


Phytoplankton Communities in the Eastern Tropical Pacific Ocean off Mexico and the Southern Gulf of California During the Strong El Niño of 2023/24

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Oceanic Niño Index
2.2. Vertical Distribution of Conservative Temperature and Chlorophyll-a Levels Along Transect A-A′ from In Situ Data
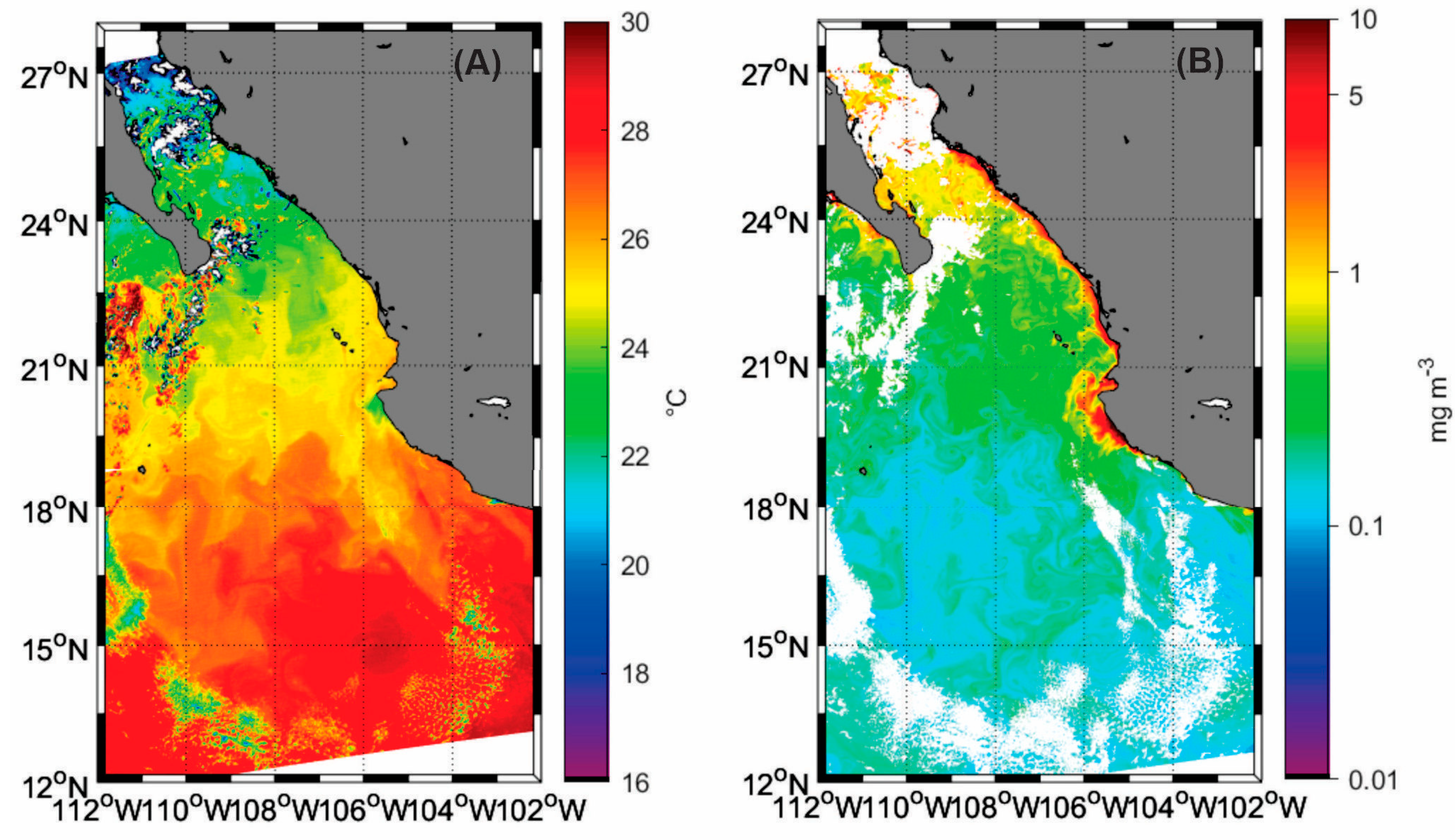
2.3. Sea Surface Temperature and Chlorophyll-a Derived from Satellite
2.4. Phytoplankton Community Structure
2.5. Phytoplankton Horizontal Distribution
3. Discussion
4. Materials and Methods
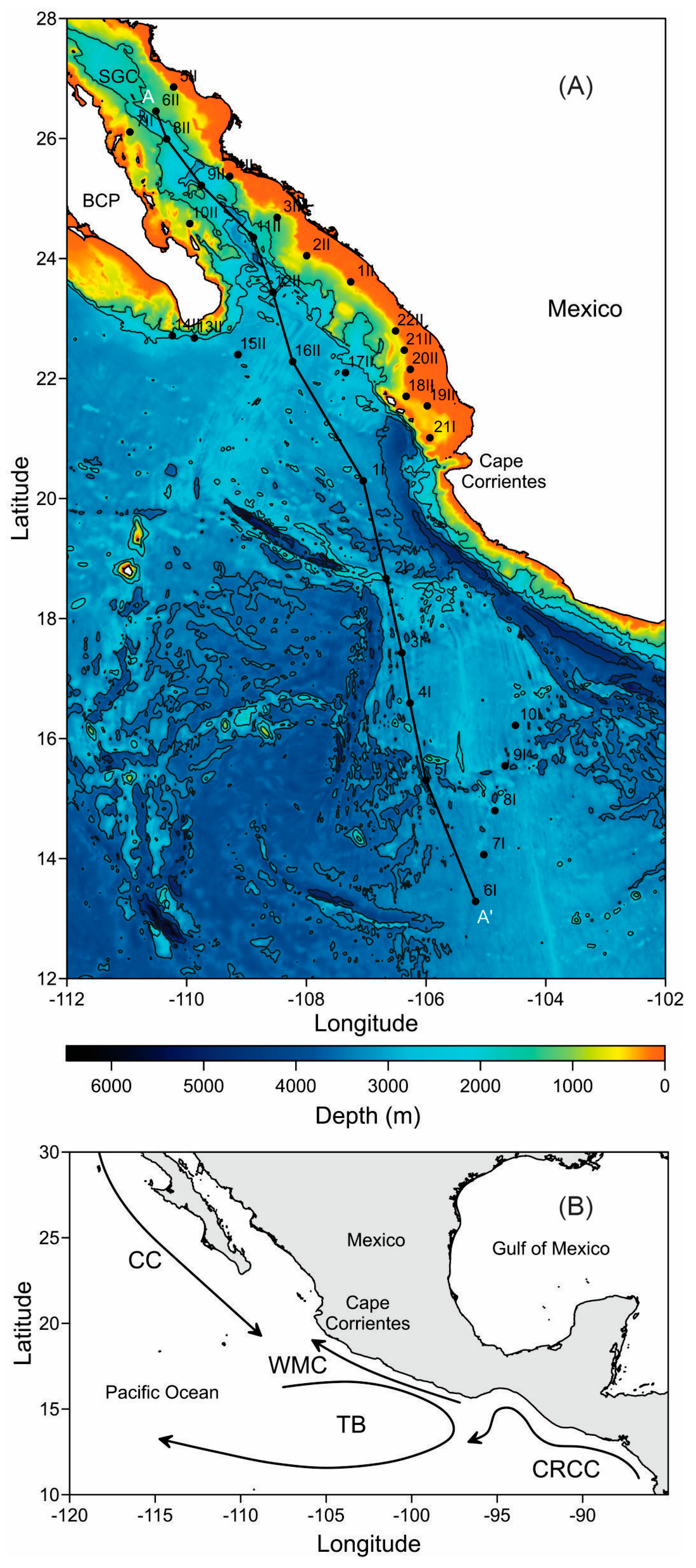
4.1. Study Area
4.2. Sampling
4.3. Laboratory Analysis
4.4. Data Reduction
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jin, F.-F. Toward Understanding El Niño Southern-Oscillation’s Spatiotemporal Pattern Diversity. Front. Earth Sci. 2022, 10, 899139. [Google Scholar] [CrossRef]
- de Moraes, D.F.; da Rocha Marcolin, C.; Lins-Silva, N.; Brito-Lolaia, M.; Varona, H.L.; de Albuquerque Lira, S.M.; Oliveira de Almeida, A.; Schwamborn, R. Declines in coastal macrozooplankton communities within a recent climate and ocean tipping point in the Tropical South Atlantic. Sci. Total Environ. 2024, 954, 176722. [Google Scholar] [CrossRef]
- Castrejón, M.; Pittman, J.; Ramírez-González, J.; Defeo, O. An overview of social-ecological impacts of the El Niño-Southern Oscillation and climate change on Galapagos small-scale fisheries. Ocean Coast. Manag. 2024, 259, 107436. [Google Scholar] [CrossRef]
- Bastianin, A.; Lanza, A.; Manera, M. Economic impacts of El Niño southern oscillation: Evidence from the Colombian coffee market. Agric. Econ. 2018, 49, 623–633. [Google Scholar] [CrossRef]
- Xiong, L.; Gong, K.; Tang, Q.; Dong, Y.; Xu, W. The Impact of the El Niño Southern Oscillation on the Number of Visitors to Natural Attractions: The Moderating Effect of Disposable Personal Income, per Capita GDP and Population. Atmosphere 2021, 12, 1189. [Google Scholar] [CrossRef]
- Gasparin, F.; Roemmich, D. The strong freshwater anomaly during the onset of the 2015/2016 El Niño. Geophys. Res. Lett. 2016, 43, 6452–6460. [Google Scholar] [CrossRef]
