
Physiological Adaptations of Vigna radiata to Heavy Metal Stress: Soluble Sugar Accumulation and Biomass Enhancement


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
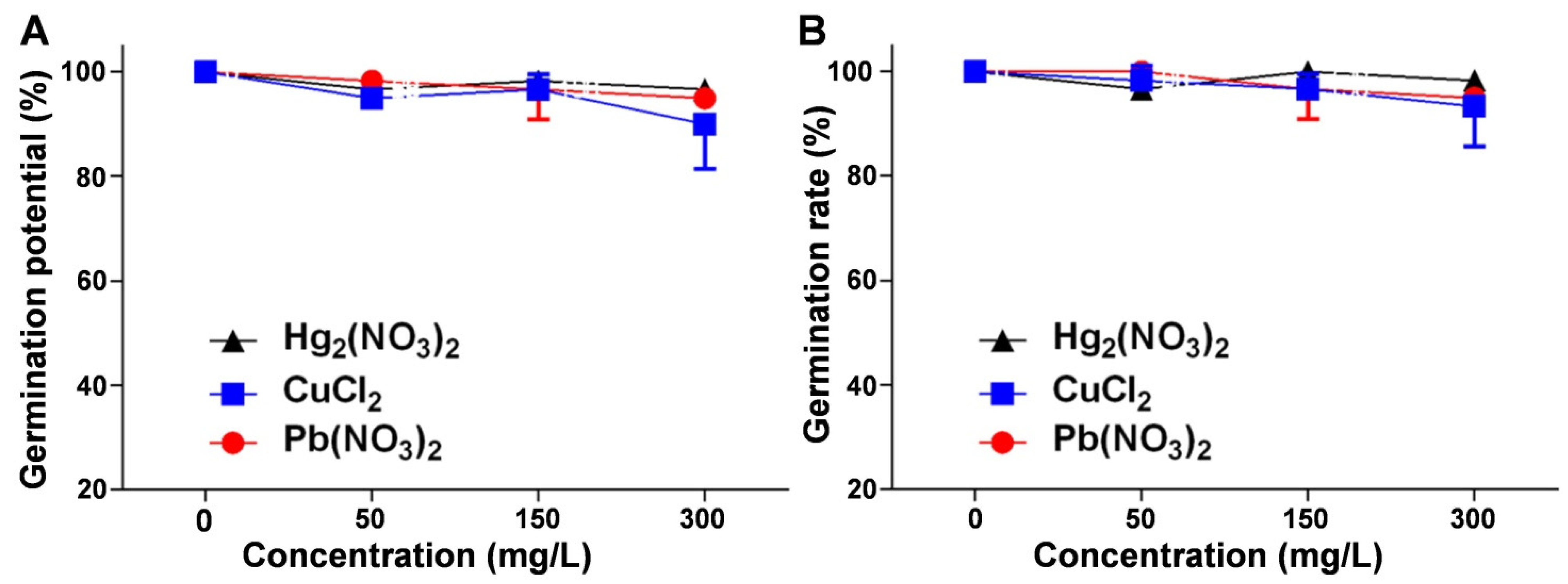
2.1. Analysis of the Effects of Heavy Metal Ion Stress on V. radiata Germination
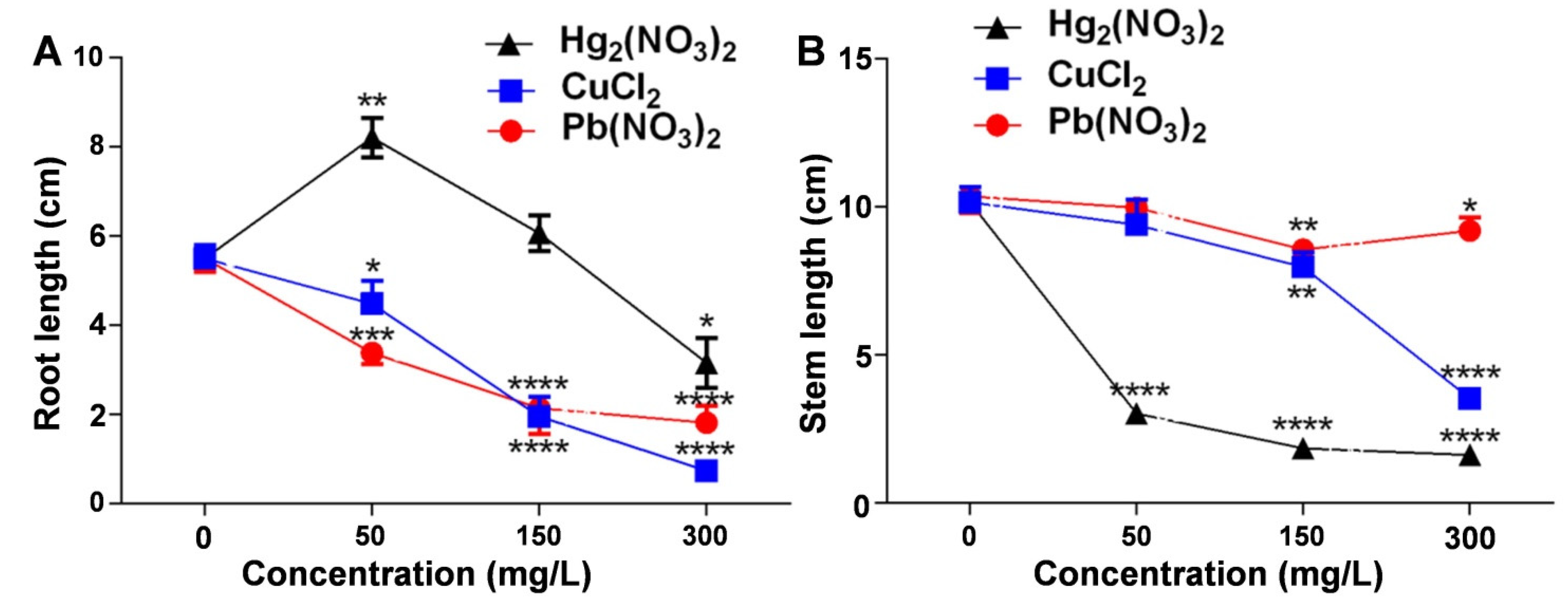
2.2. Comparative Study of the Effects of Different Heavy Metal Ion Concentrations on the Root and Shoot Growth of V. radiata Seedlings
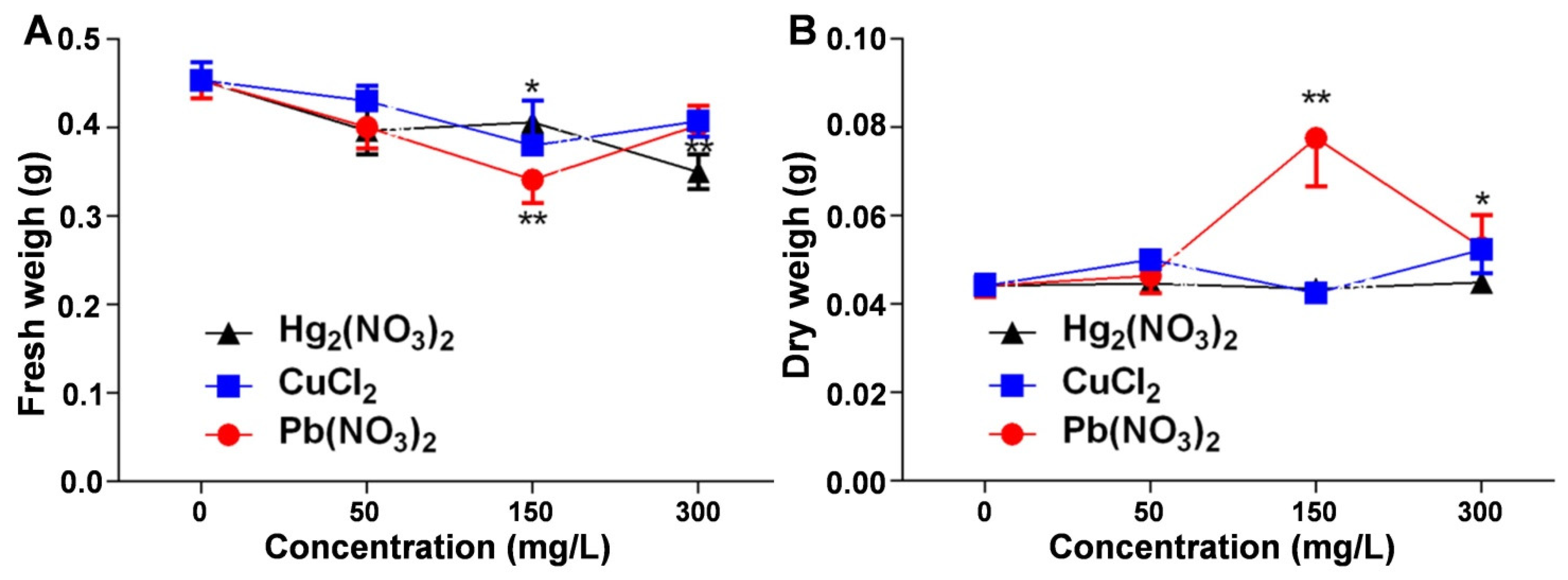
2.3. Analysis of the Effects of Heavy Metal Ion Stress on the Fresh and Dry Weight of V. radiata Seedlings
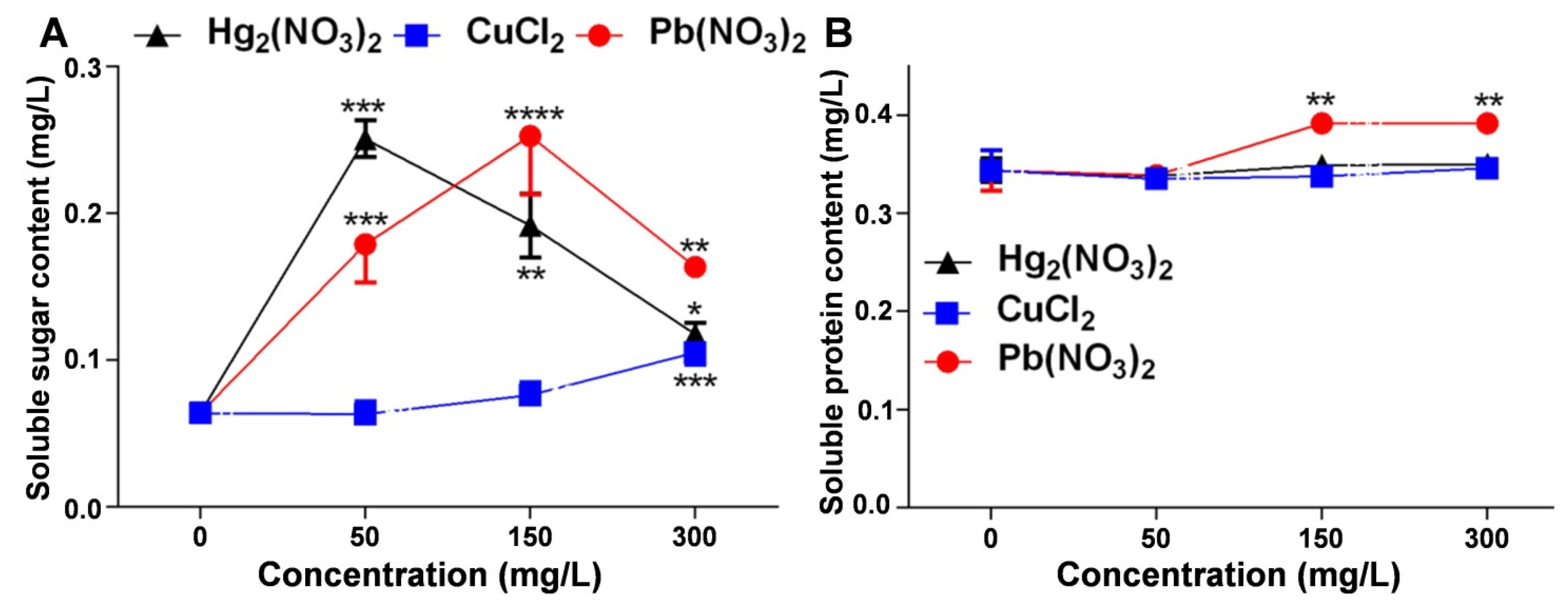
2.4. Regulatory Effects of Heavy Metal Ion Stress on Soluble Sugar and Protein Content in V. radiata Seedlings
2.5. PCA Reveals Dual Impacts of Heavy Metal Stress on Plant Morphology and Metabolism
3. Materials and Methods
3.1. Materials and Equipment
3.2. Methodology for Assessing V. radiata Responses to Different Heavy Metal Stress Concentrations
3.3. Determination of Germination Potential and Germination Rate
3.4. Determination of Soluble Sugar
3.5. Determination of Soluble Protein
3.6. Principal Component Analysis (PCA) and Statistical Analysis
3.7. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| PCA | Principal component analysis |
References
- Maqsood, Q.; Hussain, N.; Mumtaz, M.; Bilal, M.; Iqbal, H.M.N. Novel strategies and advancement in reducing heavy metals from the contaminated environment. Arch. Microbiol. 2022, 204, 478. [Google Scholar] [CrossRef] [PubMed]
- Idowu, G.A. Heavy metals research in Nigeria: A review of studies and prioritization of research needs. Environ. Sci. Pollut. Res. Int. 2022, 29, 65940–65961. [Google Scholar] [CrossRef] [PubMed]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Li, N.; Wang, H.; Su, W.; Song, Q.; Liang, Q.; Liang, M.; Sun, C.; Li, Y.; Lowe, S.; et al. Combined exposure to multiple metals on cardiovascular disease in NHANES under five statistical models. Environ. Res. 2022, 215, 114435. [Google Scholar] [CrossRef]
