Cuticular Waxes of Arabidopsis thaliana Shoots: Cell-Type-Specific Composition and Biosynthesis

Abstract
1. Introduction
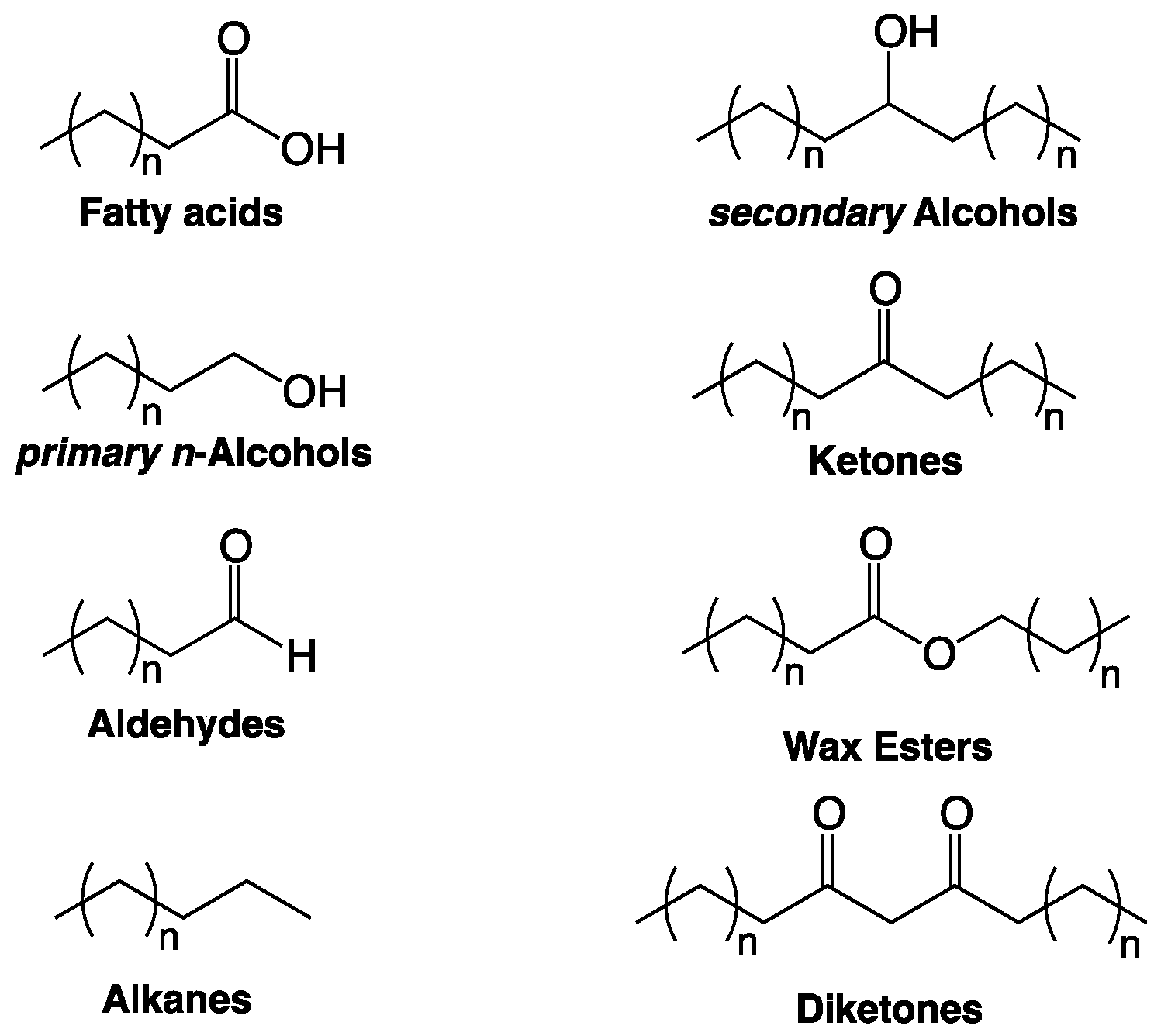
2. Composition of Cuticular Wax Covering Arabidopsis Trichomes
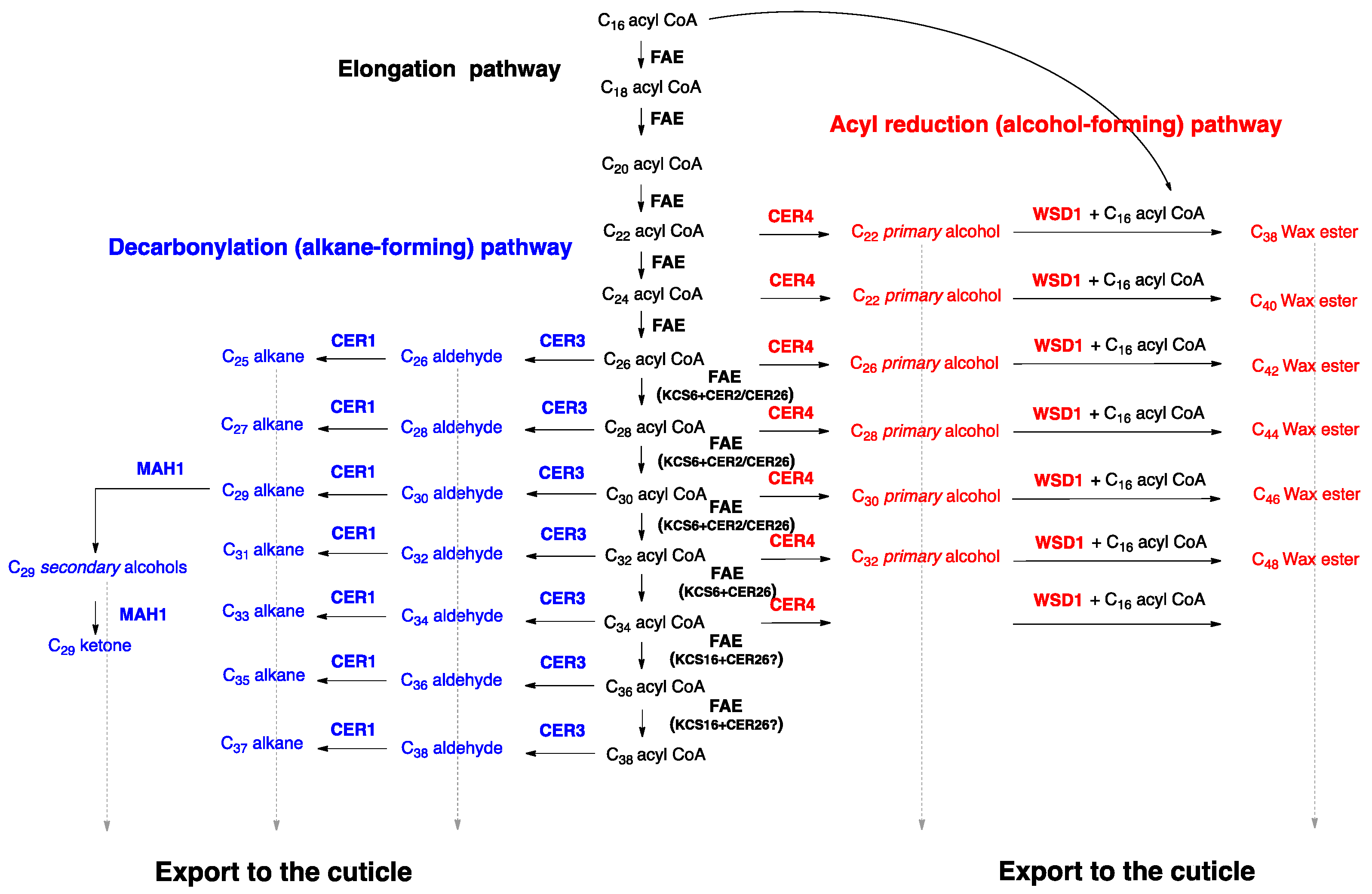
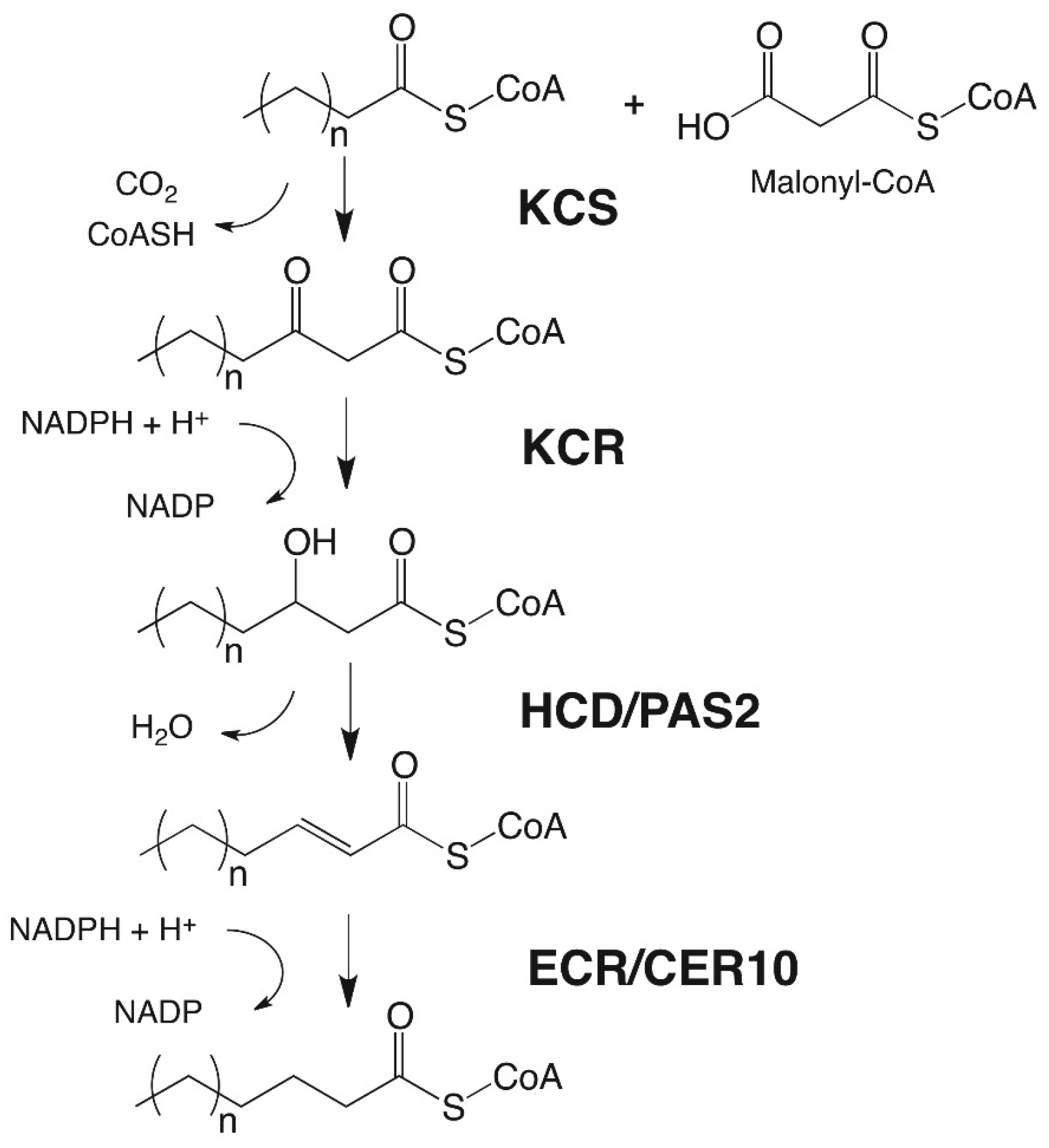
3. Wax Biosynthesis in Arabidopsis Trichome Cells
4. Extra-Long Compounds in the Wax Mixtures of Diverse Plant Species
5. Possible Functions of Extra-Long-Chain Compounds in Trichome Wax
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ramsay, N.A.; Glover, B.J. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 2005, 10, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Busta, L.; Hegebarth, D.; Kroc, E.; Jetter, R. Changes in cuticular wax coverage and composition on developing Arabidopsis leaves are influenced by wax biosynthesis gene expression levels and trichome density. Planta 2016. [Google Scholar] [CrossRef] [PubMed]
- Kearns, E.V.; Assmann, S.M. The guard cell-environment connection. Plant Physiol. 1993, 102, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Autran, D.; Jonak, C.; Belcram, K.; Beemster, G.T.S.; Kronenberger, J.; Grandjean, O.; Inzé, D.; Traas, J. Cell numbers and leaf development in Arabidopsis: A functional analysis of the struwwelpeter gene. EMBO J. 2002, 21, 6036–6049. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.J.; Wang, E.; Shepherd, R.W. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 2004, 93, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Hegebarth, D.; Buschhaus, C.; Wu, M.; Bird, D.; Jetter, R. The composition of surface wax on trichomes of Arabidopsis thaliana differs from wax on other epidermal cells. Plant J. 2016, 1–13. [Google Scholar] [CrossRef]
- Hegebarth, D.; Buschhaus, C.; Joubes, J.; Thoroval, D.; Bird, D.; Jetter, J. Arabidopsis ketoacyl-CoA synthase 16 forms C36/C38 acyl precursors for leaf trichome and pavement surface wax. Plant Cell Environ. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kolattukudy, P.E. Biopolyester Membranes of Plants: Cutin and suberin. Science 1980, 208, 990–1000. [Google Scholar] [CrossRef] [PubMed]
- Graça, J.; Schreiber, L.; Rodrigues, J.; Pereira, H. Glycerol and glyceryl esters of ω-hydroxyacids in cutins. Phytochemistry 2002, 61, 205–215. [Google Scholar] [CrossRef]
- Xiao, F.; Goodwin, S.M.; Xiao, Y.; Sun, Z.; Baker, D.; Tang, X.; Jenks, M.A.; Zhou, J.-M. Arabidopsis CYP86A2 represses Pseudomonas syringae type III genes and is required for cuticle development. EMBO J. 2004, 23, 2903–2913. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, M.; Sun, Y.; Hegebarth, D.; Li, T.; Jetter, R.; Wang, Z. Molecular characterization of TaFAR1 involved in primary alcohol biosynthesis of cuticular wax in hexaploid wheat. Plant Cell Physiol. 2015, 56, 1944–1961. [Google Scholar] [CrossRef] [PubMed]
- Pulsifer, I.; Kluge, S.; Rowland, O. Arabidopsis LONG-CHAIN ACYL-COA SYNTHETASE 1 (LACS1), LACS2, and LACS3 facilitate fatty acid uptake in yeast. Plant Physiol. Biochem. 2012, 51, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Joubes, J.; Raffalele, S.; Bourdenx, B.; Garcia, C.; Laroche-Traineau, J.; Morea, P.; Domergue, F.; Lessire, R. The VLCFA elongase gene family in Arabidopsis thaliana: Phylogenetic analysis, 3D modelling and expression profiling. Plant Mol. Biol. 2008, 67, 547–566. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, F.; Wu, X.; Li, F.; Haslam, R.P.; Markham, J.E.; Zheng, H.; Napier, J.; Kunst, L. Functional characterization of the Arabidopsis beta-ketoacyl-coenzyme A reductase candidates of the fatty acid elongase. Plant Physiol. 2009, 150, 1174–1191. [Google Scholar] [CrossRef] [PubMed]
