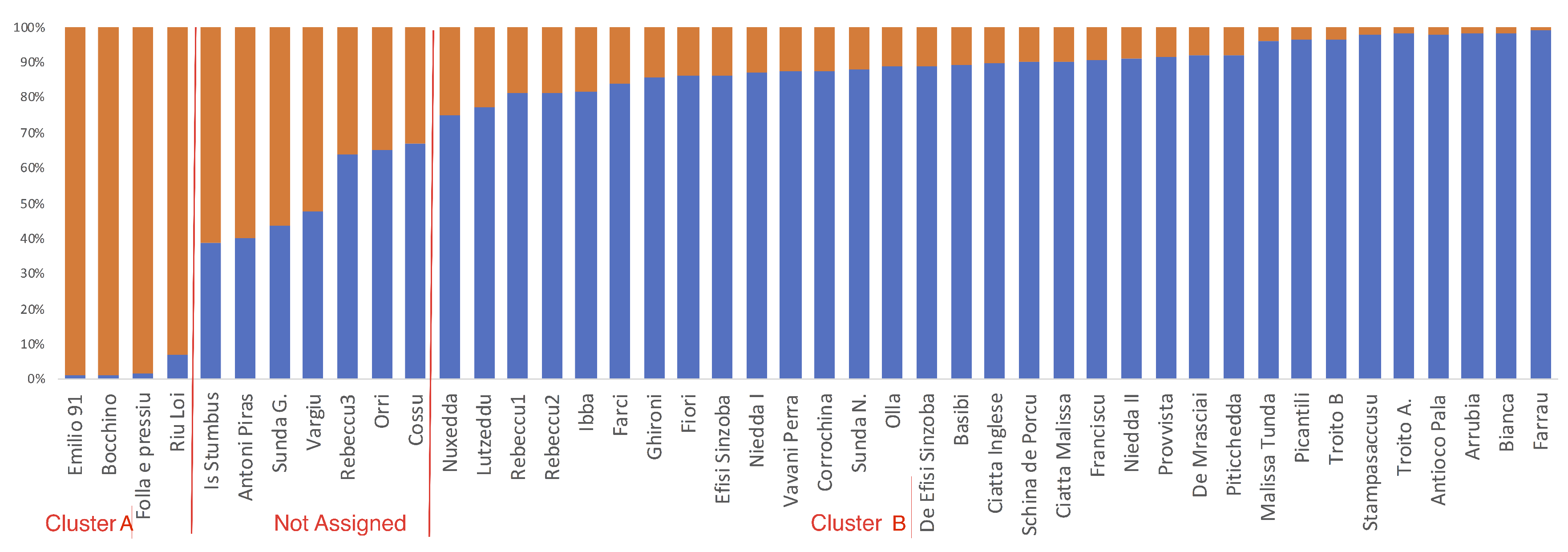
2.4. Genetic and Phenotypic Similarities among Cultivars
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Phenotypic Traits
4.3. Oil Traits
4.4. -Tocopherol Determination
4.5. DNA Extraction and SSR Genotyping
4.6. Genetic and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ladizinsky, G. On the origin of almond. Genet. Resour. Crop Evol. 1999, 46, 143–147. [Google Scholar] [CrossRef]
- Avanzato, D.; Vassallo, I. Following Almond Footprints (Amygdalus Communis L.): Across Sicily Cultivation and Culture, Folk and History, Traditions and Uses; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2006. [Google Scholar]
- Romojaro, F.; Riquelme, F.; Giménez, J.; Llorente, S. Fat content and oil characteristics of some almond varieties. In Fruit Science Reports (Poland); FAO: Roma, Italy, 1988. [Google Scholar]
- Yada, S.; Lapsley, K.; Huang, G. A review of composition studies of cultivated almonds: Macronutrients and micronutrients. J. Food Compos. Anal. 2011, 24, 469–480. [Google Scholar] [CrossRef]
- Kodad, O.; Gracia Gomez, M.; Socias i Company, R. Fatty acid composition as evaluation criterion for kernel quality in almond breeding. In XI Eucarpia Symposium on Fruit Breeding and Genetics 663; ISHS: Leuven, Belgium, 2003; pp. 301–304. [Google Scholar]
- Socias, R.; Kodad, O.; Alonso, J.; Gradziel, T. Almond quality: A breeding perspective. Hortic. Rev. 2008, 34, 197–238. [Google Scholar]
- Ahmad, Z. The uses and properties of almond oil. Complement. Ther. Clin. Pract. 2010, 16, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Kendall, C.W.; Marchie, A.; Parker, T.L.; Connelly, P.W.; Qian, W.; Haight, J.S.; Faulkner, D.; Vidgen, E.; Lapsley, K.G.; et al. Dose response of almonds on coronary heart disease risk factors: Blood lipids, oxidized low-density lipoproteins, lipoprotein (a), homocysteine, and pulmonary nitric oxide: A randomized, controlled, crossover trial. Circulation 2002, 106, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Hyson, D.A.; Schneeman, B.O.; Davis, P.A. Almonds and almond oil have similar effects on plasma lipids and LDL oxidation in healthy men and women. J. Nutr. 2002, 132, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Kester, D.E.; Cunningham, A.K.S. Almonds. In Enciclopedia of Food Science; Janick, J., Moore, J., Eds.; Academic Press: London, UK, 1993; pp. 121–126. [Google Scholar]
