Cinnamic Acid Inhibited Growth of Faba Bean and Promoted the Incidence of Fusarium Wilt

Abstract
1. Introduction
2. Results
2.1. Effect of Cinnamic Acid on Plant Growth
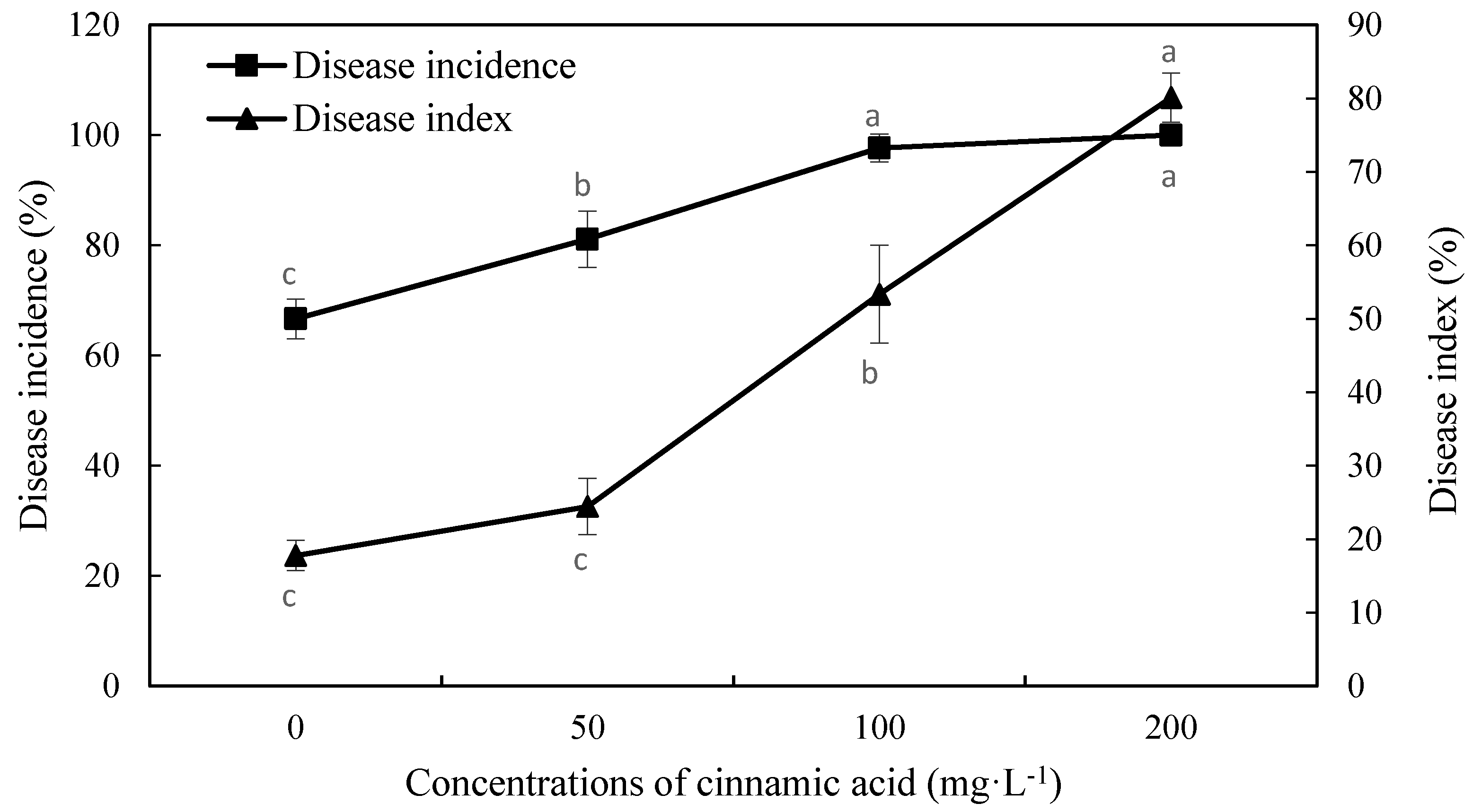
2.2. Effect of Cinnamic Acid on Fusarium Wilt Incidence and Disease Indices
2.3. Effect of Cinnamic Acid on Seedling Root Physiological Activity
2.4. Effect of Cinnamic Acid on Pathogenesis-Related Faba Bean Seedling Root Proteins
2.5. Effect of Cinnamic Acid on FOF Mycotoxin Production
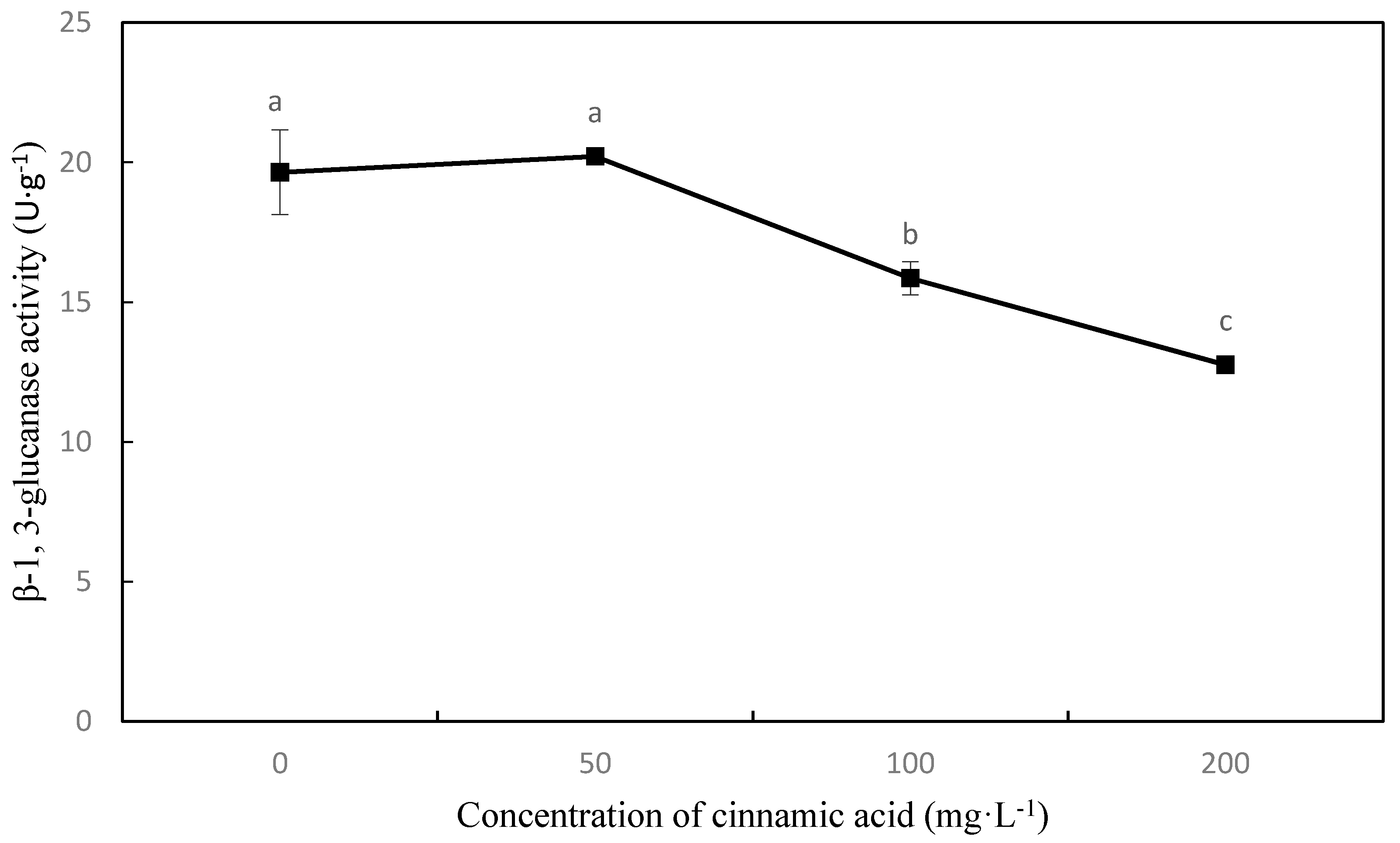
2.6. Effect of Cinnamic Acid on the Activities of Hydrolytic Enzymes Related to Pathogenesis
3. Discussion
4. Materials and Methods
4.1. Test Materials
4.2. Preparation of Spore Suspension
4.3. Greenhouse Experiments
4.4. Determination of Faba Bean Growth Indices
4.5. Measurement of the Incidence of Fusarium Wilt
4.6. Determination of Antioxidant Enzymes and Membrane Lipid Peroxidation
4.7. Determination of Pathogenesis-Related Protein activities
4.8. Extraction and Quantification of Mycotoxin
4.9. Measurement of Pathogenesis-Related Hydrolytic Enzyme Activities
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stoddard, F.L.; Nicholas, A.; Rubiales, D.; Thomas, J.; Villegas-Fernández, A.M. Integrated pest, disease and weed management in faba bean. Field Crops Res. 2010, 115, 308–318. [Google Scholar] [CrossRef]
- Jensen, E.S.; Peoples, M.; Hauggaard-Nielsen, H. Faba bean in cropping systems. Field Crops Res. 2010, 115, 203–216. [Google Scholar] [CrossRef]
- Zhen, W.C. Study on the Mechanism and Control of Replant Disease on Strawberry (Fragaria ananassa Duch). Master’s Thesis, Agricultural University of Hebei Province, Baoding, China, 2003. [Google Scholar]
- Wu, X.H.; Lu, Z.J.; Wang, P.P.; Yang, L.Y. Current advances in integrated management of watermelon Fusarium wilt. Plant Prot. 2011, 37, 27–32. [Google Scholar]
- Zhang, F.G.; Zhu, Z.; Yang, X.M.; Ran, W.; Shen, Q.R. Trichoderma harzianum T–E5 significantly affects cucumber root exudates and fungal community in the cucumber rhizosphere. Appl. Soil Ecol. 2013, 72, 41–48. [Google Scholar] [CrossRef]