- Sun, Y.; Farnsworth, A.; Joachimski, M.M.; Wignall, P.B.; Krystyn, L.; Bond, D.P.G.; Ravidà, D.C.G.; Valdes, P.J. Mega El Niño instigated the end-Permian mass extinction. Science 2024, 385, 6714. [Google Scholar] [CrossRef] [PubMed]
- McPhaden, M.J.; Santoso, S.; Cai, W. Introduction to El Niño Southern Oscillation in a Changing Climate. In El Niño Southern Oscillation in a Changing Climate; Geophysical Monograph Series; McPhaden, M.J., Santoso, A., Cai, W., Eds.; American Geophysical Union: Washington, DC, USA, 2021; pp. 1–19. [Google Scholar]
- Conde, A.; Hurtado, M.; Prado, M. Phytoplankton response to a weak El Niño event. Ecol. Indic. 2018, 95, 394–404. [Google Scholar] [CrossRef]
- Basu, S.; Mackey, K.R.M. Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate. Sustainability 2018, 10, 869. [Google Scholar] [CrossRef]
- Gittings, J.A.; Raitsos, D.E.; Brewin, R.J.W.; Hoteit, I. Links between Phenology of Large Phytoplankton and Fisheries in the Northern and Central Red Sea. Remote Sens. 2021, 13, 231. [Google Scholar] [CrossRef]
- Arteaga, L.A.; Rousseaux, C.S. Impact of Pacific Ocean heatwaves on phytoplankton community composition. Commun. Biol. 2023, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.A.; Nemcek, N.; Robert, M. Phytoplankton responses to the 2014–2016 warming anomaly in the northeast subarctic Pacific Ocean. Limnol. Oceanogr. 2018, 64, 515–525. [Google Scholar] [CrossRef]
- Jiménez-Quiroz, M.; Cervantes-Duarte, R.; Funes-Rodríguez, R.; Barón-Campis, S.A.; García-Romero, F.; Hernández-Trujillo, S.; Hernández-Becerril, D.U.; González-Armas, R.; Martell-Dubois, R.; Cerdeira-Estrada, S.; et al. Impact of “The Blob” and “El Niño” in the SW Baja California Peninsula: Plankton and environmental variability of Bahia Magdalena. Front. Mar. Sci. 2019, 6, 25. [Google Scholar] [CrossRef]
- Blanchot, J.; Rodier, M.; Le Bouteiller, A. Effect of El Nino Southern Oscillation events on the distribution and abundance of phytoplankton in the Western Pacific Tropical Ocean along 165°E. J. Plank. Res. 1992, 14, 137–156. [Google Scholar] [CrossRef]
- Durán-Campos, E.; Salas-de-León, D.A.; Coria-Monter, E.; Monreal-Gómez, M.A.; Quiroz-Martínez, B. Phytoplankton Structure in a Coastal Region of the Eastern Entrance of the Gulf of California during La Niña 2022. Oceans 2024, 5, 647–661. [Google Scholar] [CrossRef]
