
Light Intensity Dependence of CO2 Assimilation Is More Related to Biochemical Capacity Rather than Diffusional Conductance

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Condition
2.2. Gas Exchange and Chlorophyll Fluorescence Measurements
2.3. Calculation of the Mesophyll Conductance
2.4. Quantitative Calculation of Photosynthetic Limitation
2.5. Calculation of Electron Flow for Photorespiration
2.6. Statistical Analysis
3. Results
3.1. The Steady-State Photosynthesis at Saturating Light
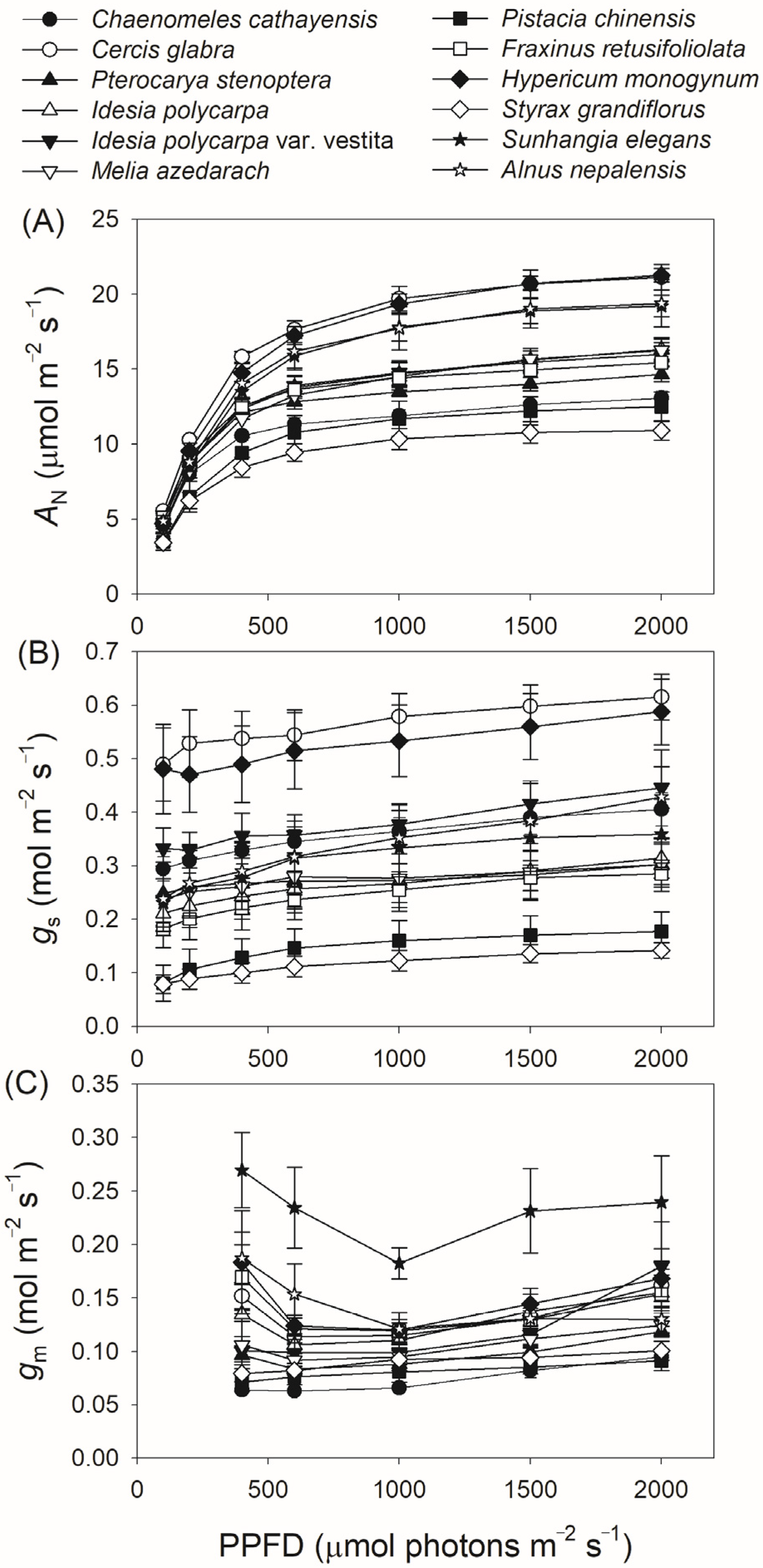
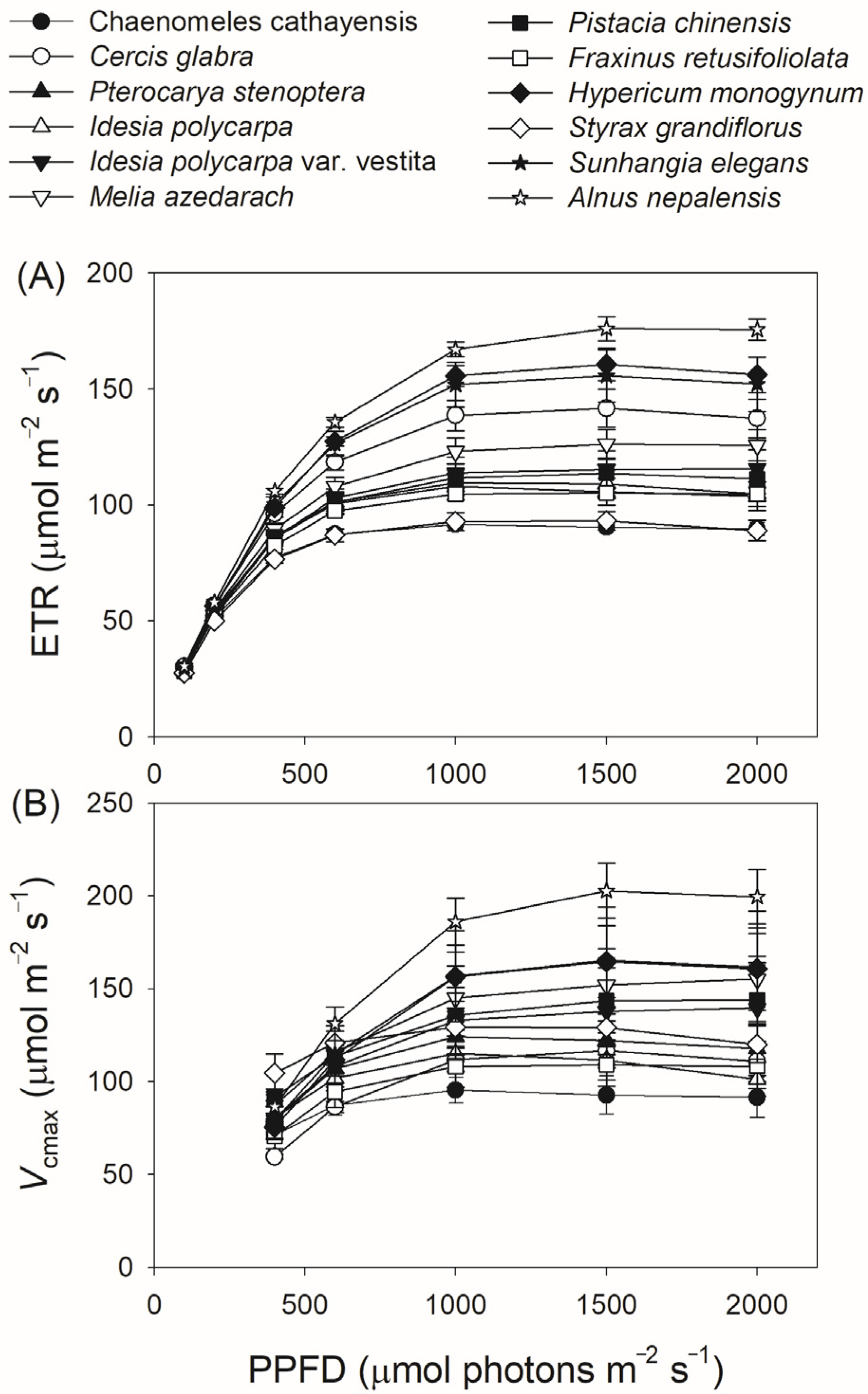
3.2. Light Intensity Dependence of Photosynthetic Parameters
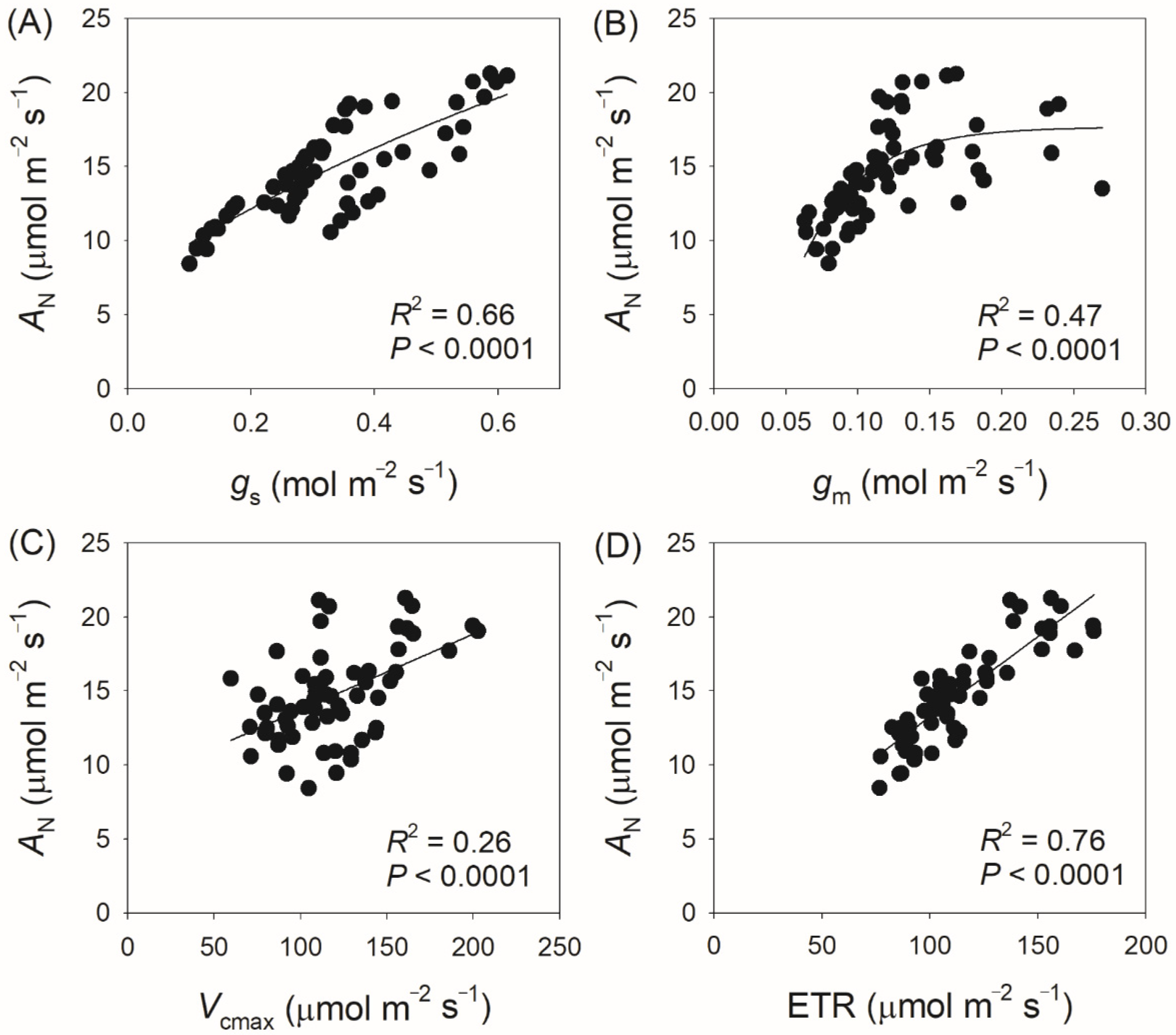
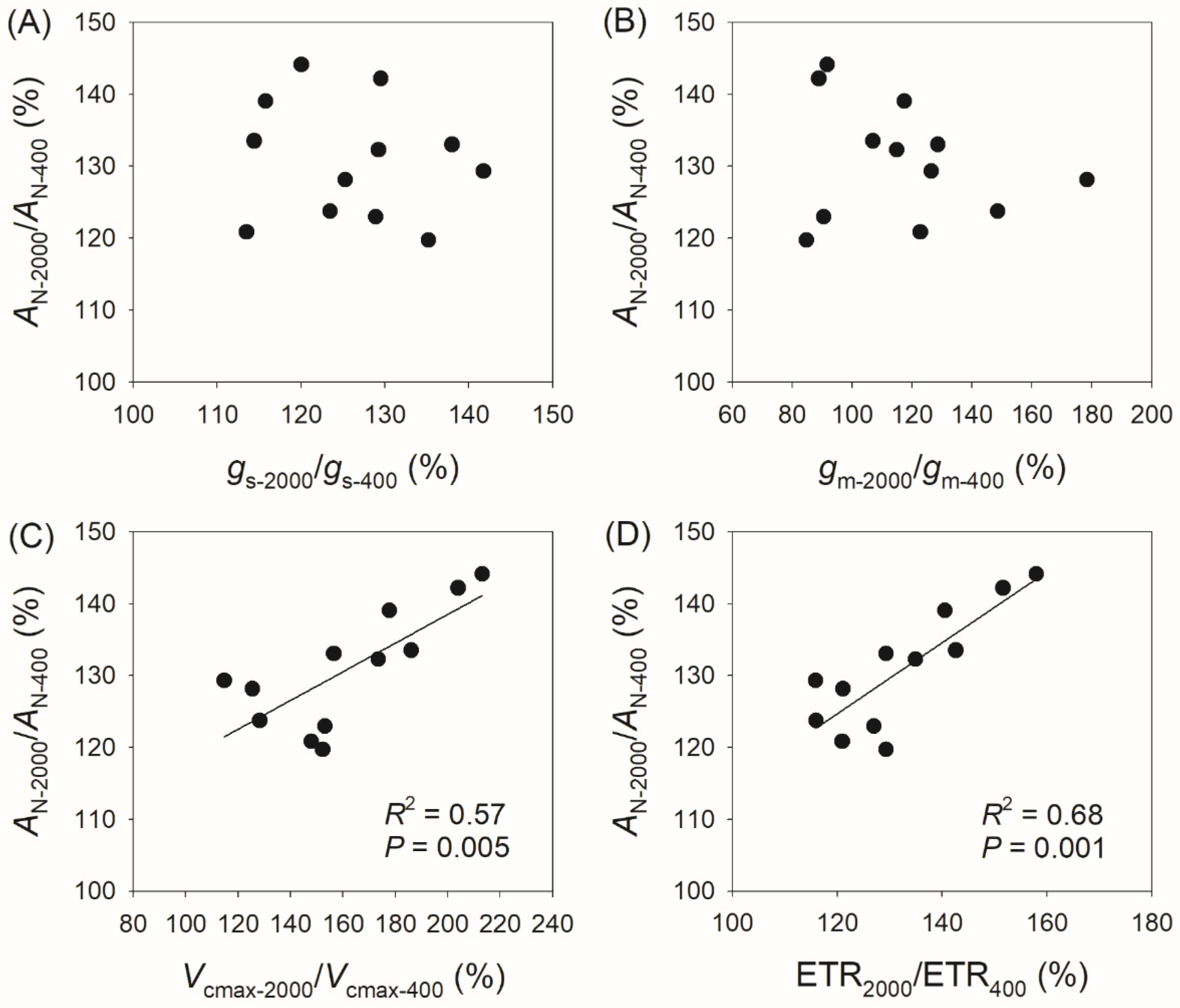
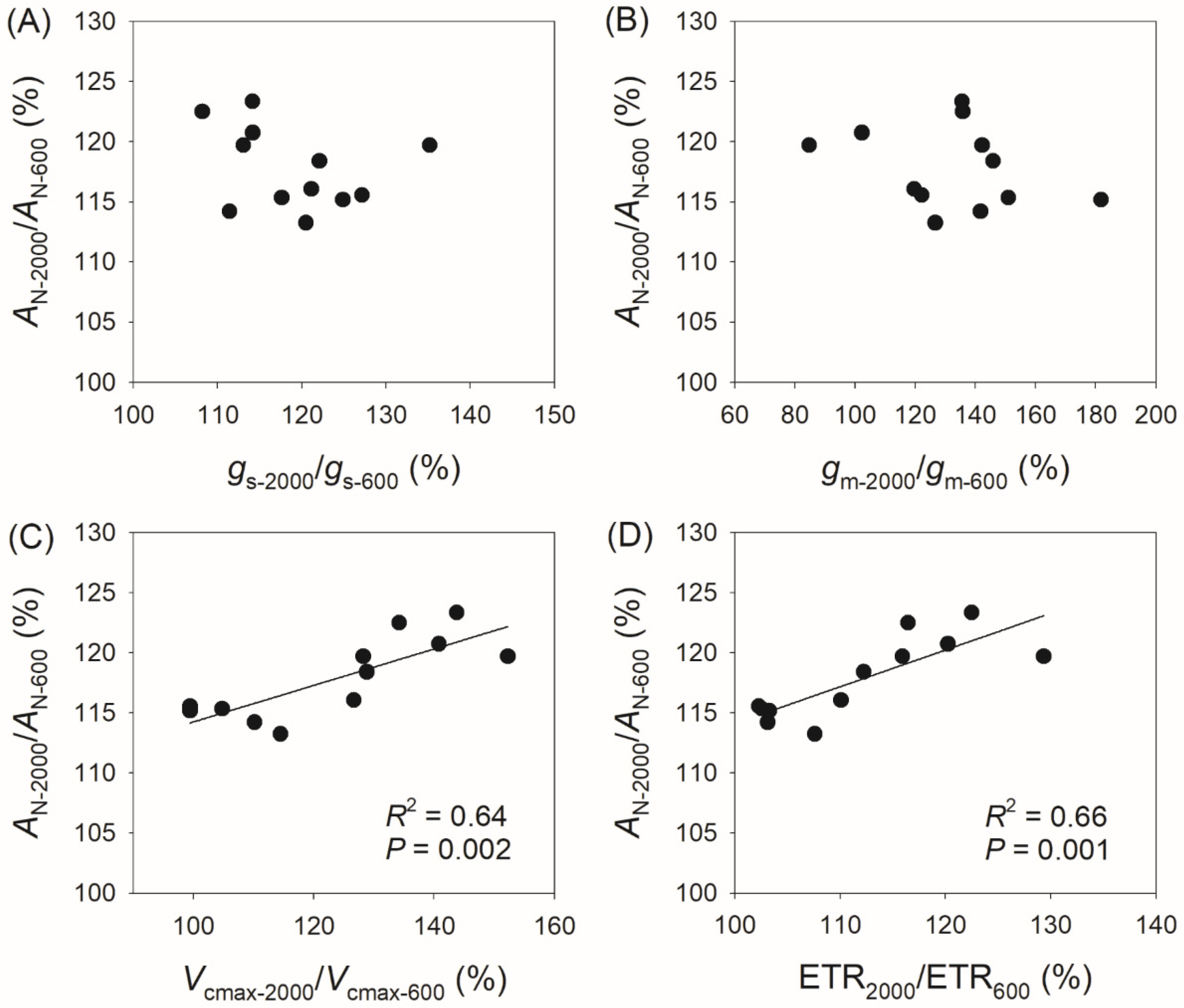
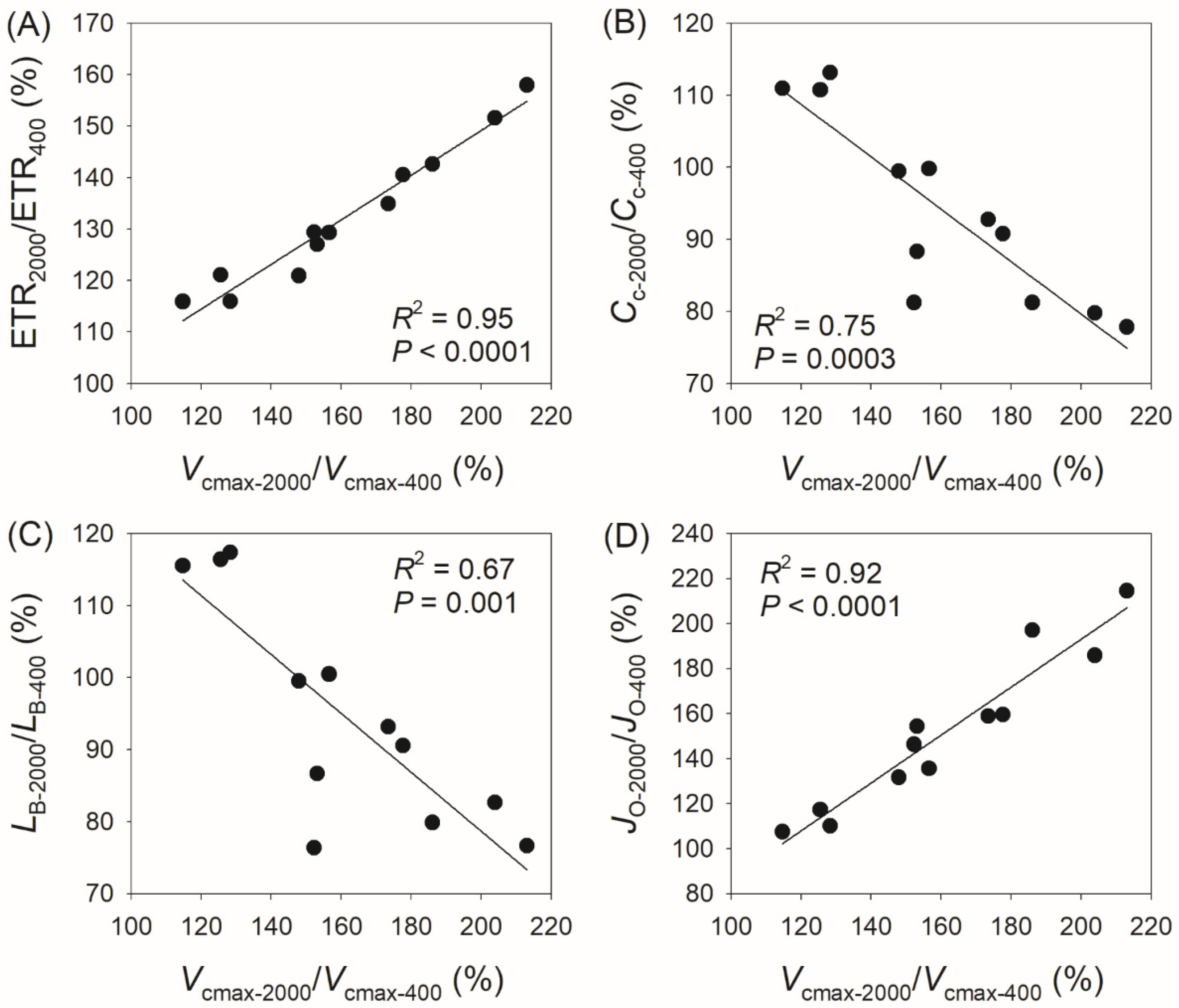
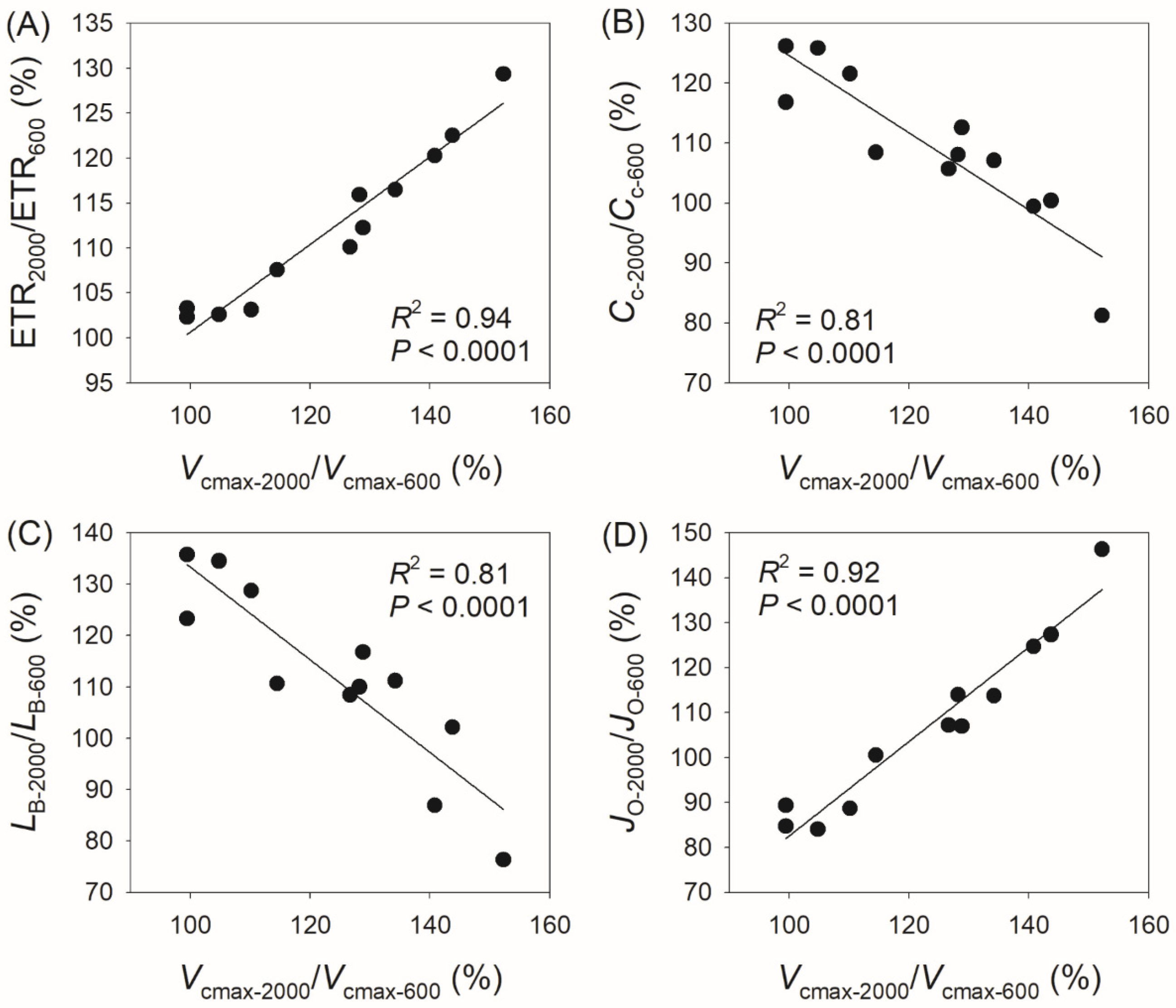
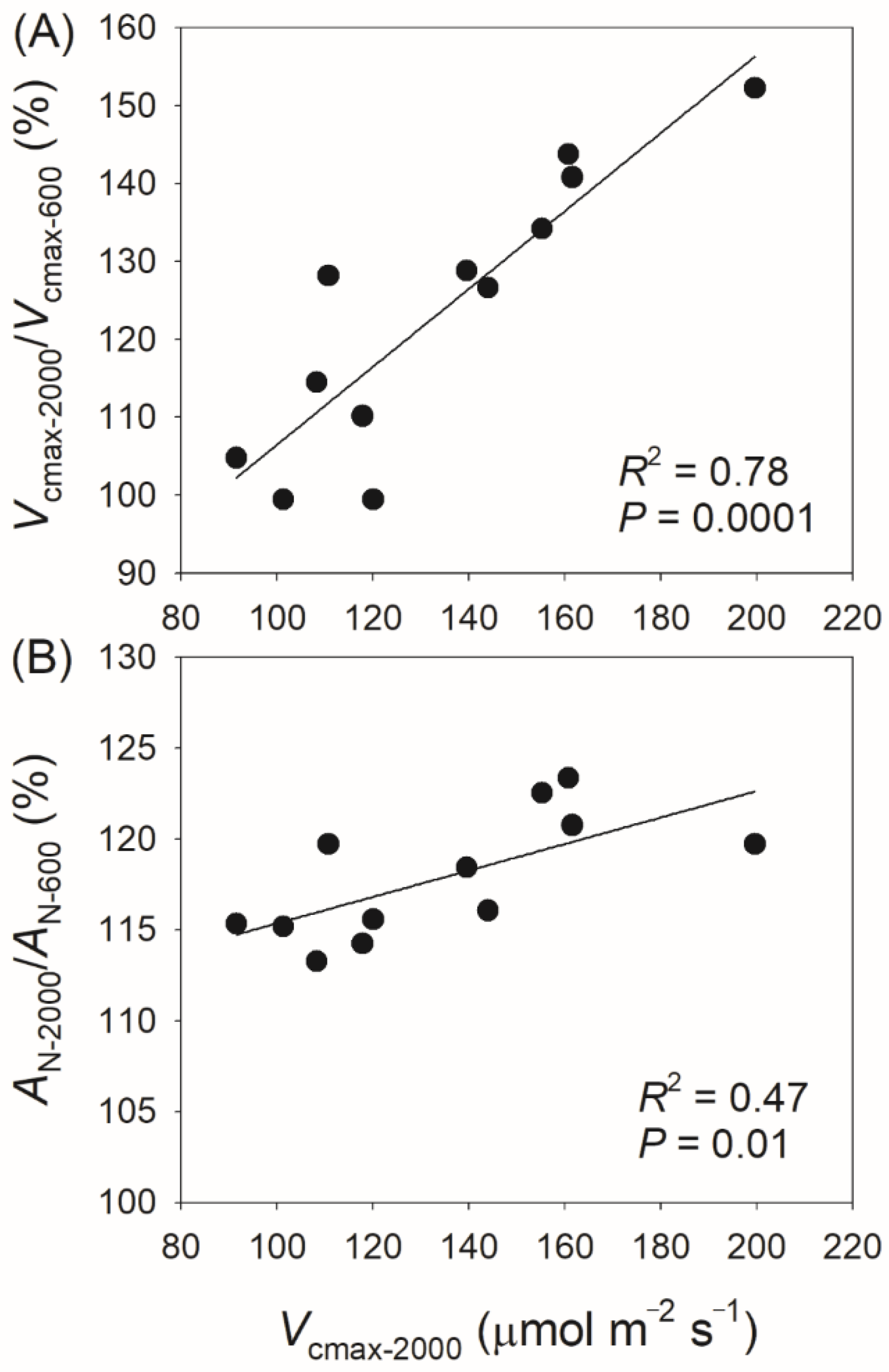
3.3. Correlation Between Physiological Parameters and CO2 Assimilation Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Carriquí, M.; Cabrera, H.M.; Conesa, M.; Coopman, R.E.; Douthe, C.; Gago, J.; Gallé, A.; Galmés, J.; Ribas-Carbo, M.; Tomás, M.; et al. Diffusional Limitations Explain the Lower Photosynthetic Capacity of Ferns as Compared with Angiosperms in a Common Garden Study. Plant Cell Environ. 2015, 38, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Gago, J.; Daloso, D.; Figueroa, C.M.; Flexas, J.; Fernie, A.R.; Nikoloski, Z. Relationships of Leaf Net Photosynthesis, Stomatal Conductance, and Mesophyll Conductance to Primary Plant Metabolism: A Multi-Species Meta-Analysis Approach. Plant Physiol. 