- Domergue, F.; Chevalier, S.; Creach, A.; Cassagne, C.; Lessire, R. Purification of the acyl-CoA elongase complex from developing rapeseed and characterization of the 3-ketoacyl-CoA synthase and the 3-hydroxyacyl-CoA dehydratase. Lipids 2000, 35, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Gable, K.; Kohlwein, S.D.; Beaudoin, F.; Napier, J.A.; Dunn, T.M. The Saccharomyces cerevisiae YBR159w gene encodes the 3-ketoreductase of the microsomal fatty acid elongase. J. Biol. Chem. 2002, 277, 35440–35449. [Google Scholar] [CrossRef] [PubMed]
- Kunst, L.; Samuels, L. Plant cuticles shine: advances in wax biosynthesis and export. Curr. Opin. Plant Biol. 2009, 12, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Acyl-lipid metabolism. Arabidopsis Book 2013, 11, e0161. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.A.; Kunst, L. Very-long-chain fatty acid biosynthesis is controlled through the expression and specificity of the condensing enzyme. Plant J. 1997, 12, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Gable, K.; Beaudoin, F.; Cahoon, E.; Jaworski, J.; Napier, J.A.; Dunn, T.M. Members of the Arabidopsis FAE1-like 3-ketoacyl-CoA synthase gene family substitute for the elop proteins of Saccharomyces cerevisiae. J. Biol. Chem. 2006, 281, 9018–9029. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Rowland, O.; Kunst, L. Disruptions of the Arabidopsis Enoyl-CoA reductase gene reveal an essential role for very-long-chain fatty acid synthesis in cell expansion during plant morphogenesis. Plant Cell 2005, 17, 1467–1481. [Google Scholar] [CrossRef] [PubMed]
- Haslam, T.M.; Mañas Fernández, A.; Zhao, L.; Kunst, L. Arabidopsis ECERIFERUM2 is a component of the fatty acid elongation machinery required for fatty acid extension to exceptional lengths. Plant Physiol. 2012. [Google Scholar] [CrossRef]
- Pascal, S.; Bernard, A.; Sorel, M.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; Lessire, R.; Domergue, F.; Joubès, J. The Arabidopsis cer26 mutant, like the cer2 mutant, is specifically affected in the very-long-chain fatty acid elongation process. Plant J. 2013, 73, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Haslam, T.M.; Haslam, R.; Thoraval, D.; Pascal, S.; Delude, C.; Domergue, F.; Fernández, A.M.; Beaudoin, F.; Napier, J.A.; Kunst, L.; Joubès, J. ECERIFERUM2-LIKE proteins have unique biochemical and physiological functions in very-long-chain fatty acid elongation. Plant Physiol. 2015, 167, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Haslam, T.M.; Gerelle, W.K.; Graham, S.W.; Kunst, L. The Unique Role of the ECERIFERUM2-LIKE Clade of the BAHD Acyltransferase Superfamily in Cuticular Wax Metabolism. Plants 2017, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Domergue, F.; Pascal, S.; Jetter, R.; Renne, C.; Faure, J.-D.; Haslam, R.P.; Napier, J.A.; Lessire, R.; Joubès, J. Reconstitution of plant alkane biosynthesis in yeast demonstrates that Arabidopsis ECERIFERUM1 and ECERIFERUM3 are core components of a very-long-chain alkane synthesis complex. Plant Cell 2012, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cheesbrough, T.M.; Kolattukudy, P.E. Alkane biosynthesis by decarbonylation of aldehydes catalyzed by a particulate preparation from Pisum sativum. Proc. Natl. Acad. Sci. USA 1984, 81, 6613–6617. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Belhaddad, F.; Kolattukudy, P. Solubilization, partial purification, and characterization of a fatty aldehyde decarbonylase from a higher plant, Pisum sativum. Arch. Biochem. Biophys. 