- Martins, A.C.; Ferrer, B.; Tinkov, A.A.; Caito, S.; Deza-Ponzio, R.; Skalny, A.V.; Bowman, A.B.; Aschner, M. Association between Heavy Metals, Metalloids and Metabolic Syndrome: New Insights and Approaches. Toxics 2023, 11, 670. [Google Scholar] [CrossRef]
- Chaplygin, V.A.; Burachevskay, M.V.; Minkina, T.M.; Mandzhieva, S.S.; Siromlya, T.I.; Chernikova, N.P.; Dudnikova, T.S. Accumulation and Distribution of Heavy Metals in Soils and Medicinal Plants in the Impact Zone of Novocherkassk Power Station. Eurasian Soil Sci. 2024, 57, 1746–1758. [Google Scholar] [CrossRef]
- Terron-Camero, L.C.; Pelaez-Vico, M.A.; Del-Val, C.; Sandalio, L.M.; Romero-Puertas, M.C. Role of nitric oxide in plant responses to heavy metal stress: Exogenous application versus endogenous production. J. Exp. Bot. 2019, 70, 4477–4488. [Google Scholar] [CrossRef]
- Mohammadi, A.; Mansour, S.N.; Najafi, M.L.; Toolabi, A.; Abdolahnejad, A.; Faraji, M.; Miri, M. Probabilistic risk assessment of soil contamination related to agricultural and industrial activities. Environ. Res. 2022, 203, 111837. [Google Scholar] [CrossRef]
- Yang, J.; Sun, Y.; Wang, Z.; Gong, J.; Gao, J.; Tang, S.; Ma, S.; Duan, Z. Heavy metal pollution in agricultural soils of a typical volcanic area: Risk assessment and source appointment. Chemosphere 2022, 304, 135340. [Google Scholar] [CrossRef]
- Azarin, K.; Usatov, A.; Minkina, T.; Duplii, N.; Fedorenko, A.; Plotnikov, A.; Mandzhieva, S.; Kumar, R.; Yong, J.W.H.; Sehar, S. Evaluating the phytotoxicological effects of bulk and nano forms of zinc oxide on cellular respiration-related indices and differential gene expression in Hordeum vulgare L. Ecotoxicol. Environ. Saf. 2024, 282, 116670. [Google Scholar] [CrossRef]
- Du, Y.; Yu, C.; Sun, Z.; Liu, Y.; Liu, X.; Feng, Y.; Wang, H.; Zhou, J.; Li, X. Soil resource availability regulates the response of micro-food web multitrophic interactions to heavy metal contamination. Environ. Res. 2025, 273, 121222. [Google Scholar] [CrossRef] [PubMed]