- Ye, Y. Chinese Faba Beans, 1st ed.; China Agriculture Press: Beijing, China, 2003; pp. 450–451. ISBN 7-109-08243-1. [Google Scholar]
- Zhou, X.G.; Wu, F.Z. P-Coumaric Acid Influenced cucumber rhizosphere Soil microbial communities and the growth of Fusarium oxysporum f. sp. cucumerinum owen. PLoS ONE 2012, 7, e48288. [Google Scholar] [CrossRef]
- Chen, S.L.; Zhou, B.L.; Lin, S.S.; Xia, L.; Ye, X.L. Accumulation of cinnamic acid and vanillin in eggplant root exudates and the relationship with continuous cropping obstacle. Afr. J. Biotechnol. 2011, 10, 2659–2665. [Google Scholar]
- Hao, W.Y.; Ran, W.; Shen, Q.R.; Ren, L.X. Effects of root exudates from watermelon, rice plants and phenolic acids on Fusarium oxysporum f. sp. niveum. Sci. Agric. Sin. 2010, 43, 2443–2452. [Google Scholar]
- Li, Z.F.; Yang, Y.Q.; Xie, D.F.; Zhu, L.F.; Zhang, Z.G.; Lin, W.X. Identification of autotoxic compounds in fibrous roots of rehmannia (RehmannHia glutinosa Libosch.). PLoS ONE 2012, 7, e28806. [Google Scholar] [CrossRef]
- Wu, Z.J.; Yang, L.; Wang, R.Y.; Zhang, Y.B.; Shang, Q.H.; Wang, L.; Ren, Q.; Xie, Z.K. In vitro study of the growth, development and pathogenicity responses of Fusarium oxysporum to phthalic acid, an autotoxin from Lanzhou lily. World J. Microbiol. Biotechnol. 2015, 31, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.F. Research on Promotive Effects of Fusarium Wilt in Cucumis sativus by Cinnamic Acid, an Autotoxin in Root Exudates of Cucumis saitvus L., and Mitigation Mechanism by Grafting and Cinnamic Acid—Degrading Microbial Strains. Master’s Thesis, Zhejiang University, Hangzhou, China, 2004. [Google Scholar]
- Asaduzzaman, M.; Asao, T. Autotoxicity in beans and their allelochemicals. Sci. Hortic. 2012, 134, 26–31. [Google Scholar] [CrossRef]
- Hu, Y.S.; Wu, K.; Li, C.X.; Sun, F.L.; Jia, X.C. Effects of phenolic compounds on the growth of cucumis sativus seedlings and Fusarium oxysporum hypha. Chin. J. Ecol. 2007, 26, 1738–1742. [Google Scholar]
- Wang, R.H.; Zhou, B.L.; Zhang, Q.F.; Lian, H.; Fu, Y.W. Effects of vanillin and cinnamic acid in root exudates of eggplants on Verticillium dahliae etc. Acta Ecol. Sin. 2006, 26, 3152–3155. [Google Scholar]