- Durán-Campos, E.; Torres-Martínez, C.M.; Coria-Monter, E.; Salas-de-León, D.A.; Monreal-Gómez, M.A. Phytoplankton community structure at the eastern entrance of the Gulf of California during El Niño 2023. Lat. Am. J. Aquat. Res. 2024, 52, 659–668. [Google Scholar] [CrossRef]
- Kessler, W.S. The circulation of the eastern tropical Pacific: A review. Progr. Oceanogr. 2006, 69, 181–217. [Google Scholar] [CrossRef]
- Yang, G.; Liu, L.; Zhao, X.; Li, Y.; Duan, Y.; Liu, B.; Huang, K.; Yu, W. Impacts of Different Types of ENSO Events on Thermocline Variability in the Southern Tropical Indian Ocean. Geophys. Res. Lett. 2019, 46, 6775–6785. [Google Scholar] [CrossRef]
- Pan, Y.; Parsons, M.L.; Busman, M.; Moeller, P.D.R.; Dortch, Q.; Powell, C.L.; Doucette, G.L. Pseudo-nitzschia sp. cf. pseudodelicatissima a confirmed producer of domoic acid from the northern Gulf of Mexico. Mar. Ecol. Progr. Ser. 2001, 220, 83–92. [Google Scholar]
- Galeano-Chavarría, A.M.; Arteaga-Sogamoso, E. Distribución y abundancia de diatomeas del género Pseudo-Nitzschia en condición de El Niño 2007 sobre la cuenca Pacífica Colombiana. Bol. Investig. Mar. Costeras 2010, 39, 25–39. [Google Scholar] [CrossRef]
- Horner, R.A.; Hickey, B.M.; Postel, J.R. Pseudo-nitzschia blooms and physical oceanography off Washington State, USA. S. Afr. J. Mar. Sci. 2000, 22, 299–308. [Google Scholar] [CrossRef]
- Zhu, A.; Qu, P.; Fu, F.; Tennenbaum, N.; Tatters, A.O.; Hutchins, D.A. Understanding the blob bloom: Warming increases toxicity and abundance of the harmful bloom diatom Pseudo-nitzschia in California coastal waters. Harmful Algae 2017, 67, 36–43. [Google Scholar] [CrossRef]
- Meave-del-Castillo, M.A.; Zamudio-Resendiz, M.E.; Castillo-Rivera, M. Riqueza fitoplanctónica de la Bahía de Acapulco y zona costera aledaña, Guerrero, México. Acta Bot. Mex. 2012, 100, 405–487. [Google Scholar] [CrossRef]
- Verdugo-Díaz, G.; Martínez-López, A.; Gárate- Lizarraga, I. Indicadores ecológicos de la estructura de la comunidad fitoplanctónica en Bahía Concepción, Golfo de California, México. CICIMAR Oceánides 2010, 25, 95–102. [Google Scholar] [CrossRef]