- Niekerk, L.A.; Gokul, A.; Basson, G.; Badiwe, M.; Nkomo, M.; Klein, A.; Keyster, M. Heavy metal stress and mitogen activated kinase transcription factors in plants: Exploring heavy metal-ROS influences on plant signalling pathways. Plant Cell Environ. 2024, 47, 2793–2810. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Wang, R.; Sun, X.; Liu, L.; Liu, P.; Tang, J.; Zhang, C.; Liu, H. Heavy metal stress in plants: Ways to alleviate with exogenous substances. Sci. Total Environ. 2023, 897, 165397. [Google Scholar] [CrossRef] [PubMed]
- Nazir, F.; Fariduddin, Q.; Khan, T.A. Hydrogen peroxide as a signalling molecule in plants and its crosstalk with other plant growth regulators under heavy metal stress. Chemosphere 2020, 252, 126486. [Google Scholar] [CrossRef]
- Zhou, Y.; Fu, J.; Ye, Y.; Xu, Q.; Liang, J.; Chen, Y.; Mo, Y.; Liu, K. Physiological and molecular response mechanisms of tomato seedlings to cadmium (Cd) and lead (Pb) stress. PeerJ 2024, 12, e18533. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Sun, H.; Li, Y.; Ma, L.; Li, Y.; Wang, L.; Zhang, L.; Li, X. Integrated transcriptomics and metabolomics reveal the role of soluble sugars and GABA in rice leaf response to Pb stress. Plant Physiol. Biochem. 2025, 221, 109595. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, M.; Wu, R.; Chen, X.; Liu, F.; Xing, B. Genome-wide analysis of OSCA gene family members in Vigna radiata and their involvement in the osmotic response. BMC Plant Biol. 2021, 21, 408. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Lone, J.K.; Pandey, R.; Mondal, N.; Dhandapani, R.; Meena, S.K.; Khan, S.; Gayacharan. Insights into morphological and physio-biochemical adaptive responses in mungbean (Vigna radiata L.) under heat stress. Front. Genet. 2023, 14, 1206451. [Google Scholar] [CrossRef]
- Seneviratne, M.; Weerasundara, L.; Ok, Y.S.; Rinklebe, J.; Vithanage, M. Phytotoxicity attenuation in Vigna radiata under heavy metal stress at the presence of biochar and N fixing bacteria. J. Environ. Manag. 2017, 186, 293–300. [Google Scholar] [CrossRef]
- Chau, T.P.; Devanesan, S.; Farhat, K.; Liu, X.; Jhanani, G.K. Phytoremediation efficiency of Vigna mungo with the amalgamation of indigenous metal tolerant bacterial strain on metal polluted agriculture soil. Environ. Res. 2023, 232, 116291. [Google Scholar] [CrossRef]