2000, 377, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Greer, S.; Wen, M.; Bird, D.; Wu, X.; Samuels, L.; Kunst, L.; Jetter, R. The cytochrome P450 enzyme CYP96A15 is the midchain alkane hydroxylase responsible for formation of secondary alcohols and ketones in stem cuticular wax of Arabidopsis. Plant Physiol. 2007, 145, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Rowland, O.; Zheng, H.; Hepworth, S.R.; Lam, P.; Jetter, R.; Kunst, L. CER4 encodes an alcohol-forming fatty acyl-coenzyme A reductase involved in cuticular wax production in Arabidopsis. Plant Physiol. 2006, 142, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wu, X.; Lam, P.; Bird, D.; Zheng, H.; Samuels, L.; Jetter, R.; Kunst, L. Identification of the wax ester synthase/acyl-coenzyme A: Diacylglycerol acyltransferase WSD1 required for stem wax ester biosynthesis in Arabidopsis. Plant Physiol. 2008, 148, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Riederer, M. Thermodynamics of the water permeability of plant cuticles: Characterization of the polar pathway. J. Exp. Bot. 2006, 57, 2937–2942. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, S.M.; Rashotte, A.M.; Rahman, M.; Feldmann, K.A.; Jenks, M.A. Wax constituents on the inflorescence stems of double eceriferum mutants in Arabidopsis reveal complex gene interactions. Phytochemistry 2005, 66, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Kunst, L.; Jetter, R.; Samuels, A.L. Biosynthesis and transport of plant cuticular waxes. Biol. Plant Cuticle 2006, 23, 182–215. [Google Scholar]
- Koornneef, M.; Hanhart, C.J.; Thiel, F. A Genetic and phenotypic description of Eceriferum (cer) mutants in Arabidopsis thaliana. J. Hered. 1989, 80, 118–122. [Google Scholar] [CrossRef]
- Xue, Y.; Xiao, S.; Kim, J.; Lung, S.-C.; Chen, L.; Tanner, J.A.; Suh, M.C.; Chye, M.-L. Arabidopsis membrane-assoicayted acyl-CoA-binding protein ACBP1 is involved in stem cuticle formation. J. Exp. Bot. 2014, 65, 5473–5483. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.M.; Gray, J.E. Signals from the cuticle affect epidermal cell differentiation. New Phytol. 2003, 157, 9–23. [Google Scholar] [CrossRef]
- Marks, M.D.; Wenger, J.P.; Gilding, E.; Jilk, R.; Dixon, R.A. Transcriptome analysis of Arabidopsis wild-type and gl3-sst sim trichomes identifies four additional genes required for trichome development. Mol. Plant 2009, 2, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Jeffree, C.E. The fine structure of the plant cuticle. In Biology of the Plant Cuticle; Riederer, M., Müller, C., Eds.; Blackwell Publishing Ltd: Oxford, UK, 2006; pp. 11–125. [Google Scholar]
- Jetter, R.; Kunst, L.; Samuels, A.L. Composition of plant cuticular waxes. Annu. Plant Rev. 2007, 23, 145–181. [Google Scholar] [CrossRef]
- Schreiber, L. Review of sorption and diffusion of lipophilic molecules in cuticular waxes and the effects of accelerators on solute mobilities. J. Exp. Bot. 2006, 57, 2515–2523. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Nikolau, B.J.; Schnable, P.S. Developmental and hormonal regulation of the arabidopsis CER2 gene that codes for a nuclear-localized protein required for the normal accumulation of cuticular waxes. Plant Physiol. 1997, 115, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Bourdenx, B.; Bernard, A.; Domergue, F.; Pascal, S.; Léger, A.; Roby, D.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; Lessire, R.; Joubès, J. Overexpression of Arabidopsis ECERIFERUM1 promotes wax very-long-chain alkane biosynthesis and influences plant response to biotic and abiotic stresses. Plant Physiol. 2011, 156, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Ma, S.; Lee, I.; Kim, J.; Liu, S. Saline-induced changes of epicuticular waxy layer on the Puccinellia tenuiflora and Oryza sativa leave surfaces. Biol. Res. 2015, 48, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fahn, A. Structural and functional-properties of trichomes of xeromorphic leaves. Ann. Bot. 1986, 57, 631–637. [Google Scholar] [CrossRef]