2016, 171, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Peguero-Pina, J.J.; Sisó, S.; Flexas, J.; Galmés, J.; García-Nogales, A.; Niinemets, Ü.; Sancho-Knapik, D.; Saz, M.Á.; Gil-Pelegrín, E. Cell-Level Anatomical Characteristics Explain High Mesophyll Conductance and Photosynthetic Capacity in Sclerophyllous Mediterranean Oaks. New Phytol. 2017, 214, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, S.B.; Peguero-Pina, J.J.; Huang, W. Cell-Anatomical and Physiological Mechanisms Underlying the Faster Carbon Gain of Deciduous Trees Relative to Evergreen Trees. Environ. Exp. Bot. 2023, 209, 105286. [Google Scholar] [CrossRef]
- Shi, Q.; He, B.; Knauer, J.; Peguero-Pina, J.J.; Zhang, S.-B.; Huang, W. Leaf Nutrient Basis for the Differentiation of Photosynthetic Traits between Subtropical Evergreen and Deciduous Trees. Plant Physiol. 2025, 197, kiae566. [Google Scholar] [CrossRef]
- Pearcy, R.W. Sunflecks and Photosynthesis in Plant Canopies. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 421–453. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Krall, J.P.; Sassenrath-Cole, G.F. Photosynthesis in Fluctuating Light Environments. In Photosynthesis and the Environment; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 321–346. [Google Scholar]
- Yang, Y.-J.; Hu, H.; Huang, W. The Light Dependence of Mesophyll Conductance and Relative Limitations on Photosynthesis in Evergreen Sclerophyllous Rhododendron Species. Plants 2020, 9, 1536. [Google Scholar] [CrossRef]
- Xiong, D.; Douthe, C.; Flexas, J. Differential Coordination of Stomatal Conductance, Mesophyll Conductance, and Leaf Hydraulic Conductance in Response to Changing Light across Species. Plant Cell Environ. 2018, 41, 436–450. [Google Scholar] [CrossRef]