- Kumar, N.; Sharma, V.; Kaur, G.; Lata, C.; Dasila, H.; Perveen, K.; Khan, F.; Gupta, V.K.; Khanam, M.N. Brassinosteroids as promoters of seedling growth and antioxidant activity under heavy metal zinc stress in mung bean (Vigna radiata L.). Front. Microbiol. 2023, 14, 1259103. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Chattha, M.U.; Khan, I.; Mahmood, A.; Hassan, M.U.; Al-Huqail, A.A.; Salem, M.Z.M.; Ali, H.M.; Hano, C.; El-Esawi, M.A. Exogenously Applied Trehalose Augments Cadmium Stress Tolerance and Yield of Mung Bean (Vigna radiata L.) Grown in Soil and Hydroponic Systems through Reducing Cd Uptake and Enhancing Photosynthetic Efficiency and Antioxidant Defense Systems. Plants 2022, 11, 822. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Sharma, N.L.; Singh, C.K.; Yerramilli, V.; Narayan, R.; Sarkar, S.K.; Singh, I. Chromium (VI)-Induced Alterations in Physio-Chemical Parameters, Yield, and Yield Characteristics in Two Cultivars of Mungbean (Vigna radiata L.). Front. Plant Sci. 2021, 12, 735129. [Google Scholar] [CrossRef]
- Chen, F.; Aqeel, M.; Maqsood, M.F.; Khalid, N.; Irshad, M.K.; Ibrahim, M.; Akhter, N.; Afzaal, M.; Ma, J.; Hashem, M.; et al. Mitigation of lead toxicity in Vigna radiata genotypes by silver nanoparticles. Environ. Pollut. 2022, 308, 119606. [Google Scholar] [CrossRef] [PubMed]
- Mendes, N.A.C.; Cunha, M.L.O.; Bosse, M.A.; Silva, V.M.; Moro, A.L.; Agathokleous, E.; Vicente, E.F.; Reis, A.R.D. Physiological and biochemical role of nickel in nodulation and biological nitrogen fixation in Vigna unguiculata L. Walp. Plant Physiol. Biochem. 2023, 201, 107869. [Google Scholar] [CrossRef]
- Karmous, I.; Tlahig, S.; Loumerem, M.; Lachiheb, B.; Bouhamda, T.; Mabrouk, M.; Debouba, M.; Chaoui, A. Assessment of the risks of copper- and zinc oxide-based nanoparticles used in Vigna radiata L. culture on food quality, human nutrition and health. Environ. Geochem. Health 2022, 44, 4045–4061. [Google Scholar] [CrossRef]