- Guhling, O.; Kinzler, C.; Dreyer, M.; Bringmann, G.; Jetter, R. Surface composition of myrmecophilic plants: Cuticular wax and glandular trichomes on leaves of Macaranga tanarius. J. Chem. Ecol. 2005, 31, 2323–2341. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, G.; Pfeifhofer, H.W.; Stabentheiner, E.; Sawidis, T. Glandular hairs of Sigesbeckia jorullensis Kunth (Asteraceae): Morphology, histochemistry and composition of essential oil. Ann. Bot. 2002, 89, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Fernández, V.; Khayet, M.; Montero-Prado, P.; Heredia-Guerrero, J.A.; Liakopoulos, G.; Karabourniotis, G.; Del Río, V.; Domínguez, E.; Tacchini, I.; Nerín, C.; et al. New insights into the properties of pubescent surfaces: Peach fruit as a model. Plant Physiol. 2011, 156, 2098–2108. [Google Scholar] [CrossRef] [PubMed]
- Hemmers, H.; Gulz, P.G. Waxes of five Euphorbia Species. Phytochemistry 1986, 25, 2103–2107. [Google Scholar]
- Clark, L.V.; Ryan Stewart, J.; Nishiwaki, A.; Toma, Y.; Kjeldsen, J.B.; Jörgensen, U.; Zhao, H.; Peng, J.; Yoo, J. H.; Heo, K.; et al. Genetic structure of Miscanthus sinensis and Miscanthus sacchariflorus in Japan indicates a gradient of bidirectional but asymmetric introgression. J. Exp. Bot. 2015, 66, 4213–4225. [Google Scholar] [CrossRef] [PubMed]
- Motomura, H. Distribution of silicified cells in the leaf blades of Pleioblastus chino (Franchet et Savatier) Makino (Bambusoideae). Ann. Bot. 2000, 85, 751–757. [Google Scholar] [CrossRef]
- Hameed, M.; Ashraf, M.; Naz, N.; Nawaz, T.; Batool, R.; Sajid Aqeel Ahmad, M.; Ahmad, F.; Hussain, M. Anatomical adaptations of Cynodon dactylon (L.) Pers. from the salt range (Pakistan) to salinity stress. II. leaf anatomy. Pakistan J. Bot. 2013, 45, 133–142. [Google Scholar]
- Gruber, M.; Skadhauge, B.; Yu, M.; Muir, A.; Richards, K. and flavonoids within a Lotus germplasm collection. Can. J. Plant Sci. 2008, 88, 121–132. [Google Scholar] [CrossRef]
- Retallack, B.; Willison, J.H.M. Morphology, anatomy, and distribution of capitate glandular trichomes on selected Trifolium species. Crop Sci. 1988, 28, 677–680. [Google Scholar] [CrossRef]
- Ågren, J.; Schemske, D.W. Artificial selection on trichome number in Brassica rapa. Theor. Appl. Genet. 1992, 83, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Farnese, F.S.; Oliveira, J.A.; Lima, F.S.; Leao, G.A.; Gusman, G.S.; Silva, L.C. Evaluation of the potential of Pistia stratiotes L. (water lettuce) for bioindication and phytoremediation of aquatic environments contaminated with arsenic. Braz. J. Biol. 2014, 74, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Titah, H.S.; Abdullah, S.R.S.; Mushrifah, I.; Anuar, N.; Basri, H.; Mukhlisin, M. Effect of applying rhizobacteria and fertilizer on the growth of Ludwigia octovalvis for arsenic uptake and accumulation in phytoremediation. Ecol. Eng. 2013, 58, 303–313. [Google Scholar] [CrossRef]
- Dodd, R.S.; Rafii, Z.A.; Power, A.B. Ecotypic adaptation in Austrocedrus chilensis in cuticular hydrocarbon composition. New Phytol. 1998, 138, 699–708. [Google Scholar] [CrossRef]
- Jetter, R.; Riederer, M. Cuticular waxes from the leaves and fruit capsules of eight Papaveraceae species. Can. J. Bot. 1996, 74, 419–430. [Google Scholar] [CrossRef]