- Yamori, W.; Evans, J.R.; Von Caemmerer, S. Effects of Growth and Measurement Light Intensities on Temperature Dependence of CO2 Assimilation Rate in Tobacco Leaves. Plant Cell Environ. 2010, 33, 332–343. [Google Scholar] [CrossRef]
- Long, S.P.; Taylor, S.H.; Burgess, S.J.; Carmo-Silva, E.; Lawson, T.; De Souza, A.P.; Leonelli, L.; Wang, Y. Into the Shadows and Back into Sunlight: Photosynthesis in Fluctuating Light. Annu. Rev. Plant Biol. 2022, 73, 617–648. [Google Scholar] [CrossRef]
- Chang, W.; Zhang, S.B.; Li, S.Y.; Hu, H. Ecophysiological Significance of Leaf Traits in Cypripedium and Paphiopedilum. Physiol. Plant. 2011, 141, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, W.; Zhang, S.-B. The Study of a Determinate Growth Orchid Highlights the Role of New Leaf Production in Photosynthetic Light Acclimation. Plant Ecol. 2017, 218, 997–1008. [Google Scholar] [CrossRef]
- Zeng, Z.-L.; Wang, X.-Q.; Zhang, S.-B.; Huang, W. Mesophyll Conductance Limits Photosynthesis in Fluctuating Light under Combined Drought and Heat Stresses. Plant Physiol. 2024, 194, 1498–1511. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Magnani, F. Stomatal, Mesophyll Conductance and Biochemical Limitations to Photosynthesis as Affected by Drought and Leaf Ontogeny in Ash and Oak Trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Yamori, W.; Sakata, N.; Suzuki, Y.; Shikanai, T.; Makino, A. Cyclic Electron Flow around Photosystem I via Chloroplast NAD(P)H Dehydrogenase (NDH) Complex Performs a Significant Physiological Role during Photosynthesis and Plant Growth at Low Temperature in Rice. Plant J. 2011, 68, 966–976. [Google Scholar] [CrossRef]