- Avaria, S.; Muñoz, P. Effects of the 1982–1983 El Niño on the marine phytoplankton off northern Chile. J. Geophys. Res. 1987, 92, 14363–14382. [Google Scholar] [CrossRef]
- Torres, G.; Carnicer, O.; Canepa, A.; De La Fuente, P.; Recalde, S.; Narea, R.; Pinto, E.; Borbor-Córdova, M.J. Spatio-temporal pattern of dinoflagellates along the Tropical Eastern Pacific coast (Ecuador). Front. Mar. Sci. 2019, 6, 145. [Google Scholar] [CrossRef]
- Yin, K.; Harrison, P.J.; Chen, J.; Huang, W.; Qian, P.-Y. Red tides during spring 1998 in Hong Kong: Is El Niño responsible? Mar. Ecol. Progr. Ser. 1999, 187, 289–294. [Google Scholar] [CrossRef]
- Sherr, E.B.; Sherr, B.F. Heterotrophic dinoflagellates: A significant component of microzooplankton biomass and major grazers of diatoms in the sea. Mar. Ecol. Progr. Ser. 2007, 352, 187–197. [Google Scholar] [CrossRef]
- Bonnet, S.; Guieu, C.; Chiaverini, J.; Ras, J.; Stock, A. Effect of atmospheric nutrients on the autotrophic communities in a low nutrient, low chlorophyll system. Limnol. Oceanogr. 2005, 50, 1810–1819. [Google Scholar] [CrossRef]
- Duarte, C.M.; Regaudie-de-Gioux, A.; Arrieta, J.M.; Delgado-Huertas, A.; Agustí, S. The Oligotrophic Ocean Is Heterotrophic. Annu. Rev. Mar. Sci. 2013, 5, 551–569. [Google Scholar] [CrossRef]
- Hernández-Becerril, D.U.; Bravo-Sierra, E. Planktonic Silicoflagellates (Dictyochophyceae) from the Mexican Pacific Ocean. Bot. Mar. 2001, 44, 417–423. [Google Scholar] [CrossRef]
- Pérez-Cruz, L.; Molina-Cruz, A. El Niño 1983: Effect on the distribution of the Silicoflagellates in the Gulf of California. Cienc. Mar. 1988, 14, 9–38. [Google Scholar] [CrossRef]
- Ochoa, N.; Gómez, O. Dinoflagellates as indicators of water masses during El Niño, 1982–1983. J. Geophys. Res. Ocean. 1987, 92, 14355–14367. [Google Scholar] [CrossRef]
- Matrai, P.A. The distribution of the dinoflagellate Ceratium in relation to environmental factors along 28°N in the eastern North Pacific. J. Plank. Res. 1986, 8, 105–118. [Google Scholar] [CrossRef]
- Castillo, F.A.; Vizcaino Bravo, Z. Observación del fitoplancton del Pacífico Colombiano durante 1991–1992 en condiciones El Niño. Bull. Inst. Fr. Études Andin. 1993, 22, 179–190. [Google Scholar] [CrossRef]