- Zhang, Y.; Togamura, Y.; Otsuki, K. Study on the n-alkane patterns in some grasses and factors affecting the n-alkane patterns. J. Agric. Sci. 2004, 142, 469–475. [Google Scholar] [CrossRef]
- Bugalho, M.N.; Dove, H.; Kelman, W.; Wood, J.T.; Mayes, R.W. Plant wax alkanes and alcohols as herbivore diet composition markers. J. Range Manag. 2004, 57, 259–268. [Google Scholar] [CrossRef]
- Buschhaus, C.; Herz, H.; Jetter, R. Chemical composition of the epicuticular and intracuticular wax layers on adaxial sides of Rosa canina leaves. Ann. Bot. 2007, 100, 1557–1564. [Google Scholar] [CrossRef] [PubMed]
- Dragota, S.; Riederer, M. Epicuticular wax crystals of Wollemia nobilis: Morphology and chemical composition. Ann. Bot. 2007, 100, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Luo, G.; Meyers, P.A.; Gu, Y.; Wang, H.; Xie, S. Leaf wax n-alkane chemotaxonomy of bamboo from a tropical rain forest in Southwest China. Plant Syst. Evol. 2012, 298, 731–738. [Google Scholar] [CrossRef]
- Domínguez, E.; Cuartero, J.; Heredia, A. An overview on plant cuticle biomechanics. Plant Sci. 2011, 181, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Sheng, G.; Liu, X.; Li, C.; Fu, J. Molecular and carbon and hydrogen isotopic composition of n-alkanes in plant leaf waxes. Org. Geochem. 2005, 36, 1405–1417. [Google Scholar] [CrossRef]
- Diefendorf, A.F.; Freeman, K.H.; Wing, S.L.; Graham, H. V. Production of n-alkyl lipids in living plants and implications for the geologic past. Geochim. Cosmochim. Acta 2011, 75, 7472–7485. [Google Scholar] [CrossRef]
- Gao, L.; Huang, Y. Inverse gradients in leaf wax δD and δ13C values along grass blades of Miscanthus sinensis: Implications for leaf wax reproduction and plant physiology. Oecologia 2013, 172, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Nadiminti, P.P.; Rookes, J.E.; Boyd, B.J.; Cahill, D. M. Confocal laser scanning microscopy elucidation of the micromorphology of the leaf cuticle and analysis of its chemical composition. Protoplasma 2015. [Google Scholar] [CrossRef] [PubMed]
- Mihailova, A.; Abbado, D.; Pedentchouk, N. Differences in n-alkane profiles between olives and olive leaves as potential indicators for the assessment of olive leaf presence in virgin olive oils. Eur. J. Lipid Sci. Technol. 2015, 117, 1480–1485. [Google Scholar] [CrossRef]
- Mitra, S.; Sarkar, N.; Barik, A. Long-chain alkanes and fatty acids from Ludwigia octovalvis weed leaf surface waxes as short-range attractant and ovipositional stimulant to Altica cyanea (Weber) (Coleoptera: Chrysomelidae). Bull. Entomol. Res. 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Von Wettstein-Knowles, P. Analyses of barley spike mutant waxes identify alkenes, cyclopropanes and internally branched alkanes with dominating isomers at carbon 9. Plant J. 2007, 49, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Wollrab, V. Olefine und Paraffine aus den Wachsen einiger Pflanzen der Familie Rosaceae. Ueber Naturwachse 1986, 33, 1584–1600. [Google Scholar]
- Wang, W.; Zhang, Y.; Xu, C.; Ren, J.; Liu, X.; Black, K.; Gai, X.; Wang, Q.; Ren, H. Cucumber ECERIFERUM1 (CsCER1), which influences the cuticle properties and drought tolerance of cucumber, plays a key role in VLC alkanes biosynthesis. Plant Mol. Biol. 2015, 87, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Leide, J.; Hildebrandt, U.; Reussing, K.; Riederer, M.; Vogg, G. The developmental pattern of tomato fruit wax accumulation and its impact on cuticular transpiration barrier properties: Effects of a deficiency in a beta-ketoacyl-coenzyme A synthase (LeCER6). Plant Physiol. 2007, 144, 1667–1679. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, G.; Murelli, C.; Ottaviano, E. Maize pollen lipids. Phytochemistry 1990, 29, 739–744. [Google Scholar] [CrossRef]