- Yamori, W.; Nagai, T.; Makino, A. The Rate-Limiting Step for CO2 Assimilation at Different Temperatures Is Influenced by the Leaf Nitrogen Content in Several C3 Crop Species. Plant Cell Environ. 2011, 34, 764–777. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A Biochemical Model of Photosynthetic CO2 Assimilation in Leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- Zhu, X.-G.; Long, S.P.; Ort, D.R. Improving Photosynthetic Efficiency for Greater Yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef]
- Long, S.P.; Bernacchi, C.J. Gas Exchange Measurements, What Can They Tell Us about the Underlying Limitations to Photosynthesis? Procedures and Sources of Error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef]
- Yamori, W.; Kusumi, K.; Iba, K.; Terashima, I. Increased Stomatal Conductance Induces Rapid Changes to Photosynthetic Rate in Response to Naturally Fluctuating Light Conditions in Rice. Plant. Cell Environ. 2020, 43, 1230–1240. [Google Scholar] [CrossRef]
- Liu, T.; Barbour, M.M.; Yu, D.; Rao, S.; Song, X. Mesophyll Conductance Exerts a Significant Limitation on Photosynthesis during Light Induction. New Phytol. 2022, 233, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A. Light Dependence of Carboxylation Capacity for C3 Photosynthesis Models. Photosynthetica 2016, 54, 484–490. [Google Scholar] [CrossRef]
- Loriaux, S.D.; Avenson, T.J.; Welles, J.M.; Mcdermitt, D.K.; Eckles, R.D.; Riensche, B.; Genty, B. Closing in on Maximum Yield of Chlorophyll Fluorescence Using a Single Multiphase Flash of Sub-Saturating Intensity. Plant Cell Environ. 2013, 36, 1755–1770. [Google Scholar] [CrossRef] [PubMed]
