Evidence of Adaptation to Recent Changes in Atmospheric CO2 in Four Weedy Species

Abstract
1. Introduction
2. Results
2.1. Seedling Growth
2.2. Dry Mass at Maturity
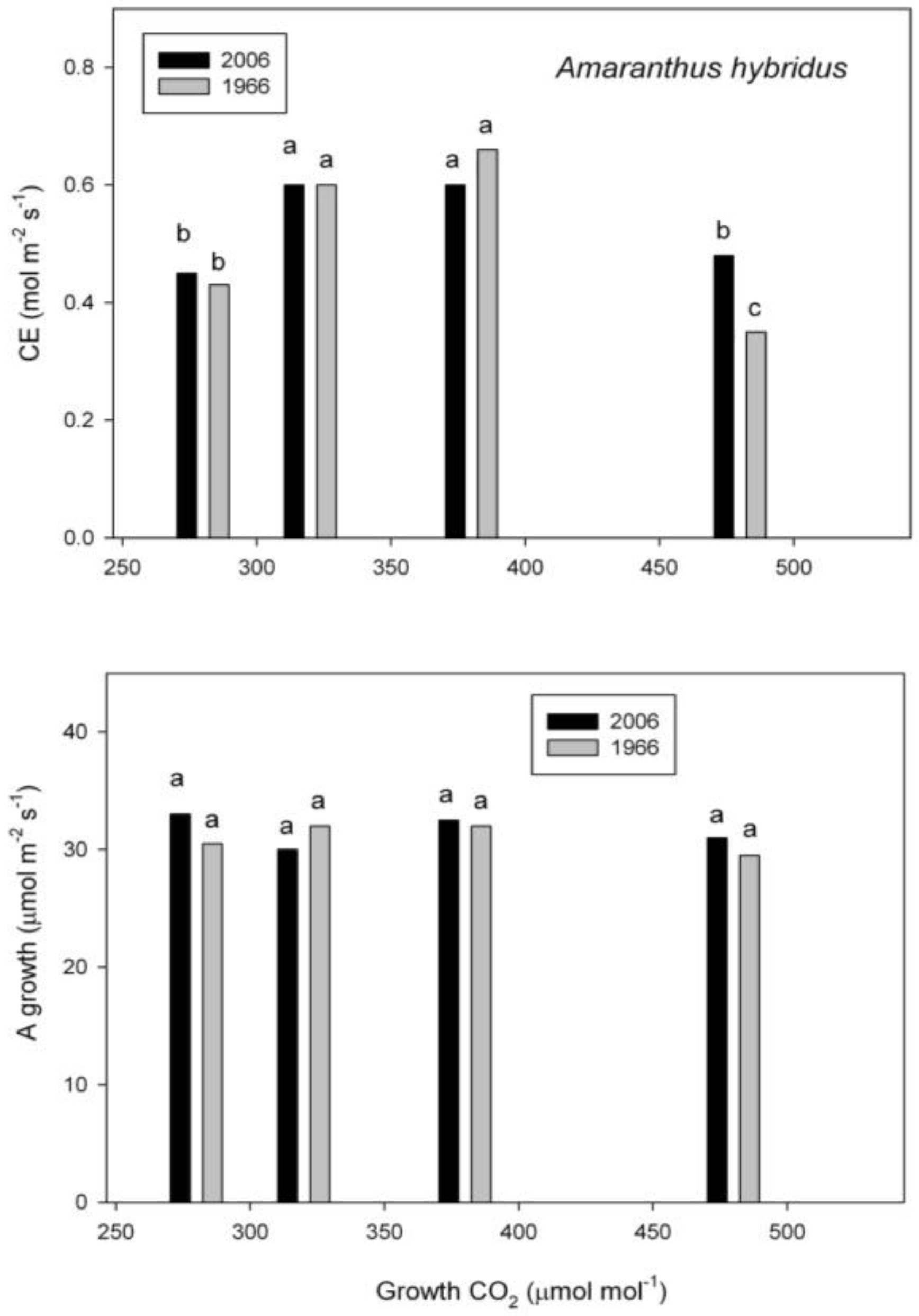
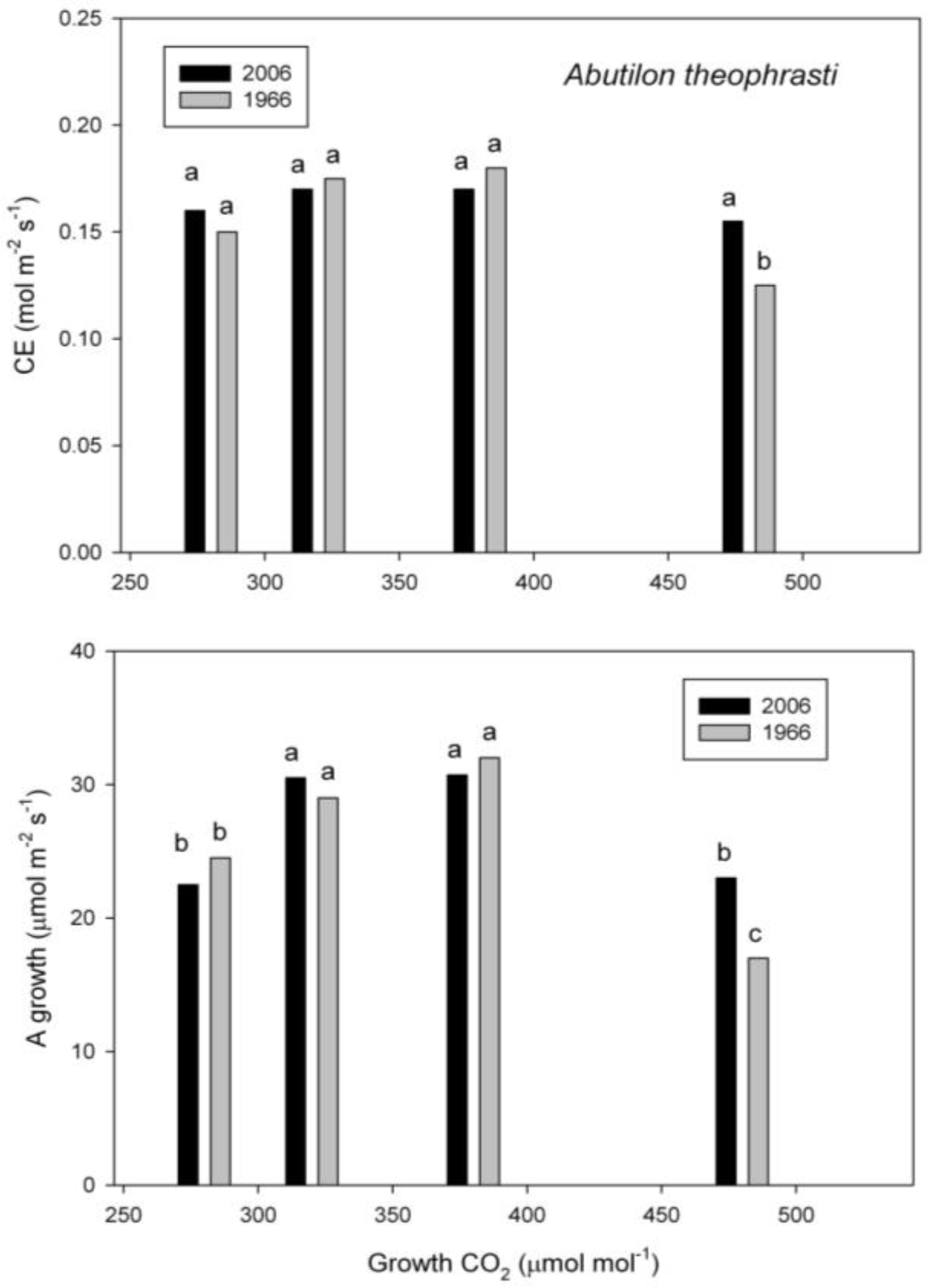
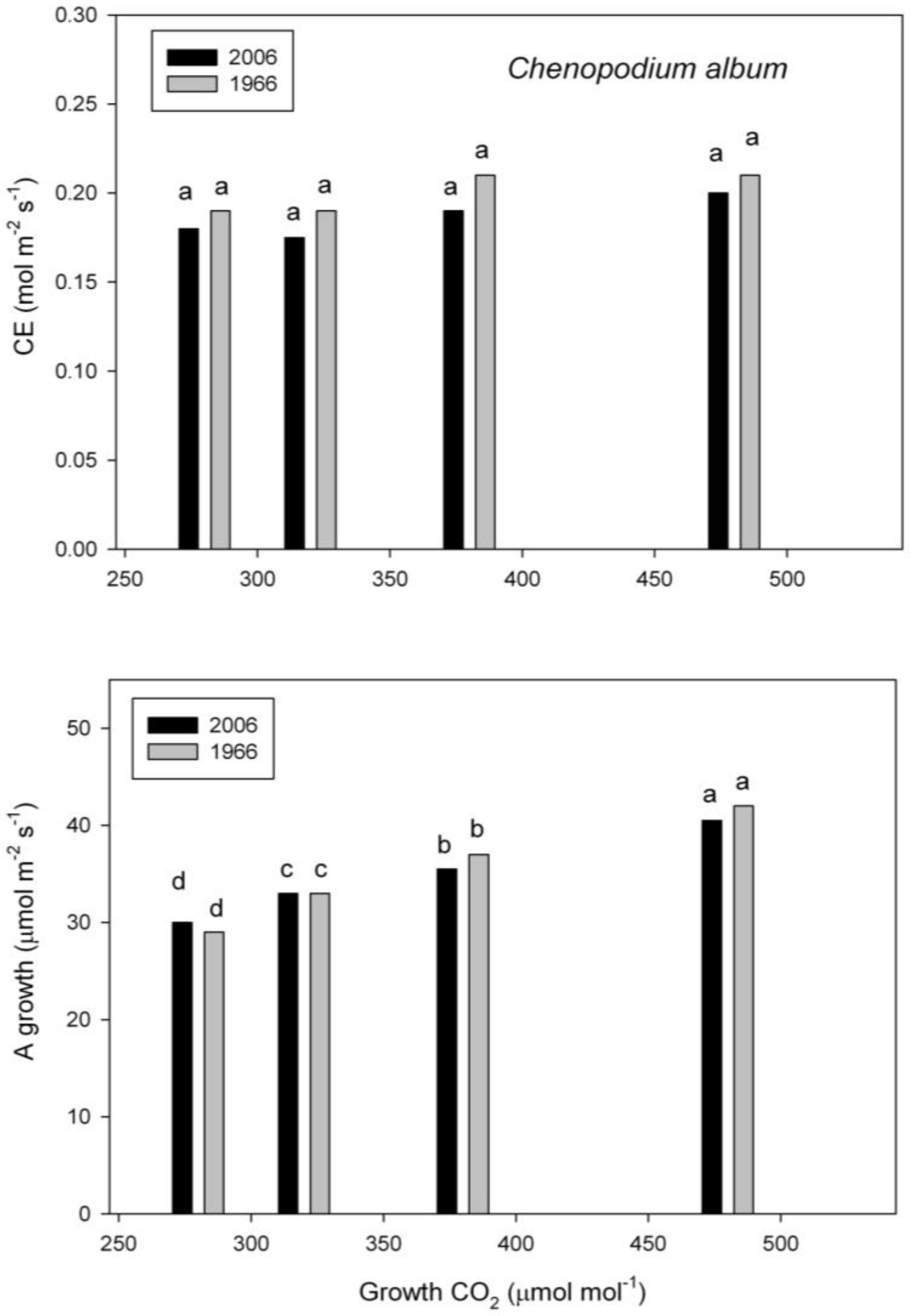