- Gárate-Lizárraga, I. Primer registro de Ceratium dens (Dinophyceae) en el Golfo de California. CICIMAR Ocean. 2009, 24, 167–173. [Google Scholar] [CrossRef]
- Lavaniegos-Espejo, B.E.; Lara-Lara, J.R. Zooplankton of the Gulf of California after the 1982-1983 EI Niño Event: Biomass Distribution and Abundance. Pac. Sci. 1990, 44, 297–310. [Google Scholar]
- Santamaría-del-Angel, E.; Álvarez-Borrego, S.; Müller-Karger, F.E. The 1982–1984 El Niño in the Gulf of California as seen in coastal zone color scanner imagery. J. Geophys. Res. Ocean. 1994, 99, 7423–7431. [Google Scholar] [CrossRef]
- Sánchez-Velasco, L.; Beier, E.; Godínez, V.M.; Barton, E.D.; Santamaría-del-Angel, E.; Jiménez-Rosemberg, S.P.A.; Marinone, S.G. Hydrographic and fish larvae distribution during the “Godzilla El Niño 2015-2016” in the northern end of the shallow oxygen minimum zone of the Eastern Tropical Pacific Ocean. J. Geophys. Res. Ocean. 2017, 122, 2156–2170. [Google Scholar] [CrossRef]
- Coria-Monter, E.; Monreal-Gómez, M.A.; Salas de León, D.A.; Durán-Campos, E. Impact of the “Godzilla El Niño” event of 2015-2016 on sea surface temperature and chlorophyll-a in the southern Gulf of California, Mexico, as evidenced by satellite and in situ data. Pac. Sci. 2018, 72, 411–422. [Google Scholar] [CrossRef]
- Durán-Campos, E.; Coria-Monter, E.; Monreal-Gómez, M.A.; Salas-de-León, D.A. Impact of “the Blob” 2014 and 2019 in the sea surface temperature and chlorophyll-a levels of the Gulf of California: A satellite-based study. Lat. Am. J. Aquat. Res. 2022, 50, 479–491. [Google Scholar] [CrossRef]
- Coria-Monter, E.; Salas de León, D.A.; Monreal-Gómez, M.A.; Durán-Campos, E. Satellite observations of the effect of the “Godzilla El Niño” on the Tehuantepec upwelling system in the Mexican Pacific. Helgol. Mar. Res. 2019, 73, 3. [Google Scholar] [CrossRef]
- Durán-Campos, E.; Salas-de-Léon, D.A.; Coria-Monter, E.; Monreal-Gómez, M.A.; Aldeco-Ramírez, J.; Quiroz-Martínez, B. ENSO effects in the southern gulf of California estimated from satellite data. Cont. Shelf Res. 2023, 266, 105084. [Google Scholar] [CrossRef]
- Fernández-Alamo, M.A.; Farber-Lorda, J. Zooplankton and the oceanography of the Eastern Tropical Pacific: A review. Progr. Oceanogr. 2006, 69, 318–359. [Google Scholar] [CrossRef]