- Bortolomeazzi, R.; Berno, P.; Pizzale, L.; Conte, L.S. Sesquiterpene, alkene, and alkane hydrocarbons in virgin olive oils of different varieties and geographical origins. J. Agric. Food Chem. 2001, 49, 3278–3283. [Google Scholar] [CrossRef] [PubMed]
- Piper, S.H.; Chibnall, A.C.; Williams, E. Melting-points and long crystal spacings of the higher primary alcohols and n-fatty acids. Biochem. J. 1934, 28, 2175–2188. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Merk, S. Phase behaviour and crystallinity of plant cuticular waxes studies by Fourier transform infrared spectroscopy. Planta 1998, 204, 44–53. [Google Scholar] [CrossRef]
- Jetter, R.; Riederer, M. Localization of the transpiration barrier in the epi- and intracuticular waxes of eight plant species: Water transport resistances are associated with fatty acyl rather than alicyclic components. Plant Physiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Karabourniotis, G. Epicuticular Phenolics Over Guard Cells: Exploitation for in situ stomatal counting by fluorescence microscopy and combined image analysis. Ann. Bot. 2001, 87, 631–639. [Google Scholar] [CrossRef]
- Gray, J.E.; Holroyd, G.H.; van der Lee, F.M.; Bahrami, R.; Sijmons, P.C.; Woodward, F.I.; Schuch, W.; Hetherington, A.M. The HIC signalling pathway links CO2 perception to stomatal development. Nature 2000, 408, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Geisler, M.; Nadeau, J.; Sack, F.D. Oriented asymmetric divisions that generate the stomatal spacing pattern in arabidopsis are disrupted by the too many mouths mutation. Plant Cell 2000, 12, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, J.; Sack, F.D. Stomatal development in Arabidopsis. Arabidopsis Book 2002, 1, e0066. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Sack, F.D. The too many mouths and four lips mutations affect stomatal production in Arabidopsis. Plant Cell 1995, 7, 2227–2239. [Google Scholar] [CrossRef] [PubMed]
- Berger, D.; Altmann, T. A subtilisin-like serine protease involved in the regulation of stomatal density and distribution in Arabidopsis thaliana. Genes Dev. 2000, 14, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Von Groll, U.; Berger, D.; Altmann, T. The subtilisin-like serine protease SDD1 mediates cell-to-cell signaling during Arabidopsis stomatal development. Plant Cell 2002, 14, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.E.; Hetherington, A.M. Plant development: YODA the stomatal switch. Curr. Biol. 2004, 14, 488–490. [Google Scholar] [CrossRef] [PubMed]
- Hunt, L.; Gray, J.E. The signaling peptide EPF2 controls asymmetric cell divisions during stomatal development. Curr. Biol. 2009, 19, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Kajita, R.; Torii, K.U.; Bergmann, D.C.; Kakimoto, T. The secretory peptide gene EPF1 enforces the stomatal one-cell-spacing rule. Genes Dev. 2007, 21, 1720–1725. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Projected Surface Area of Blade (mm2) | Number of | Surface Area of | |||
|---|---|---|---|---|---|
| Trichomes (Blade−1) | Guard Cell Pairs (Blade−1) | Pavement Cells (Blade−1) | Pavement Cells (μm2 cell−1) | Trichome Cells (μm2 cell−1) | |
| 138 | 72 | 10366 | 29602 | 4646 | 40000 |
| Plant Species | Family | Tissue | Analyzed Compound Classes | Compound Class | C max. | Carbon Chain Length Range | Abundance of C36/C35 | Abundance of C38/C37 | Climatic Zone | Trichomes | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Euphorbia characias | Euphorbiaceae | Leaves | Complete wax profile | Alkanes | 31 | C19–C37 | + | + | Temperate | N/A | [48] |