- Yang, J.X.; Gao, W.W. Effects of Phenolic Allelochemicals on the pathogen of Panax quiquefolium L. Chin. Agric. Sci. Bull. 2009, 25, 207–211. [Google Scholar]
- Dong, Y.; Dong, K.; Yang, Z.X.; Zheng, Y.; Tang, L. Microbial and physiological mechanisms for alleviating Fusarium wilt of faba bean in intercropping system. Chin. J. Appl. Ecol. 2016, 27, 1010–1020. [Google Scholar]
- Song, L.; Pan, K.W.; Wang, J.C.; Ma, Y.H. Effects of phenolic acids on seed germination and seedling antioxidant enzyme activity of alfalfa. Acta Ecol. Sin. 2006, 26, 3393–3403. [Google Scholar]
- Li, X.; Zhou, B.L.; Chen, S.L.; Lin, S.S. Effects of the auto toxic substance in eggplant on pepper seed germination and Fusarium oxysporum f sp. Acta Ecol. Sin. 2009, 29, 960–965. [Google Scholar]
- Tian, G.L.; Bi, Y.M.; Sun, Z.J.; Zhang, L.S. Phenolic acids in the plow layer soil of strawberry fields and their effects on the occurence of strawberry anthracnose. Eur. J. Plant Pathol. 2015, 143, 581–594. [Google Scholar] [CrossRef]
- Qi, Y.Z.; Su, Y.; Wang, N.; Zhen, W.C. Observation on histological structure of strawberry roots after inoculating Fusarium oxysporum f. sp. fragariae under 4-hydroxybenzoic acid stress. Acta Hortic. Sin. 2015, 42, 1909–1918. [Google Scholar]
- Wang, Q.; Li, X.L. Effects of benzoic and cinnamic acids on watermelon seedling growth and fusarium wilt occurrence. J. China Agric. Univ. 2003, 8, 83–86. [Google Scholar] [CrossRef]
- Guo, H.W.; Guo, S.R.; Liu, L.; Sun, J.; Huang, B.J. Effects of continuous cropping on physical and chemical properties of soil, physiological resistance and ion absorption of pepper. Soils 2012, 44, 1041–1047. [Google Scholar]
- Wang, R.H.; Qu, G.F.; Zhang, Q.F.; Li, D.D.; Liu, F.; Qin, Z.H. Effect of vanillin on seedling growth and physiological characteristics of tomato. North. Hortic. 2014, 5, 12–14. [Google Scholar]
- Wu, F.Z. Studies on Physiological and Biochemical Mechanism of Exogenous Phenolic Acid on Cucumber’s Auto-Toxic Effect. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2002. [Google Scholar]