- Krall, J.P.; Edwards, G.E. Relationship between Photosystem II Activity and CO2 Fixation in Leaves. Physiol. Plant. 1992, 86, 180–187. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The Relationship between the Quantum Yield of Photosynthetic Electron Transport and Quenching of Chlorophyll Fluorescence. Biochim. Biophys. Acta-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Harley, P.C.; Loreto, F.; Di Marco, G.; Sharkey, T.D. Theoretical Considerations When Estimating the Mesophyll Conductance to CO2 Flux by Analysis of the Response of Photosynthesis to CO2. Plant Physiol. 1992, 98, 1429–1436. [Google Scholar] [CrossRef]
- Flexas, J.; Ribas-Carbó, M.; Diaz-Espejo, A.; Galmés, J.; Medrano, H. Mesophyll Conductance to CO2: Current Knowledge and Future Prospects. Plant. Cell Environ. 2008, 31, 602–621. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Lin, Y.S.; Wright, I.J.; Medlyn, B.E.; Crous, K.Y.; Ellsworth, D.S.; Maire, V.; Prentice, I.C.; Atkin, O.K.; Rogers, A.; et al. A Test of the “one-Point Method” for Estimating Maximum Carboxylation Capacity from Field-Measured, Light-Saturated Photosynthesis. New Phytol. 2016, 210, 1130–1144. [Google Scholar] [CrossRef]
- Valentini, R.; Epron, D.; Angelis, P.D.E.; Matteucci, G.; Dreyer, E. In Situ Estimation of Net CO2 Assimilation, Photosynthetic Electron Flow and Photorespiration in Turkey Oak (Q. cerris L.) Leaves: Diurnal Cycles under Different Levels of Water Supply. Plant Cell Environ. 1995, 18, 631–640. [Google Scholar] [CrossRef]