- Senesi, E.; Rizzolo, A.; Colombo, C.; Testoni, A. Influence of pre-processing storage conditions on peeled almond quality. In Italian Journal of Food Science: IJFS= Rivista Italiana di Scienza Degli Alimenti; Chiriotti Editori: Pinerolo, Italy, 1996. [Google Scholar]
- Kodad, O.; Socias i Company, R.; Prats, M.; López Ortiz, M. Variability in tocopherol concentrations in almond oil and its use as a selection criterion in almond breeding. J. Hortic. Sci. Biotechnol. 2006, 81, 501–507. [Google Scholar] [CrossRef]
- Zacheo, G.; Cappello, M.; Gallo, A.; Santino, A.; Cappello, A. Changes associated with post-harvest ageing in almond seeds. LWT-Food Sci. Technol. 2000, 33, 415–423. [Google Scholar] [CrossRef]
- Socias i Company, R.; Kodad, O.; Alonso, J.M.; Font-Forcada, C. Fruit Quality in Almond: Chemical Aspects for Breeding Strategies; Zakynthinos, G., Ed.; XIV GREMPA meeting on pistachios and almonds Zaragoza: Zaragoza, Spain, 2011. [Google Scholar]
- López-Ortiz, C.M.; Prats-Moya, S.; Sanahuja, A.B.; Maestre-Pérez, S.E.; Grané-Teruel, N.; Martín-Carratalá, M.L. Comparative study of tocopherol homologue content in four almond oil cultivars during two consecutive years. J. Food Compos. Anal. 2008, 21, 144–151. [Google Scholar] [CrossRef]
- Abdallah, A.; Ahumada, M.H.; Gradziel, T.M. Oil content and fatty acid composition of almond kernels from different genotypes and California production regions. J. Am. Soc. Hortic. Sci. 1998, 123, 1029–1033. [Google Scholar]
- Sathe, S.; Seeram, N.; Kshirsagar, H.; Heber, D.; Lapsley, K. Fatty acid composition of California grown almonds. J. Food Sci. 2008, 73, C607–C614. [Google Scholar] [CrossRef] [PubMed]
- Kodad, O.; Estopañan, G.; Juan, T.; Molino, F.; Mamouni, A.; Messaoudi, Z.; Lahlou, M.; Socias i Company, R. Plasticity and stability in the major fatty acid content of almond kernels grown under two Mediterranean climates. J. Hortic. Sci. Biotechnol. 2010, 85, 381–386. [Google Scholar] [CrossRef]
- Kodad, O.; Estopañán, G.; Juan, T.; Socias i Company, R. Protein content and oil composition of almond from Moroccan seedlings: Genetic diversity, oil quality and geographical origin. J. Am. Oil Chem. Soc. 2013, 90, 243–252. [Google Scholar] [CrossRef]
- Zinelabidine, L. Pomological and biochemical characterization of almond cultivars in Morocco. Türk Tarım ve Doğa Bilimleri 2014, 6, 743–753. [Google Scholar]
- Ledbetter, C.A.; Sisterson, M.S. Distinguishing Nonpareil Marketing Group Almond Cultivars through Multivariate Analyses. J. Food Sci. 2013, 78, S1430–S1436. [Google Scholar] [CrossRef] [PubMed]