- Qi, Y.Z. Study on Synergistic Effect of Root Allelochemical and Pathogen in the Replant Disease of Strawberry. Master’s Thesis, Agricultural University of Hebei Province, Baoding, China, 2008. [Google Scholar]
- Wang, Y.F.; Pan, F.B.; Zhan, X.; Wang, G.S.; Zhang, G.D.; Hu, Y.L.; Chen, X.S.; Mao, Z.Q. Effects of five kinds of phenolic acid on the function of mitochondria and antioxidant systems in roots of Malus hupehensis Rehd seedlings. Acta Ecol. Sin. 2015, 35, 6566–6573. [Google Scholar]
- Jiao, X.L.; Bi, X.B.; Gao, W.W. Allelopathic effect of p-coumaric acid on American ginseng and its physiological mechanism. Acta Ecol. Sin. 2015, 35, 3006–3013. [Google Scholar]
- Xu, W.H.; Liu, D.; Wu, F.Z.; Liu, S.W. Root exudates of wheat are involved in suppression of Fusarium wilt in watermelon in watermelon-wheat companion cropping. Eur. J. Plant Pathol. 2015, 141, 209–216. [Google Scholar] [CrossRef]
- Zuo, Y.H.; Kang, Z.S.; Yang, C.P.; Rui, H.Y.; Lou, S.B.; Liu, X.R. Relationship between activities of 13-1.3-glacanase and chitinase and resistance to phytophthora root rot in soybean. Acta Phytopathol. Sin. 2009, 39, 600–607. [Google Scholar]
- Kang, Y.Y.; Zhou, X.M.; Yang, X.; Lin, J. Effect of p-hydroxybenzoic acid on induced resistance of flowering Chinese cabbage to anthracnose and the associated plant physiological characters. Acta Phytopathol. Sin. 2014, 44, 393–404. [Google Scholar]
- Ren, L.X. Physiological Mechanisms for Suppressing Watermelon Wilt Disease by Intercropped with Aerobic Rice. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2012. [Google Scholar]
- Ye, S.F.; Yu, J.Q.; Peng, Y.H.; Zheng, J.H.; Zou, L.Y. Incidence of fusarium wilt in Cucumis sativus L. is promoted by cinnamic acid, an autotoxin root exudate. Plant Soil 2004, 263, 143–150. [Google Scholar] [CrossRef]
- Wu, H.S.; Liu, D.Y.; Ling, N.; Bao, W.; Ying, R.R.; Shen, Q.R. Influence of root exudates of watermelon on Fusarium oxysporum f. sp. niveum. Soil Biol. Biochem. 2009, 73, 1150–1156. [Google Scholar]
- Zhen, W.C.; Wang, X.Y.; Kong, J.Y.; Cao, K.Q. Determination of phenolic acids in root exudates and decomposing products of strawberry and their allelopathy. J. Agric. Univ. Hebei 2004, 27, 74–78. [Google Scholar]