- Yamori, W.; Masumoto, C.; Fukayama, H.; Makino, A. Rubisco Activase Is a Key Regulator of Non-Steady-State Photosynthesis at Any Leaf Temperature and, to a Lesser Extent, of Steady-State Photosynthesis at High Temperature. Plant J. 2012, 71, 871–880. [Google Scholar] [CrossRef]
- Kanazawa, A.; Kramer, D.M. In Vivo Modulation of Nonphotochemical Exciton Quenching (NPQ) by Regulation of the Chloroplast ATP Synthase. Proc. Natl. Acad. Sci. USA 2002, 99, 12789–12794. [Google Scholar] [CrossRef] [PubMed]
- Rott, M.; Martins, N.F.; Thiele, W.; Lein, W.; Bock, R.; Kramer, D.M.; Schöttler, M.A. ATP Synthase Repression in Tobacco Restricts Photosynthetic Electron Transport, CO2 Assimilation, and Plant Growth by Overacidification of the Thylakoid Lumen. Plant Cell 2011, 23, 304–321. [Google Scholar] [CrossRef] [PubMed]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of Photosystem II under Environmental Stress. Biochim. Biophys. Acta-Bioenerg. 2007, 1767, 414–421. [Google Scholar] [CrossRef]
- Zivcak, M.; Kalaji, H.M.; Shao, H.B.; Olsovska, K.; Brestic, M. Photosynthetic Proton and Electron Transport in Wheat Leaves under Prolonged Moderate Drought Stress. J. Photochem. Photobiol. B Biol. 2014, 137, 107–115. [Google Scholar] [CrossRef]
- Sakoda, K.; Yamori, W.; Groszmann, M.; Evans, J.R. Stomatal, Mesophyll Conductance, and Biochemical Limitations to Photosynthesis during Induction. Plant Physiol. 2021, 185, 146–160. [Google Scholar] [CrossRef]
- Salter, W.T.; Merchant, A.M.; Richards, R.A.; Trethowan, R.; Buckley, T.N. Rate of Photosynthetic Induction in Fluctuating Light Varies Widely among Genotypes of Wheat. J. Exp. Bot. 2019, 70, 2787–2796. [Google Scholar] [CrossRef]