- Leng, Y.; Li, Y.; Wen, Y.; Zhao, H.; Wang, Q.; Li, S.W. Transcriptome analysis provides molecular evidences for growth and adaptation of plant roots in cadimium-contaminated environments. Ecotoxicol. Environ. Saf. 2020, 204, 111098. [Google Scholar] [CrossRef]
- Rizvi, A.; Khan, M.S. Biotoxic impact of heavy metals on growth, oxidative stress and morphological changes in root structure of wheat (Triticum aestivum L.) and stress alleviation by Pseudomonas aeruginosa strain CPSB1. Chemosphere 2017, 185, 942–952. [Google Scholar] [CrossRef]
- Fan, C.; Liang, Q.; Wang, Y.; Chen, P.; Wu, J.; Wu, Q.; Jiang, S.; Zhou, Y.; He, R.; Tai, F. Cu-II-directed self-assembly of fullerenols to ameliorate copper stress in maize seedlings. Sci. Total Environ. 2024, 929, 172416. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Gupta, P.; Singh, A. Conjoint application of novel bacterial isolates on dynamic changes in oxidative stress responses of axenic Brassica juncea L. in Hg-stress soils. J. Hazard. Mater. 2022, 434, 128854. [Google Scholar] [CrossRef]
- Cai, X.; Fu, J.; Li, X.; Peng, L.; Yang, L.; Liang, Y.; Jiang, M.; Ma, J.; Sun, L.; Guo, B.; et al. Low-molecular-weight organic acid-mediated tolerance and Pb accumulation in centipedegrass under Pb stress. Ecotoxicol. Environ. Saf. 2022, 241, 113755. [Google Scholar] [CrossRef] [PubMed]
- Rondel, C.; Marcato-Romain, C.E.; Girbal-Neuhauser, E. Development and validation of a colorimetric assay for simultaneous quantification of neutral and uronic sugars. Water Res. 2013, 47, 2901–2908. [Google Scholar] [CrossRef] [PubMed]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef]
- Malik, A.; Khan, J.M.; Sen, P.; Alamri, A.; Karan, R.; Emerson, I.A. Coomassie Brilliant Blue Induces Coiled-Coil Aggregation in Lysozyme at pH 7.4 by Hydrophobic and Electrostatic Forces. ACS Omega 2024, 10, 1829–1838. [Google Scholar] [CrossRef]
- Anas, M.; Khattak, W.A.; Fahad, S.; Alrawiq, N.; Alrawiq, H.S.; Abdelsalam, N.R.; Jaremko, M.; Quraishi, U.M. Mitigating heavy metal pollution in agriculture: A multi-omics and nanotechnology approach to safeguard global wheat production. J. Hazard. Mater. Adv. 2024, 17, 100584. [Google Scholar] [CrossRef]