- Sottile, F.; Barone, E.; Barbera, G.; Palasciano, M. The Italian almond industry: New perspectives and ancient tradition. In VI International Symposium on Almonds and Pistachios 1028; ISHS: Leuven, Belgium, 2013; pp. 401–407. [Google Scholar]
- Stylianides, D. La culture de l’amandier in Grece. Options Mediterr; CiHEAM: Paris, France, 1976; pp. 72–74. [Google Scholar]
- Altuntas, E.; Gercekcioglu, R.; Kaya, C. Selected mechanical and geometric properties of different almond cultivars. Int. J. Food Prop. 2010, 13, 282–293. [Google Scholar] [CrossRef]
- Rigoldi, M.P.; Rapposelli, E.; De Giorgio, D.; Resta, P.; Porceddu, A. Genetic diversity in two Italian almond collections. Electron. J. Biotechnol. 2015, 18, 40–45. [Google Scholar] [CrossRef]
- Kodad, O.; Alonso, J.M.; Espiau, M.T.; Estopañán, G.; Juan, T. Chemometric characterization of almond germplasm: Compositional aspects involved in quality and breeding. J. Am. Soc. Hortic. Sci. 2011, 136, 273–281. [Google Scholar]
- Nassar, A.R.; El-Tahawi, B.S.; El-Deen, S.A.S. Chromatographic identification of oil and amino acid constitutents in kernels of some almond varieties. J. Am. Oil Chem. Soc. 1977, 54, 553–556. [Google Scholar] [CrossRef]
- Nanos, G.D.; Kazantzis, I.; Kefalas, P.; Petrakis, C.; Stavroulakis, G.G. Irrigation and harvest time affect almond kernel quality and composition. Sci. Hortic. 2002, 96, 249–256. [Google Scholar] [CrossRef]
- Kumar, K.; Ahuja, K.; Uppal, D. Kernel quality of almonds (Prunus amygdalus Batsch) in terms of oil content, fatty acid composition and phospholipid content. J. Food Sci. Technol. (India) 1994, 31, 335–337. [Google Scholar]
- Kumar, K.; Sharma, S.; Goyal, R. Enzymatic changes and oil accumulation during almond kernel development. In XXV International Horticultural Congress, Part 5: Culture Techniques with Special Emphasis on Environmental Implications 515; ISHS: Leuven, Belgium, 1998; pp. 287–296. [Google Scholar]
- Mehran, M.; Filsoof, M. Characteristics of Iranian almond nuts and oils. J. Am. Oil Chem. Soc. 1974, 51, 433–434. [Google Scholar] [CrossRef]
- Barbera, G.; La Mantia, T.; Monastra, F.; De Palma, L.; Schirra, M. Response of Ferragnes and Tuono almond cultivars to different environmental conditions in southern Italy. In I International Congress on Almond 373; ISHS: Leuven, Belgium, 1993; pp. 125–128. [Google Scholar]
- Ruggeri, S.; Cappelloni, M.; Gambelli, L.; Nicoli, S.; Carnovale, E. Chemical composition and nutritive value of nuts grown in Italy. Available online: http://agris.fao.org/agris-search/search.do?recordID=IT2000060508 (accessed on 13 October 2018).