- Smith, K.; Strand, D.D.; Kramer, D.M.; Walker, B.J. The Role of Photorespiration in Preventing Feedback Regulation via ATP Synthase in Nicotiana Tabacum. Plant. Cell Environ. 2024, 47, 416–428. [Google Scholar] [CrossRef]
- Walker, B.J.; VanLoocke, A.; Bernacchi, C.J.; Ort, D.R. The Costs of Photorespiration to Food Production Now and in the Future. Annu. Rev. Plant Biol. 2016, 67, 107–129. [Google Scholar] [CrossRef]
- Yamori, W.; Takahashi, S.; Makino, A.; Price, G.D.; Badger, M.R.; von Caemmerer, S. The Roles of ATP Synthase and the Cytochrome B6/f Complexes in Limiting Chloroplast Electron Transport and Determining Photosynthetic Capacity. Plant Physiol. 2011, 155, 956–962. [Google Scholar] [CrossRef]
- Wang, X.-Q.; Sun, H.; Zeng, Z.-L.; Huang, W. Within-Branch Photosynthetic Gradients Are More Related to the Coordinated Investments of Nitrogen and Water than Leaf Mass per Area. Plant Physiol. Biochem. 2023, 198, 107681. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Aarea (μmol m−2 s−1) | gs (mol m−2 s−1) | gm (mol m−2 s−1) | Vcmax (μmol m−2 s−1) | ETR (μmol m−2 s−1) |
|---|---|---|---|---|---|
| Ch. cathayensis | 13.1 ± 0.43 | 0.406 ± 0.031 | 0.095 ± 0.01 | 91.5 ± 11 | 89.6 ± 2.8 |
| Ce. glabra | 21.1 ± 0.85 | 0.615 ± 0.043 | 0.16 ± 0.02 | 111 ± 11 | 137 ± 8.2 |
| Pt. stenoptera | 14.7 ± 0.49 | 0.303 ± 0.042 | 0.12 ± 0.005 | 118 ± 14 | 104 ± 6.0 |
| Id. polycarpa | 16.3 ± 0.77 | 0.314 ± 0.037 | 0.16 ± 0.01 | 101 ± 9.1 | 105 ± 5.4 |
| Id. Polycarpa var. vestita | 16.0 ± 0.78 | 0.446 ± 0.039 | 0.18 ± 0.04 | 139 ± 25 | 116±8.3 |