- Chen, S.L.; Zhou, B.L.; Wang, R.H.; Fu, Y.W. Regulation effect of grafting on cinnamic acid and vanillin in eggplant root exudates. Chin. J. Appl. Ecol. 2008, 19, 2394–2399. [Google Scholar]
- Hao, W.Y.; Ren, L.X.; Ran, W.; Shen, Q.R. Allelopathic effects of root exudates from watermelon and rice plants on Fusarium oxysporum f. sp. niveum. Plant Soil 2010, 336, 485–497. [Google Scholar] [CrossRef]
- Qi, Y.Z.; Jin, J.J.; Chang, N.; Zhang, X.J.; Yin, B.Z.; Zhen, W.C. Improvement effect of 4-hydroxybenzoic acid on occurrence of wilt disease of strawberry. China Plant Prot. 2016, 9, 5–10. [Google Scholar]
- Johnson, R.D.; Johnson, L.; Itoh, Y.; Kodama, M.; Otani, H.; Kohmoto, K. Cloning and characterization of a cyclic peptide synthetase gene from Alternaria alternata apple pathotype whose product is involved in AM-toxin synthesis and pathogenicity. Mol. Plant-Microbe Interact. 2000, 13, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Masumake, A.; Tamaka, A.; Tsuge, T.; Peever, T.L.; Timmer, L.W.; Yamamoto, M.; Yamamoto, H.; Akimitsu, K. Distribution and characterization of AKT homologs in the tangerine pathotype of Alternaria alternata. Phyotopathology 2000, 90, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Masumake, A.; Ohtani, K.; Peever, T.L.; Timmer, L.W.; Tsuge, T.; Yamamoto, M.; Yamamoto, H.; Akimitsu, K. An Isolate of Alternaria alternate that is pathogenic to both tangerines and rough lemon and produces two host-selective Toxins, ACT-and ACR-toxins. Phyotopathology 2005, 95, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Geimba, M.P.; Corbellini, V.A.; Scroferneker, M.L. Chemical and immunological differentiation of exoantigens from four Bipolaris sorokiniara strains. Process Biochem. 2005, 40, 2051–2057. [Google Scholar]
- Kosiak, E.B.; Jensen, A.H.; Rundberget, T.; Torp, M.T.G.M. Morphological, chemical and molecular differentiation of Fusarium equiseti isolated from Norwegian cereals. Int. J. Food Microbiol. 2005, 99, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Ghédira-Chékir, L.; Maaroufi, K.; Zakhama, A.; Ellouz, F.; Dhouib, S.; Creppy, E.E.; Bacha, H. Induction of a SOS repaire system in lysogenic bacteria by zearalenone and its derivatives by vitamin E. Chem.-Biol. Interact. 1998, 113, 15–25. [Google Scholar] [CrossRef]