| Aldehydes | 31 | C24–C36 | + | N/A | |||||||
| Euphorbia cyparissias | Alkanes | 31 | C19–C37 | ++ | + | ||||||
| Aldehydes | 31 | C24–C36 | + | N/A | |||||||
| Euphorbia lathyris | Alkanes | 31 | ++ | + | |||||||
| Aldehydes | 31 | C24–C36 | + | N/A | |||||||
| Euphorbia niccaensis | Alkanes | 31 | C19–C37 | + | + | ||||||
| Euphorbia peplus | Alkanes | 31 | C19–C37 | + | N/A | ||||||
| Austrocedrus chilensis | Cupressaceae | Leaves | Alkanes only | Alkanes | 33 | C21–C37 | ++++ | + | Temperate to Subtropical | N/A | [57] |
| Eschscholzia california | Papaveraceae | Leaves | Complete wax profile | Alkanes | 29 | C21–C37 | + | N/A | Temperate | N/A | [58] |
| Papaver orientale | Leaves | 29 | C21–C35 | + | N/A | ||||||
| Papaver somniferum | Capsules | 29 | C21–C37 | + | N/A | ||||||
| Miscanthus sinensis | Poaceae | Leaves | Alkanes only | Alkanes | 31 | C25–C35 | +++ | N/A | Temperate | Yes [49] | [59] |
| Senescent leaves | 31 | +++ | N/A | ||||||||
| Stems | 31 | ++++ | N/A | No [49] | |||||||
| Inflorescence | 31 | ++ | N/A | N/A | |||||||
| Pleioblastus chino | Leaves | 31 | ++++ | N/A | Yes [50] | ||||||
| Sasa nipponica | Leaves | 31 | +++ | N/A | N/A | ||||||
| Senescent leaves | 31 | +++ | N/A | ||||||||
| Zoysia japonica | Leaves | 31 | ++ | N/A | N/A | ||||||
| Senescent leaves | 31 | ++++ | N/A | ||||||||
| Austrodanthonia pilosa | Poaceae | Leaf blades | Alkanes, prim. alcohols | Alkanes | 31 | C25–C35 | ++ | N/A | Temperate | N/A | [60] |
| Austrodanthonia racemosa | 33 | + | N/A | ||||||||
| Axonopus fissifolius | Shoot | 33 | ++++ | N/A | Subtropical | ||||||
| Bothriochloa macra | Leaves | 27 | ++ | N/A | Temperate | ||||||
| Bromus catharticus | Shoot | 29 | ++ | N/A | |||||||
| Chloris gayana | Leaves | 31 | ++ | N/A | Subtropical | N/A | |||||
| Shoot | 33 | ++++ | N/A | ||||||||
| Cynodon dactylon | 33 | +++ | N/A | Yes [51] | |||||||
| Digitaria didactyla | 33 | ++++ | N/A | N/A | |||||||
| Elymus scaber | Leaves | 31 | ++ | N/A | Temperate | N/A | |||||
| Festuca arundinacea | Shoot | 31 | + | N/A | N/A | ||||||
| Imperata cylindrica | 31 | +++ | N/A | Subtropical | |||||||
| Lotus corniculatus "Prostate" | Shoot | 29 | + | N/A | Temperate | No [52] | |||||
| Lotus pedunculatus cv. Maku | 29 | + | N/A | Yes [52] | |||||||
| Microlaena stipoides | Leaves | 31 | ++ | N/A | N/A | ||||||
| Paspalum dilatatum | Shoot | 33 | ++++ | N/A | Subtropical | ||||||
| Paspalum notatum | 35 | ++++ | N/A | ||||||||
| Pennisetum clandestinum | 35 | ++++ | N/A | N/A | |||||||
| Phalaris aquatica | 29 | +++ | N/A | Temperate | N/A | ||||||
| Setaria anceps | 27 | ++ | N/A | Subtropical | |||||||
| Sporobolus indicus cv. Major | 33 | ++++ | N/A | N/A | |||||||
| Themeda australis | Leaves | 31/33 | +++ | N/A | Temperate | ||||||
| Trifolium repens | Fabaceae | Shoot | 31 | + | N/A | Yes [53] | |||||
| Vulpia myuros | Poaceae | 31 | + | N/A | N/A | ||||||
| Brassica spp. | Brassicales | Leaves | Alkanes only | Alkanes | N/A | C17–C35 | N/A | N/A | Temperate | Yes [54] | [37] |
| Pisum sativum | Fabaceae | N/A | |||||||||
| Rosa canina | Rosaceae | Leaves | Complete wax profile | Sec. alcohols | N/A | C29–C35 | + | N/A | Temperate | N/A | [61] |
| Wollemia nobilis | Araucariaceae | Leaves | Alkanes only | Alkanes | N/A | C33–C35 | N/A | N/A | Temperate | N/A | [62] |
| Bambusa bambusa | Poaceae | Leaves | alkanes only | Alkanes | 31/33 | C23–C35 | +++ | N/A | Tropical | N/A | [63] |
| Bambusa dendrocalamopsis | 29 | + | N/A | ||||||||
| Bambusa dendrocalamus | 29 | ++ | |||||||||
| Alternanthera dentata | Amaranthaceae | Leaves | Alkanes only | Alkanes | 29 | C22–C35 | ++ | N/A | Subtropical | N/A | [65] |