- Huang, L.; Li, W.C.; Tam, N.F.Y.; Ye, Z. Effects of root morphology and anatomy on cadmium uptake and translocation in rice (Oryza sativa L.). J. Environ. Sci. 2019, 75, 296–306. [Google Scholar] [CrossRef]
- Song, J.; Sun, Z.; Saud, S.; Fahad, S.; Nawaz, T. Exploring the deleterious effects of heavy metal cadmium on antioxidant defense and photosynthetic pathways in higher plants. Plant Stress 2024, 15, 100716. [Google Scholar] [CrossRef]
- Azarin, K.; Usatov, A.; Minkina, T.; Alliluev, I.; Duplii, N.; Mandzhieva, S.; Singh, A.; Rajput, V.D.; Kumar, S.; Fakhr, M.A. Impact nano-and micro-form of CdO on barley growth and oxidative stress response. J. King Saud Univ. Sci. 2024, 36, 103493. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Zinta, G.; Hamed, B.A.; Selim, S.; Beemster, G.; Hozzein, W.N.; Wadaan, M.A.M.; Asard, H.; Abuelsoud, W. Maize roots and shoots show distinct profiles of oxidative stress and antioxidant defense under heavy metal toxicity. Environ. Pollut. 2020, 258, 113705. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, H.; Yang, Y.; Chen, H.; Chen, C.; Cheng, W. Metabonomics reveals the mechanism of stress resistance in Vetiveria zizanioides inoculated with AMF under copper stress. Sci. Rep. 2025, 15, 6005. [Google Scholar] [CrossRef]
- Mishra, S.; Sharma, A.; Srivastava, A.K. Ascorbic acid: A metabolite switch for designing stress-smart crops. Crit. Rev. Biotechnol. 2024, 44, 1350–1366. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhang, S.; Lu, L.; Liu, L.; Liang, J.; Lang, S.; Wang, C.; Wang, L.; Li, Z. Effects of combined ultrasound and calcium ion pretreatments on polyphenols during mung bean germination: Exploring underlying mechanisms. Food Res. Int. 2024, 195, 114947. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Ma, P.; Huang, L.; Wang, X.; Chen, C.; Liu, C.; Wei, T.; Yang, J.; Guo, J.; Li, J. Hydrogen sulphide regulates the growth of tomato root cells by affecting cell wall biosynthesis under CuO NPs stress. Plant Biol. 2022, 24, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Shar, A.G.; Zhang, L.; Lu, A.; Ahmad, M.; Saqib, M.; Hussain, S.; Zulfiqar, U.; Wang, P.; Zhang, L.; Rahimi, M. Unlocking Biochar’s Potential: Innovative Strategies for Sustainable Remediation of Heavy Metal Stress in Tobacco Plants. Scientifica 2025, 2025, 6302968. [Google Scholar] [CrossRef]