- Askin, M.; Balta, M.; Tekintas, F.; Kazankaya, A.; Balta, F. Fatty acid composition affected by kernel weight in almond [Prunus dulcis (Mill.) DA Webb.] genetic resources. J. Food Compos. Anal. 2007, 20, 7–12. [Google Scholar] [CrossRef]
- Aslantas, R.; Guleryuz, M.; Turan, M. Some chemical contents of selected almond (Prunus amygdalus Batsch) types. Cah. Options Mediterraneennes 2001, 56, 347–350. [Google Scholar]
- Kaska, N.; Kafkas, S.; Padulosi, S.; Wassimi, N.; Ak, B. Characterization of nut species of Afghanistan: I-Almond. In IV International Symposium on Pistachios and Almonds 726; ISHS: Leuven, Belgium, 2005; pp. 147–156. [Google Scholar]
- Kodad, O.; Alonso, J.M. Genotypic and Environmental Effects on Tocopherol Content in Almond. Antioxidants 2018, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Rizzolo, A.; Baldo, C.; Polesello, A. Application of high-performance liquid chromatography to the analysis of niacin and biotin in Italian almond cultivars. J. Chromatogr. A 1991, 553, 187–192. [Google Scholar] [CrossRef]
- Yada, S.; Huang, G.; Lapsley, K. Natural variability in the nutrient composition of California-grown almonds. J. Food Compos. Anal. 2013, 30, 80–85. [Google Scholar] [CrossRef]
- Kodad, O.; Estopañán, G.; Juan, T. Tocopherol concentration in almond oil from Moroccan seedlings: Geographical origin and post-harvest implications. J. Food Compos. Anal. 2014, 33, 161–165. [Google Scholar] [CrossRef]
- Maestri, D.; Martínez, M.; Bodoira, R.; Rossi, Y.; Oviedo, A.; Pierantozzi, P.; Torres, M. Variability in almond oil chemical traits from traditional cultivars and native genetic resources from Argentina. Food Chem. 2015, 170, 55–61. [Google Scholar] [CrossRef] [PubMed]
- I Martí, A.F.; i Forcada, C.F.; Kamali, K.; Rubio-Cabetas, M.J.; Wirthensohn, M. Molecular analyses of evolution and population structure in a worldwide almond [Prunus dulcis (Mill.) DA Webb syn. P. amygdalus Batsch] pool assessed by microsatellite markers. Genet. Resour. Crop Evol. 2015, 62, 205–219. [Google Scholar] [CrossRef]
- Segura, J.M.A.; Socias i Company, R.; Kodad, O. Late-blooming in almond: A controversial objective. Sci. Hortic. 2017, 224, 61–67. [Google Scholar] [CrossRef]
- Prudencio, A.; Martínez-Gómez, P.; Dicenta, F. Evaluation of breaking dormancy, flowering and productivity of extra-late and ultra-late flowering almond cultivars during cold and warm seasons in South-East of Spain. Sci. Hortic. 2018, 235, 39–46. [Google Scholar] [CrossRef]
- Gülcan, R. Descriptor List for Almond (Prunus Amygdalus)(Revised); International Board for Plant Genetic Resources (IBPGR): Roma, Italy, 1985. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Origin | Initial Flowering | Maximum Flowering | Final Flowering |