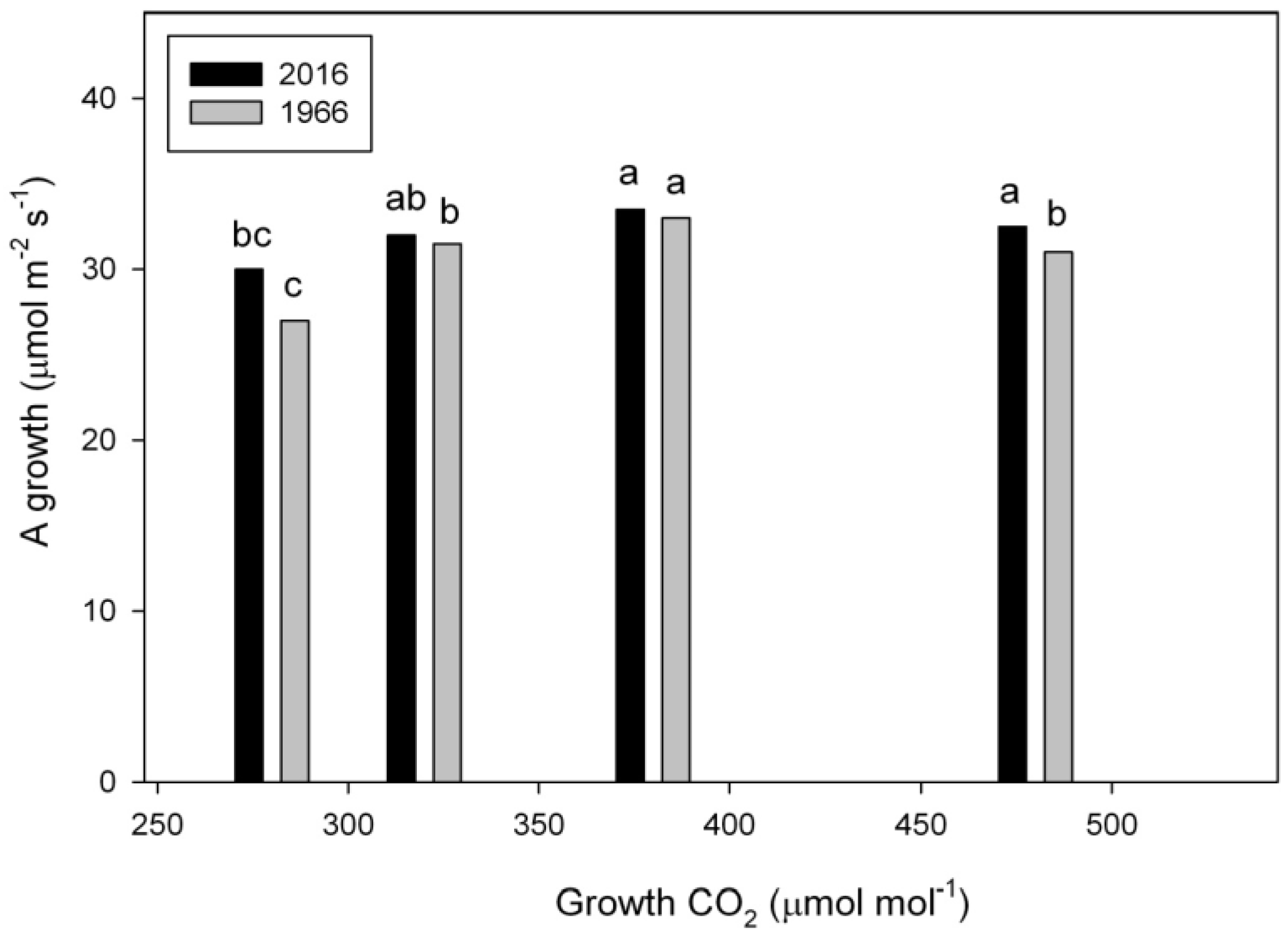
2.3. Photosynthesis
3. Discussion
4. Materials and Methods
4.1. Seedling Growth Rates
4.2. Growth to Maturity
4.3. Photosynthetic Acclimation
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Shaw, R.G.; Etterson, J.R. Rapid climate change and the rate of adaptation: Insight from experimental quantitative genetics. New Phytol. 2012, 195, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.T. Plant fitness in a rapidly changing world. New Phytol. 2015, 210, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A. Contrasting effects of carbon dioxide and irradiance on the acclimation of photosynthesis in developing leaves. Photosynthetica 2000, 38, 83–89. [Google Scholar] [CrossRef]
- ESRL Global Monitoring. Available online: https://www.esrl.noaa.gov/gmd/ccgg/trends/full.html (accessed on 18 January 2018).