- Huang, Z.; Tang, R.; Yi, X.; Xu, W.; Zhu, P.; Jiang, C.-Z. Overexpressing Phytochrome Interacting Factor 8 of Myrothamnus flabellifolia Enhanced Drought and Salt Tolerance in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 8155. [Google Scholar] [CrossRef]
- Naz, T.; Iqbal, M.M.; Raza, B.; Mubeen, M.A.; Nadeem, M.A.; Al-Ghamdi, A.A.; Elshikh, M.S.; Rizwan, M.; Iqbal, R. Green remediation of lead (pb) from Pb-toxic soil by combined use of silicon nanomaterials and leguminous Lens culinaris L. plants. Sci. Rep. 2025, 15, 4366. [Google Scholar] [CrossRef]
- Anjum, S.; Sarwar, M.; Mukhtar, A.; Hussain, T.; Liaquat, M.; Batool, A.; Alam, M.W.; Ali, Q.; Ashraf, M.I. Impact of rhizobacterial inoculation in plant growth medium to mitigate lead stress in Tomato (Solanum lycopersicum L.). Sci. Rep. 2025, 15, 707. [Google Scholar] [CrossRef]
- Feng, Z.; Ji, S.; Ping, J.; Cui, D. Recent advances in metabolomics for studying heavy metal stress in plants. TrAC Trends Anal. Chem. 2021, 143, 116402. [Google Scholar] [CrossRef]
- Li, J.; Xie, J.; Yu, J.; Lyv, J.; Zhang, J.; Ding, D.; Li, N.; Zhang, J.; Bakpa, E.P.; Yang, Y.; et al. Melatonin enhanced low-temperature combined with low-light tolerance of pepper (Capsicum annuum L.) seedlings by regulating root growth, antioxidant defense system, and osmotic adjustment. Front Plant Sci. 2022, 13, 998293. [Google Scholar] [CrossRef]
- Chauhan, J.; Srivastava, J.P.; Singhal, R.K.; Soufan, W.; Dadarwal, B.K.; Mishra, U.N.; Anuragi, H.; Rahman, M.A.; Sakran, M.I.; Brestic, M.; et al. Alterations of Oxidative Stress Indicators, Antioxidant Enzymes, Soluble Sugars, and Amino Acids in Mustard [Brassica juncea (L.) Czern and Coss.] in Response to Varying Sowing Time, and Field Temperature. Front. Plant Sci. 2022, 13, 875009. [Google Scholar] [CrossRef]