|---|---|---|---|---|
| Antioco Pala | Sardinia | |||
| Antoni Piras | Sardinia | |||
| Arrubia | Sardinia | |||
| Basibi | Sardinia | |||
| Bianca | Sardinia | |||
| Bocchino | Sardinia | |||
| Ciatta Inglese | Sardinia | 46.00 | ||
| Ciatta Malissa | Sardinia | 45.67 | ||
| Corrochina | Sardinia | |||
| Cossu | Sardinia | |||
| De Efisi Sinzoba | Sardinia | |||
| De Mrasciai | Sardinia | |||
| Efisi Sinzoba | Sardinia | |||
| Emilio 91 | Sardinia | |||
| Farci | Sardinia | |||
| Farrau | Sardinia | |||
| Fiori | Sardinia | |||
| Folla e pressiu | Sardinia | 33.33 | ||
| Franciscu | Sardinia | |||
| * Genco | Apulia | |||
| Ghironi | Sardinia | |||
| Ibba | Sardinia | |||
| Is Stumbus | Sardinia | |||
| Lutzeddu | Sardinia | |||
| Malissa Tunda | Sardinia | |||
| * Ne Plus Ultra | USA | |||
| Niedda I | Sardinia | |||
| Niedda II | Sardinia | |||
| * Nonpareil | USA | |||
| Nuxedda | Sardinia | |||
| Olla | Sardinia | |||
| Orri | Sardinia | 37.67 | 42.33 | |
| * Picantili | Russia | |||
| Pitichedda | Sardinia | |||
| Provvista | Sardinia | |||
| Riu Loi | Sardinia | |||
| Schina de porcu | Sardinia | |||
| Stampasaccusu | Sardinia | |||
| Sunda G. | Sardinia | |||
| Sunda N. | Sardinia | 61.33 | ||
| * Troito A | Italy−Greece | |||
| * Troito B | Italy−Greece | |||
| * Tuono | Sardinia | |||
| Vargiu | Sardinia | |||
| Vavani Perra | Sardinia |
| Initial Flowering | Factor | DF | SS | F | Max Flowering | Factor | DF | SS | F |
| days after 1 Jan | Cultivar | 44 | 10,543.08 | days after 1 Jan | Cultivar | 44 | 8943.06 | ||
| Year | 2 | 759.53 | Year | 2 | 96.13 | ||||
| Error | 44 | 1191.81 | Error | 88 | 899.20 | ||||
| Final Flowering | Factor | DF | SS | F | Nut Weigth (gr) | Factor | DF | SS | F |
| days after 1 jan | Cultivar | 44 | 5031.08 | Cultivar | 44 | 286.06 | |||
| Year | 2 | 477.57 | Year | 2 | 1.04 | ||||
| Error | 88 | 3426.42 | Error | 88 | 70.38 | ||||
| Kernel Weight (g) | Factor | DF | SS | F | Kernel Shelling % | Factor | DF | SS | F |
| Cultivar | 44 | 7.56 | Cultivar | 44 | 20,837.27 | ||||
| Year | 2 | 0.77 | Year | 2 | 122.29 | ||||
| Error | 88 | 2.52 | Error | 88 | 509.31 | ||||
| Kernel Yield (kg/plant) | Factor | DF | SS | F | Double Kernels % | Factor | DF | SS | F |
| Cultivar | 44 | 72,155.65 | Cultivar | 44 | 20,777.78 | ||||
| Year | 2 | 30.706 | Year | 2 | 834.311 | ||||
| Error | 88 | 11,419.6 | Error | 88 | 4729.69 | ||||
| Failed Kernels % | Factor | DF | SS | F | Nut Length (cm) | Factor | DF | SS | F |
| Cultivar | 44 | 557.34 | Cultivar | 44 | 13.30 | ||||
| Year | 2 | 13.38 | Year | 2 | 0.12 | ||||
| Error | 88 | 240.62 | Error | 88 | 5.92 | ||||
| Nut Width (cm) | Factor | DF | SS | F | Kernel Length (cm) | Factor | DF | SS | F |