| Me. azedarach | 16.3 ± 0.76 | 0.302 ± 0.05 | 0.12 ± 0.01 | 155 ± 25 | 126 ± 6.9 |
| Pi. chinensis | 12.5 ± 0.97 | 0.177 ± 0.037 | 0.091 ± 0.01 | 144 ± 24 | 111 ± 6.4 |
| Fr. retusifoliolata | 15.4 ± 0.23 | 0.285 ± 0.02 | 0.15 ± 0.02 | 108 ± 12 | 105 ± 3.0 |
| Hy. monogynum | 21.3 ± 0.45 | 0.588 ± 0.06 | 0.17 ± 0.02 | 161± 22 | 156 ± 7.6 |
| St. grandiflorus | 10.9 ± 0.66 | 0.142 ± 0.01 | 0.10 ± 0.01 | 120 ± 9.9 | 88.9 ± 4.3 |
| Su. elegans | 19.2 ± 0.71 | 0.359± 0.07 | 0.24 ± 0.04 | 162 ± 30 | 152 ± 12 |
| Al. nepalensis | 19.4 ± 1.6 | 0.428 ± 0.09 | 0.13 ± 0.01 | 200 ±15 | 176 ± 4.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Shi, Q.; Liu, N.; Cao, J.; Huang, W. Light Intensity Dependence of CO2 Assimilation Is More Related to Biochemical Capacity Rather than Diffusional Conductance. Plants 2025, 14, 986. https://doi.org/10.3390/plants14070986
Wang X, Shi Q, Liu N, Cao J, Huang W. Light Intensity Dependence of CO2 Assimilation Is More Related to Biochemical Capacity Rather than Diffusional Conductance. Plants. 2025; 14(7):986. https://doi.org/10.3390/plants14070986
Chicago/Turabian StyleWang, Xiaoqian, Qi Shi, Ningyu Liu, Jianxin Cao, and Wei Huang. 2025. "Light Intensity Dependence of CO2 Assimilation Is More Related to Biochemical Capacity Rather than Diffusional Conductance" Plants 14, no. 7: 986. https://doi.org/10.3390/plants14070986
APA StyleWang, X., Shi, Q., Liu, N., Cao, J., & Huang, W. (2025). Light Intensity Dependence of CO2 Assimilation Is More Related to Biochemical Capacity Rather than Diffusional Conductance. Plants, 14(7), 986. https://doi.org/10.3390/plants14070986