- Huang, L.; Liu, X.; Liu, Y.; Tanveer, M.; Chen, W.; Fu, W.; Wang, Q.; Guo, Y.; Shabala, S. Revealing mechanistic basis of ameliorating detrimental effects of cadmium in cherry tomatoes by exogenous application of melatonin and brassinosteroids. Ecotoxicol. Environ. Saf. 2024, 283, 116768. [Google Scholar] [CrossRef] [PubMed]
- Menhas, S.; Hayat, K.; Lin, D.; Shahid, M.; Bundschuh, J.; Zhu, S.; Hayat, S.; Liu, W. Citric acid-driven cadmium uptake and growth promotion mechanisms in Brassica napus. Chemosphere 2024, 368, 143716. [Google Scholar] [CrossRef] [PubMed]
- Khalofah, A. Integrated roles of nitric oxide and melatonin in enhancing chromium resilience in cotton plants: Modulation of thiol metabolism and antioxidant responses. Environ. Sci. Pollut. Res. Int. 2024, 31, 66463–66476. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, R.; Ahmad, F.; Kaya, C.; Upadhyay, S.K.; Muneer, S.; Kumar, V.; Meena, M.; Liu, H.; Upadhyay, H.; Seth, C.S. Decrypting proteomics, transcriptomics, genomics, and integrated omics for augmenting the abiotic, biotic, and climate change stress resilience in plants. J. Plant Physiol. 2025, 305, 154430. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, W.; Bai, J.; Yu, H.; Han, G. Physiological Adaptations of Vigna radiata to Heavy Metal Stress: Soluble Sugar Accumulation and Biomass Enhancement. Plants 2025, 14, 1191. https://doi.org/10.3390/plants14081191
Qi W, Bai J, Yu H, Han G. Physiological Adaptations of Vigna radiata to Heavy Metal Stress: Soluble Sugar Accumulation and Biomass Enhancement. Plants. 2025; 14(8):1191. https://doi.org/10.3390/plants14081191
Chicago/Turabian StyleQi, Wenjing, Jinping Bai, Han Yu, and Guojun Han. 2025. "Physiological Adaptations of Vigna radiata to Heavy Metal Stress: Soluble Sugar Accumulation and Biomass Enhancement" Plants 14, no. 8: 1191. https://doi.org/10.3390/plants14081191
APA StyleQi, W., Bai, J., Yu, H., & Han, G. (2025). Physiological Adaptations of Vigna radiata to Heavy Metal Stress: Soluble Sugar Accumulation and Biomass Enhancement. Plants, 14(8), 1191. https://doi.org/10.3390/plants14081191