- Álvarez-Borrego, S. Phytoplankton biomass and production in the Gulf of California: A review. Bot. Mar. 2012, 55, 119–128. [Google Scholar] [CrossRef]
- Quiróz-Martínez, B.; Salas-de-León, D.A.; Gil-Zurita, A.; Monreal-Gómez, M.A.; Coria-Monter, E.; Durán-Campos, E. Latitudinal and archipelago effect on the composition, distribution, and abundance of zooplanktonic organisms in the Gulf of California. Oceanologia 2023, 65, 371–385. [Google Scholar] [CrossRef]
- Edler, L.; Elbrachter, M. The Utermöhl Method for Quantitative Phytoplankton Analysis. In Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis; Karlson, B., Cusack, C., Bresnan, E., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Potsdam, Germany, 2010; pp. 13–20. [Google Scholar]
- Tomas, C.R. Identifying Marine Phytoplankton; Academic Press: San Diego, CA, USA, 1997; p. 858. [Google Scholar]
- Throndsen, J.; Hasle, G.R.; Tangen, K. Norsk Kystplankton Flora; Almater Forlag As: Oslo, Norway, 2003; p. 341. [Google Scholar]
- Licea, S.; Moreno, J.L.; Santoyo, H.; Figueroa, G. Dinoflageladas del Golfo de California; Universidad Autónoma de Baja California Sur, SEP-FOMES, PROMARCO: Baja California, Mexico, 1995; p. 165. [Google Scholar]
- Moreno, J.L.; Licea, S.; Santoyo, H. Diatomeas del Golfo de California; Universidad Autónoma de Baja California Sur, SEP-FOMES, PROMARCO: Baja California, Mexico, 1996; p. 273. [Google Scholar]
- Esqueda-Lara, L.; Hernández-Becerril, D.U. Dinoflagelados Microplanctónicos Marinos del Pacífico Central de México (Isla Isabel, Nayarit y Costas de Jalisco y Colima); Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México: Ciudad de México, Mexico, 2010; p. 206. [Google Scholar]
- Morquecho-Escamilla, L.; Reyes-Salinas, A.; Okolodkov, Y. Illustrated Taxonomic Guide of the Marine Dinoflagellate Collection (CODIMAR); Centro de Investigaciones Biológicas del Noroeste, S.C. La Paz: Baja California Sur, Mexico, 2016; p. 147. [Google Scholar]
- Hernández-Becerril, D.U.; Barón-Campis, S.A.; Ceballos-Corona, J.G.A.; Alonso-Rodríguez, R.; Rincones-Reyes, K.M.; Becerra-Reynoso, R.T.; Arce-Rocha, G. Catálogo de Fitoplancton del Pacífico Central Mexicano, Cruceros “MareaR” (2009–2019) B/O “El Puma”; Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México: Ciudad de México, Mexico, 2021; p. 254. [Google Scholar]