| Alternanthera versicolor | 31 | C18–C35 | + | N/A | |||||||
| Araucaria cunninghamii | Araucariaceae | 31 | C22–C35 | ++ | N/A | ||||||
| Bothriochloa ischaemum | Poaceae | 31 | C14–C35 | ++ | N/A | N/A | |||||
| Caryota mitis | Arecaceae | 31 | C20–C35 | + | N/A | N/A | |||||
| Cinnamomum burmannii | Lauraceae | 31 | C22–C35 | + | N/A | ||||||
| Codiaeum variegatum | Euphorbiaceae | 33 | C22–C35 | ++ | N/A | ||||||
| Euphorbia trigona | 33 | C24–C37 | ++ | N/A | |||||||
| Holmskioldia sanguinea | Lamiaceae | 35 | C22–C37 | ++++ | N/A | ||||||
| Hylocereus undatus | Cactaceae | 33 | C18–C37 | +++ | N/A | ||||||
| Imperata cylindrica | Poaceae | 31 | C14–C35 | +++ | N/A | ||||||
| Kigelia africana | Bignoniaceae | 31 | C24–C35 | +++ | N/A | ||||||
| Opuntia dillenii | Cactaceae | 29 | C23–C36 | ++++ | N/A | N/A | |||||
| Osmanthus fragrans | Oleaceae | 31 | C24–C35 | ++ | N/A | N/A | |||||
| Pistia stratiotes | Araceae | 31 | C24–C37 | + | N/A | Yes [55] | |||||
| Swietenia mahagoni | Meliaceae | 31 | C24–C35 | + | N/A | N/A | |||||
| Zoysia japonica | Poaceae | 33 | C14–C35 | +++ | N/A | N/A | |||||
| Aspidosperma spp. | Apocynaceae | Leaves | Alkanes, alkanols | Alkanes | 33 | C29–C35 | +++ | N/A | Tropical | N/A | [66] |
| Cryptomeria japonica | Cypressaceae | fatty acids | 33 | C33–C35 | ++++ | N/A | Temperate | ||||
| Juniperus osteosperma | 33 | C29–C35 | +++ | N/A | Temperate | ||||||
| Manilkara spp. | Sapotaceae | 33 | C31–C35 | +++ | N/A | Tropical | |||||
| Arabidopsis thaliana | Brassicaceae | Leaves 1 | Complete wax profile | Alkanes | 29 | C25–C35 | + | N/A | Temperate | Yes | [21] |
| Miscanthus sinensis | Poaceae | Leaves | Alkanes, fatty acids | Alkanes | 31 | C25–C37 | + | + | Temperate | Yes [52] | [67] |
| Lupinus angustifolius | Fabaceae | Leaves | Complete wax profile | Alkanes | N/A | C23–C37 | N/A | N/A | Temperate | N/A | [68] |
| Triticum aestivum | Poaceae | 31 | N/A | ||||||||
| Olea europaea | Oleaceae | Leaves 1 | Alkanes only | Alkanes | 29 | C27–C35 | + | N/A | Mediterranian | N/A | [69] |
| Olive oil | 25 | C21–C35 | + | N/A | N/A | ||||||
| Arabidopsis thaliana | Brassicaceae | Young/mature | Complete wax profile | Alkanes | 31 | C27–C37 | + | + | Temperate | N/A | [2,7] |
| leaves | Alkenes | 35 | C35–C37 | + | + | ||||||
| Arabidopsis thaliana | Brassicaceae | Leaves 1 | Complete wax profile | Alkanes | 31 | C27–C37 | + | + | Temperate | N/A | [6] |
| Alkenes | 35 | C35–C37 | + | + | |||||||
| Leaf trichomes 1 | Alkanes | 31/33 | C27–C37 | + | + | ||||||
| Alkenes | 35 | C33–C37 | + | + | |||||||
| Ludwigia octovalvis | Onagraceae | Young leaves | Alkanes, fatty acids | Alkanes | 23 | C15–C35 | + | N/A | Tropical | Yes [56] | [70] |
| Mature leaves | 23 | + | N/A |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hegebarth, D.; Jetter, R. Cuticular Waxes of Arabidopsis thaliana Shoots: Cell-Type-Specific Composition and Biosynthesis. Plants 2017, 6, 27. https://doi.org/10.3390/plants6030027
Hegebarth D, Jetter R. Cuticular Waxes of Arabidopsis thaliana Shoots: Cell-Type-Specific Composition and Biosynthesis. Plants. 2017; 6(3):27. https://doi.org/10.3390/plants6030027
Chicago/Turabian StyleHegebarth, Daniela, and Reinhard Jetter. 2017. "Cuticular Waxes of Arabidopsis thaliana Shoots: Cell-Type-Specific Composition and Biosynthesis" Plants 6, no. 3: 27. https://doi.org/10.3390/plants6030027
APA StyleHegebarth, D., & Jetter, R. (2017). Cuticular Waxes of Arabidopsis thaliana Shoots: Cell-Type-Specific Composition and Biosynthesis. Plants, 6(3), 27. https://doi.org/10.3390/plants6030027