- Bunce, J.A. Are annual plants adapted to the current atmospheric concentration of carbon dioxide? Int. J. Plant Sci. 2001, 162, 1261–1266. [Google Scholar] [CrossRef]
- Liu, G.; Yang, C.; Xu, K.; Zhang, Z.; Li, D.; Wu, Z.; Chen, Z. Development of yield and some photosynthetic characteristics during 82 years of genetic improvement of soybean genotypes in northeast China. Aust. J. Crop Sci. 2012, 6, 1416–1422. [Google Scholar]
- Luo, H.H.; Zhang, H.L.; Zhang, Y.L.; Zhang, W.F. Evolution of characteristics related to photosynthesis, growth and yield in some old and new cotton cultivars. Photosynthetica 2017, 55, 301–307. [Google Scholar] [CrossRef]
- Sadras, V.O.; Lawson, C.; Montoro, A. Photosynthetic traits in Australian wheat varieties released between 1958 and 2007. Field Crops Res. 2012, 134, 19–29. [Google Scholar] [CrossRef]
- Koester, R.P.; Nohl, B.M.; Diers, B.W.; Ainsworth, E.A. Has photosynthetic capacity increased with 80 years of soybean breeding? An examination of historical soybean cultivars. Plant Cell Environ. 2016, 39, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Frenk, G.; van der Linden, L.; Mikkelsen, T.N.; Briz, H.; Jorgensen, R.B. Response to multi-generational selection under elevated [CO2] in two temperature regimes suggests enhanced carbon assimilation and increased reproductive output in Brassica napus L. Ecol. Evol. 2013, 3, 1163–1172. [Google Scholar] [CrossRef] [PubMed]
- Alemayehu, F.R.; Frenck, G.; van der Linden, L.; Nikkelsen, T.E.; Jorgensen, R.B. Can barley (Hordeum vulgare L. s.l.) adapt to fast climate changes? A controlled selection experiment. Genet. Res. Crop Evol. 2014, 61, 151–161. [Google Scholar] [CrossRef]
- Fordham, M.; Barnes, J.D.; Bettarini, I.; Polle, A.; Slee, N.; Raines, C.; Miglietta, F.; Raschi, A. The impact of elevated CO2 on growth and photosynthesis in Agrostis canina L. ssp. montelucci adapted to contrasting atmospheric CO2 concentrations. Oecologia 1997, 110, 169–178. [Google Scholar] [CrossRef]
- Polle, A.; McKee, I.; Balschke, L. Altered physiological and growth responses to elevated [CO2] in offspring from holm oak (Quercus ilex L). mother trees with lifetime exposure to naturally elevated [CO2]. Plant Cell Environ. 2001, 24, 1075–1083. [Google Scholar] [CrossRef]
- Newton, P.C.D.; Edwards, G.E. Plant breeding for a changing environment. In Agroecosystems in a Changing Climate; Newton, P.C.D., Carran, R.A., Edwards, G.R., Niklaus, P.A., Eds.; Taylor and Prancis Publishers: London, UK, 2007; pp. 309–319. [Google Scholar]
- Nakamura, I.; Onoda, Y.; Matsushima, N.; Yokoyama, J.; Kawata, M.; Hikosaka, K. Pheotypic and genetic differences in a perennial herb across a natural gradient of CO2 concentration. Oecologia 2011, 165, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A. Elevated carbon dioxide alters the relative fitness of Taraxacum officinale genotypes. Am. J. Plant Sci. 2012, 3, 202–208. [Google Scholar] [CrossRef][Green Version]
- Ward, J.K.; Antonovics, J.; Thomas, R.B.; Strain, B.R. Is atmospheric CO2 a selective agent on model C3 annuals? Oecologia 2000, 123, 330–341. [Google Scholar] [CrossRef] [PubMed]