- Glantz, M.H.; Ramirez, I.J. Reviewing the Oceanic Niño Index (ONI) to Enhance Societal Readiness for El Niño’s Impacts. Int. J. Disaster Risk. Sci. 2020, 11, 394–403. [Google Scholar] [CrossRef]
- Lavín, M.F.; Castro, R.; Beier, E.; Godínez, V.M.; Amador, A.; Guest, P. SST, thermohaline structure, and circulation in the southern Gulf of California in June 2004 during the North American Monsoon Experiment. J. Geophys. Res. 2009, 114, C02025. [Google Scholar] [CrossRef]
- ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination; Version 5.10; Wageningen University & Research: Wageningen, The Netherlands, 2018; p. 536. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monreal-Gómez, M.A.; Pérez-Cruz, L.; Durán-Campos, E.; Salas-de-León, D.A.; Torres-Martínez, C.M.; Coria-Monter, E. Phytoplankton Communities in the Eastern Tropical Pacific Ocean off Mexico and the Southern Gulf of California During the Strong El Niño of 2023/24. Plants 2025, 14, 1375. https://doi.org/10.3390/plants14091375
Monreal-Gómez MA, Pérez-Cruz L, Durán-Campos E, Salas-de-León DA, Torres-Martínez CM, Coria-Monter E. Phytoplankton Communities in the Eastern Tropical Pacific Ocean off Mexico and the Southern Gulf of California During the Strong El Niño of 2023/24. Plants. 2025; 14(9):1375. https://doi.org/10.3390/plants14091375
Chicago/Turabian StyleMonreal-Gómez, María Adela, Ligia Pérez-Cruz, Elizabeth Durán-Campos, David Alberto Salas-de-León, Carlos Mauricio Torres-Martínez, and Erik Coria-Monter. 2025. "Phytoplankton Communities in the Eastern Tropical Pacific Ocean off Mexico and the Southern Gulf of California During the Strong El Niño of 2023/24" Plants 14, no. 9: 1375. https://doi.org/10.3390/plants14091375
APA StyleMonreal-Gómez, M. A., Pérez-Cruz, L., Durán-Campos, E., Salas-de-León, D. A., Torres-Martínez, C. M., & Coria-Monter, E. (2025). Phytoplankton Communities in the Eastern Tropical Pacific Ocean off Mexico and the Southern Gulf of California During the Strong El Niño of 2023/24. Plants, 14(9), 1375. https://doi.org/10.3390/plants14091375