- Minervini, F.; Giannoccaro, A.; Cavallini, A.; Visconti, A. Investigations on cellular proliferation induced by zearalenone and its derivatives in relation to the estrogenic parameters. Toxicol. Lett. 2005, 159, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Wang, K.Y.; Dong, G.Q.; Yuan, K.P.; Lin, K.J. The correlations in the pathogenicity to the enzymes’ activities of Thanatephorus Cucumeris causing rice sheath blight disease. Acta Agric. Zhejiangensis 1992, 4, 8–14. [Google Scholar]
- Vidhyasekaran, P. Fungal Pathogenesis in Plants and Crops, 2nd ed.; Haworth Food & Agricultural Products Press: New York, NY, USA, 1997; p. 568. ISBN 0-8247-0039-2. [Google Scholar]
- Zhou, X.G.; Yu, G.B.; Wu, F.Z. Effects of intercropping cucumber with onion or garlic on soil enzyme activities, microbial communities and cucumber yield. Eur. J. Soil Biol. 2011, 47, 279–287. [Google Scholar] [CrossRef]
- Broz, A.K.; Broeckling, C.D.; De-la-Peña, C.; Lewis, M.R.; Greene, E.; Callaway, R.M.; Sumne, L.W.; Vivanco, J.M. Plant neighbor identity influences plant biochemistry and physiology related to defense. BMC Plant Biol. 2010, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Monaim, M.F.; Abo-Elyousr, K.A.M. Effect of preceding and intercropping crops on suppression of lentil damping-off and root rot disease in New Valley e Egypt. Crop Prot. 2012, 32, 41–46. [Google Scholar] [CrossRef]
- Li, H.S. Principles and Techniques of Plant Physiological Biochemical Experiment, 1st ed.; Higher Education Press: Beijing, China, 2000; pp. 164–165. ISBN 7-04-008076-1. [Google Scholar]
- Gaumann, E. Fusaric acid as a wilt toxin. Phytopathology 1957, 47, 342–357. [Google Scholar]
- Di Pietro, A.; Roncero, M.I.G. Endopolygalacturonase from Fusarium oxysporum f. sp. lycopersici: Purification, characterization, and production during infection of tomato plants. Phytopathology 1996, 86, 1324–1330. [Google Scholar]