| Cultivar | 44 | 10.23 | Cultivar | 44 | 6.84 | ||||
| Year | 2 | 0.106 | Year | 2 | 0.78 | ||||
| Error | 88 | 7.32 | Error | 88 | 1.47 | ||||
| Kernel Width (cm) | Factor | DF | SS | F | |||||
| Cultivar | 44 | 2.59 | |||||||
| Year | 2 | 0.03 | |||||||
| Error | 88 | 0.74 |
| Cultivar Name | Nut Weight (g) | Kernel Weight (g) | Kernel Yield (kg/plant) | Nut Length (cm) | Nut Width (cm) | Kernel Length (cm) | Kernel Width (cm) | Kernel Failed (%) | Kernel Double (%) |
|---|---|---|---|---|---|---|---|---|---|
| Antioco Pala | |||||||||
| Antoni Piras | |||||||||
| Arrubia | |||||||||
| Basibi | |||||||||
| Bianca | |||||||||
| Bocchino | |||||||||
| Ciatta Inglese | |||||||||
| Ciatta Malissa | |||||||||
| Corrochina | |||||||||
| Cossu | |||||||||
| De Efisi Sinzoba | |||||||||
| De Mrasciai | |||||||||
| Efisi Sinzoba | |||||||||
| Emilio 91 | |||||||||
| Farci | |||||||||
| Farrau | |||||||||
| Fiori | |||||||||
| Folla e pressiu | |||||||||
| Franciscu | |||||||||
| * Genco | |||||||||
| Ghironi | |||||||||
| Ibba | |||||||||
| Is Stumbus | |||||||||
| Lutzeddu | |||||||||
| Malissa Tunda | |||||||||
| * Ne Plus Ultra | |||||||||
| Niedda I | |||||||||
| Niedda II | |||||||||
| * Nonpareil | |||||||||
| Nuxedda | |||||||||
| Olla | |||||||||
| Orri | |||||||||
| * Picantili | |||||||||
| Pitichedda | |||||||||
| Provvista | |||||||||
| Riu Loi | |||||||||
| Schina de porcu | |||||||||
| Stampasaccusu | |||||||||
| Sunda G. | |||||||||
| Sunda N. | |||||||||
| * Troito A | |||||||||
| * Troito B | |||||||||
| * Tuono | |||||||||
| Vargiu | |||||||||
| Vavani Perra |
| Trait | Source | DF | SS | F | Trait | Source | DF | SS | F-Value |
| Total oil | Cultivar | 44 | 3351.96 | 24.8634 | Palmitic acid | Cultivar | 44 | 72.32 | 156.01 |
| Year | 2 | 139.73 | 6.1521 | Year | 2 | 6.39 | 304.69 | ||
| C × Y | 88 | 1658.78 | 6.1521 | C × Y | 88 | 11.79 | 12.77 | ||
| Error | 270 | 827.27 | Error | 270 | 2.83 | ||||
| Trait | Source | DF | SS | F | Trait | Source | DF | SS | F |
| Palmitoleic acid | Cultivar | 44 | 2.61 | 76.74 | Stearic acid | Cultivar | 44 | 25.18 | 96.92 |
| Year | 2 | 1.67 | 1077.08 | Year | 2 | 2.87 | 243.47 | ||
| C × Y | 88 | 8.76 | 12.77 | C × Y | 88 | 6.44 | 12.40 | ||
| Error | 270 | 0.20 | Error | 270 | 1.59 | ||||
| Trait | Source | DF | SS | F | Trait | Source | DF | SS | F |
| Oleic acid | Cultivar | 44 | 3669.07 | 241.95 | Linoleic acid | Cultivar | 44 | 2928.90 | 244.40 |
| Year | 2 | 262.96 | 381.51 | Year | 2 | 295.56 | 542.59 | ||
| C × Y | 88 | 1078.70 | 35.57 | C × Y | 88 | 877.18 | 36.60 | ||
| Error | 270 | 93.05 | Error | 270 | 73.53 | ||||
| Trait | Source | DF | SS | F | Trait | Source | DF | SS | F |
| Linolenic acid | Cultivar | 44 | 1534,850.9 | 126.1863 | Tocopherol acid | Cultivar | 44 | 0.008 | 22.32 |