- Bazzaz, F.A.; Jasienski, M.; Thomas, S.C.; Wayne, P. Microevolutionary responses in experimental populations of plants to CO2-enriched environments: Parallel results from two model systems. Proc. Natl. Acad. Sci. USA 1995, 92, 8161–8165. [Google Scholar] [CrossRef] [PubMed]
- Steinger, R.; Stephan, A.; Schmid, B. Predicting adaptive evolution under elevated atmospheric CO2 in the perennial grass Bromus erectus. Glob. Chang. Biol. 2007, 13, 1028–1039. [Google Scholar] [CrossRef]
- Wieneke, S.; Prati, D.; Barndl, R.; Stocklin, J. Genetic variation in Sanguisorba minor after 6 years in situ selection under elevated CO2. Glob. Chang. Biol. 2004, 10, 1389–1401. [Google Scholar] [CrossRef]
- Springer, C.J.; Ward, J.K. Flowering time and elevated atmospheric CO2. New Phytol. 2007, 176, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A. Limitations to soybean photosynthesis at elevated carbon dioxide in free-air enrichment and open top chamber systems. Plant Sci. 2014, 226, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Massad, R.-S.; Tuzet, A.; Bethenod, O. The effect of temperature on C4-type photosynthesis parameters. Plant Cell Environ. 2007, 30, 1191–1204. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Year of Collection | Growth CO2 | Total DM | LAR | RGR |
|---|---|---|---|---|---|
| A. theophrasti | 1966 | 320 | 4.9a | 139a | 0.29a |
| 1966 | 380 | 5.0a | 134a | 0.28a | |
| 2006 | 320 | 4.0b | 140a | 0.21b | |
| 2006 | 380 | 4.9a | 136a | 0.20b | |
| C. album | 1966 | 320 | 5.2a | 127a | 0.29a |
| 1966 | 380 | 5.3a | 123a | 0.31a | |
| 2006 | 320 | 3.2b | 122a | 0.22b | |
| 2006 | 380 | 5.6a | 120a | 0.29a | |
| D. stramonium | 1966 | 320 | 6.7b | 154a | 0.36a |
| 1966 | 380 | 7.1b | 135b | 0.32b | |
| 2006 | 320 | 7.6b | 145ab | 0.32b | |
| 2006 | 380 | 9.0a | 116c | 0.27c | |
| A. hybridus | 1966 | 320 | 4.1a | 178b | 0.30a |
| 1966 | 380 | 4.4a | 134c | 0.33a | |
| 2006 | 320 | 3.7a | 215a | 0.32a | |
| 2006 | 380 | 3.8a | 165b | 0.29a |
| Species | Year of Collection | Growth CO2 | Total Shoot DM (g) | Seed DM (g) |
|---|---|---|---|---|
| A. theophrasti | 1966 | 320 | 99c | 30c |
| 1966 | 380 | 135b | 41b | |
| 2006 | 320 | 130b | 40b | |
| 2006 | 380 | 153a | 62a | |
| C. album | 1966 | 320 | 46c | na |
| 1966 | 380 | 60b | na | |
| 2006 | 320 | 59b | na | |
| 2006 | 380 | 71a | na | |
| D. stramonium | 1966 | 320 | 315ab | 172b |
| 1966 | 380 | 350a | 193a | |
| 2006 | 320 | 289b | 172b | |
| 2006 | 380 | 351a | 205a | |
| A. hybridus | 1966 | 320 | 186a | 78a |
| 1966 | 380 | 191a | 82a | |
| 2006 | 320 | 90b | 62b | |
| 2006 | 380 | 102b | 60b |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bunce, J. Evidence of Adaptation to Recent Changes in Atmospheric CO2 in Four Weedy Species. Plants 2018, 7, 12. https://doi.org/10.3390/plants7010012
Bunce J. Evidence of Adaptation to Recent Changes in Atmospheric CO2 in Four Weedy Species. Plants. 2018; 7(1):12. https://doi.org/10.3390/plants7010012
Chicago/Turabian StyleBunce, James. 2018. "Evidence of Adaptation to Recent Changes in Atmospheric CO2 in Four Weedy Species" Plants 7, no. 1: 12. https://doi.org/10.3390/plants7010012
APA StyleBunce, J. (2018). Evidence of Adaptation to Recent Changes in Atmospheric CO2 in Four Weedy Species. Plants, 7(1), 12. https://doi.org/10.3390/plants7010012