- Cao, C.L.; Cui, B.K.; Qin, W.M. Activity changes of several extracellular enzymes in liquid culture of Phellinus mori. Inst. Microbiol. 2011, 30, 275–280. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Parameter | 0 mg·L−1 | 50 mg·L−1 | 100 mg·L−1 | 200 mg·L−1 |
|---|---|---|---|---|
| Leaf number per plant | 21.67 ± 0.58a | 16.33 ± 2.89b | 13.00 ± 1.00c | 11.83 ± 0.29d |
| Max leaf length (cm) | 8.73 ± 0.38a | 6.80 ± 0.61b | 6.47 ± 0.38b | 4.50 ± 0.10c |
| Max leaf width (cm) | 5.43 ± 0.51a | 4.13 ± 0.12b | 3.77 ± 0.55b | 2.93 ± 0.15c |
| Height (cm) | 39.23 ± 0.49a | 33.23 ± 1.48b | 27.33 ± 1.11c | 22.83 ± 0.58d |
| Main root length (cm) | 18.33 ± 1.46a | 17.53 ± 0.42a | 16.77 ± 0.45b | 13.23 ± 0.31c |
| Shoot dry weight (g) | 2.23 ± 0.05a | 1.66 ± 0.11b | 0.92 ± 0.21c | 0.71 ± 0.08c |
| Root dry weight (g) | 0.54 ± 0.03a | 0.39 ± 0.05b | 0.20 ± 0.05c | 0.14 ± 0.04c |
| Root–shoot ratio (%) | 0.24 ± 0.02a | 0.23 ± 0.02a | 0.22 ± 0.01ab | 0.20 ± 0.02b |
| Concentration (mg·L−1) | POD Activity (μ·g−1·min−1) | CAT Activity (mg·g−1·min−1) | MDA Content (μmol·g−1) |
|---|---|---|---|
| 0 | 23.33 ± 0.58a | 0.51 ± 0.02a | 12.77 ± 1.54d |
| 50 | 19.33 ± 0.58b | 0.43 ± 0.04b | 15.89 ± 0.92c |
| 100 | 16.33 ± 1.53c | 0.28 ± 0.02c | 17.94 ± 0.90b |
| 200 | 12 ± 1.73d | 0.19 ± 0.03d | 21.04 ± 0.47a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Q.; Chen, L.; Dong, K.; Dong, Y.; Xiao, J. Cinnamic Acid Inhibited Growth of Faba Bean and Promoted the Incidence of Fusarium Wilt. Plants 2018, 7, 84. https://doi.org/10.3390/plants7040084
Zhao Q, Chen L, Dong K, Dong Y, Xiao J. Cinnamic Acid Inhibited Growth of Faba Bean and Promoted the Incidence of Fusarium Wilt. Plants. 2018; 7(4):84. https://doi.org/10.3390/plants7040084
Chicago/Turabian StyleZhao, Qian, Ling Chen, Kun Dong, Yan Dong, and Jingxiu Xiao. 2018. "Cinnamic Acid Inhibited Growth of Faba Bean and Promoted the Incidence of Fusarium Wilt" Plants 7, no. 4: 84. https://doi.org/10.3390/plants7040084
APA StyleZhao, Q., Chen, L., Dong, K., Dong, Y., & Xiao, J. (2018). Cinnamic Acid Inhibited Growth of Faba Bean and Promoted the Incidence of Fusarium Wilt. Plants, 7(4), 84. https://doi.org/10.3390/plants7040084