| Year | 2 | 870,549.8 | 1574.571 | Year | 2 | 0.015 | 849.06 | ||
| C × Y | 88 | 363,388.7 | 14.9378 | C × Y | 88 | 0.013 | 17.22 | ||
| Error | 270 | 8.63e-6 | Error | 270 | 74.36 |
| Cultivar Name | Oil (% on d.w.) | Palmitic (% of Oil) | Palmitoleic (% of Oil) | Stearic (% of Oil) | Oleic (% of Oil) | Linoleic (% of Oil) | Tocopherol (mg/kg of Oil) |
|---|---|---|---|---|---|---|---|
| Antioco Pala | |||||||
| Antoni Piras | |||||||
| Arrubia | |||||||
| Basibi | |||||||
| Bianca | |||||||
| Bocchino | |||||||
| Ciatta Inglese | |||||||
| Ciatta Malissa | |||||||
| Corrochina | |||||||
| Cossu | |||||||
| De Efisi Sinzoba | |||||||
| De Mrasciai | |||||||
| Efisi Sinzoba | |||||||
| Emilio91 | |||||||
| Farci | |||||||
| Farrau | |||||||
| Fiori | |||||||
| Foll’e pressiu | |||||||
| Franciscu | |||||||
| * Genco | |||||||
| Ghironi | |||||||
| Ibba | |||||||
| Is Stumbus | |||||||
| Lutzeddu | |||||||
| Malissa Tunda | |||||||
| * Ne Plus Ultra | |||||||
| Niedda I | |||||||
| Niedda II | |||||||
| * Non Pareil | |||||||
| Nuxedda | |||||||
| Olla | |||||||
| Orri | |||||||
| * Picantili | |||||||
| Pitichedda | |||||||
| Provvista | 5.68 | ||||||
| Riu Loi | |||||||
| Schina de Porcu | |||||||
| Stampasaccusu | |||||||
| Sunda G. | |||||||
| Sunda N. | |||||||
| * TroitoA | |||||||
| * TroitoB | |||||||
| * Tuono | |||||||
| Vargiu | 5.56 | ||||||
| Vavani Perra |
| Trait | Variable | CL-A | CL-B |
|---|---|---|---|
| Flowering | Start (days after 1 Jan) | ||
| Maximum (days after the 1 Jan) | |||
| Final (days after the 1 Jan) | |||
| Kernel | Width (cm) | ||
| Oil composition | Stearic (% of oil) | ||
| Tocopherol (mg/kg) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapposelli, E.; Rigoldi, M.P.; Satta, D.; Delpiano, D.; Secci, S.; Porceddu, A. Genetic, Phenotypic, and Commercial Characterization of an Almond Collection from Sardinia. Plants 2018, 7, 86. https://doi.org/10.3390/plants7040086
Rapposelli E, Rigoldi MP, Satta D, Delpiano D, Secci S, Porceddu A. Genetic, Phenotypic, and Commercial Characterization of an Almond Collection from Sardinia. Plants. 2018; 7(4):86. https://doi.org/10.3390/plants7040086
Chicago/Turabian StyleRapposelli, Emma, Maria Pia Rigoldi, Daniela Satta, Donatella Delpiano, Sara Secci, and Andrea Porceddu. 2018. "Genetic, Phenotypic, and Commercial Characterization of an Almond Collection from Sardinia" Plants 7, no. 4: 86. https://doi.org/10.3390/plants7040086
APA StyleRapposelli, E., Rigoldi, M. P., Satta, D., Delpiano, D., Secci, S., & Porceddu, A. (2018). Genetic, Phenotypic, and Commercial Characterization of an Almond Collection from Sardinia. Plants, 7(4), 86. https://doi.org/10.3390/plants7040086