
Unraveling the Complexities of Flowering in Ornamental Plants: The Interplay of Genetics, Hormonal Networks, and Microbiome

 ,
,  ,
,  and
and
Abstract
1. Introduction
1.1. Brief Overview of the Importance of Flowering in Ornamental Plants
1.2. Introduction to the Genetic and Hormonal Networks Regulating Flowering
1.3. Microbial Diversity and Its Impact on Floral Biology and Pollination
2. Genetic Regulation of Flowering
2.1. Overview of Key Genes Involved in Flowering Time Regulation
2.2. Discussion of Major Flowering Pathways (Photoperiod, Vernalization, Autonomous Pathways)
2.3. Summary of Current Understanding of Genetic Regulation in Ornamental Plants
3. Hormonal Regulation of Flowering
3.1. Role of Key Hormones in Flowering
3.2. Interaction Between Hormones and Genetic Pathways in Flowering Regulation
3.3. Hormonal Crosstalk and Its Impact on Flowering Induction
3.4. Recent Advances in Hormone Research and Their Implications for Ornamental Plant Breeding
4. The Microbiome and Flowering Induction
4.1. Introduction to the Plant Microbiome and Its Importance in Plant Health and Development
4.2. Evidence Suggesting a Role of the Microbiome in Flowering Induction
4.3. Mechanisms by Which Microbiota May Influence Flowering Time (e.g., Hormone Production, Nutrient Availability)
5. Interconnection Between Endogenous and External Factors
5.1. Integration of Findings from Endogenous and External Factors Studies to Provide a Holistic Understanding of Flowering Induction
5.2. Implications for Ornamental Plant Breeding and Cultivation Practices
5.3. Future Directions for Research in Understanding the Complex Networks Underlying Flowering in Ornamental Plants
6. Summary and Future Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Volckaert, E.; Gobin, B.; Verheye, W. Ornamental plants and floriculture. In Soils, Plant Growth and Crop Production; Verheye, W.H., Ed.; EOLSS Publications: Paris, France, 2010; Volume 3. [Google Scholar]
- Yahia, E.M. Classification of horticultural commodities. In Postharvest Technology of Perishable Horticultural Commodities; Elsevier: Amsterdam, The Netherlands, 2019; pp. 71–97. [Google Scholar]
- Gabellini, S.; Scaramuzzi, S. Evolving consumption trends, marketing strategies, and governance settings in ornamental horticulture: A grey literature review. Horticulturae 2022, 8, 234. [Google Scholar] [CrossRef]
- Hendricks, J.; Briercliffe, T.; Oosterom, B.; Treer, A.; Kok, G.; Edwards, T.; Kong, H. Ornamental Horticulture, A Growing Industry. In International Vision Project Reports; AIPH Horticulture House: Chilton Didcot, Oxfordshire, UK, 2019. [Google Scholar]
- Löbke, A. Record Sales for the Flower and Plant Market; Messe Essen GmbH: Essen, Germany, 2022. [Google Scholar]
- Bugallo, V.; Facciuto, G. Selection process in ornamental plant breeding. Ornam. Hortic. 2023, 29, 68–75. [Google Scholar] [CrossRef]
- Arslan, M.; Yanmaz, R. Use of ornamental vegetables, medicinal and aromatic plants in urban landscape design. In Proceedings of the II International Conference on Landscape and Urban Horticulture 881, Bologna, Italy, 9–13 June 2009; pp. 207–211. [Google Scholar]
- Coyago-Cruz, E.; Moya, M.; Méndez, G.; Villacís, M.; Rojas-Silva, P.; Corell, M.; Mapelli-Brahm, P.; Vicario, I.M.; Meléndez-Martínez, A.J. Exploring Plants with Flowers: From Therapeutic Nutritional Benefits to Innovative Sustainable Uses. Foods 2023, 12, 4066. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.J.A.J. Allusions to Floral Emblem of G20 Nations. Int. J. Multidiscip. Res. 2023, 5. [Google Scholar] [CrossRef]
- Fernandes, L.; Casal, S.; Pereira, J.A.; Saraiva, J.A.; Ramalhosa, E. An overview on the market of edible flowers. Food Rev. Int. 2020, 36, 258–275. [Google Scholar] [CrossRef]
- Urakami, J.; Huss, E.; Nagamine, M.; Czamanski-Cohen, J.; Zaccai, M. The Emotional Experience of Flowers: Zoomed In, Zoomed Out and Painted. Horticulturae 2022, 8, 668. [Google Scholar] [CrossRef]
- Sood, Y.; Lal, M.; Kalia, A.; Verma, S. Edible Flowers: Super Foods with Potential Health Benefits. Int. J. Plant Soil Sci. 2024, 36, 213–221. [Google Scholar] [CrossRef]
- Marchioni, I.; Gabriele, M.; Carmassi, G.; Ruffoni, B.; Pistelli, L.; Pistelli, L.; Najar, B. Phytochemical, Nutritional and Mineral Content of Four Edible Flowers. Foods 2024, 13, 939. [Google Scholar] [CrossRef]
- Jadhav, H.B.; Badwaik, L.S.; Annapure, U.; Casanova, F.; Alaskar, K. A review on the journey of edible flowers from farm to consumer’s plate. Appl. Food Res. 2023, 3, 100312. [Google Scholar] [CrossRef]
- Pires, E.D.O., Jr.; Di Gioia, F.; Rouphael, Y.; García-Caparrós, P.; Tzortzakis, N.; Ferreira, I.C.; Barros, L.; Petropoulos, S.A.; Caleja, C. Edible flowers as an emerging horticultural product: A review on sensorial properties, mineral and aroma profile. Trends Food Sci. Technol. 2023, 137, 31–54. [Google Scholar]
- Chávez-Hernández, E.C.; Quiroz, S.; García-Ponce, B.; Álvarez-Buylla, E.R. The flowering transition pathways converge into a complex gene regulatory network that underlies the phase changes of the shoot apical meristem in Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 852047. [Google Scholar] [CrossRef]
- Sasaki, E.; Frommlet, F.; Nordborg, M. The genetic architecture of the network underlying flowering time variation in Arabidopsis thaliana. bioRxiv 2017. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, E.; Liu, Q.A.; Wang, J. High CO2 adaptation mechanisms revealed in the miR156-regulated flowering time pathway. PLoS Comput. Biol. 2023, 19, e1011738. [Google Scholar]
- He, T.; Hill, C.B.; Angessa, T.T.; Zhang, X.-Q.; Chen, K.; Moody, D.; Telfer, P.; Westcott, S.; Li, C. Gene-set association and epistatic analyses reveal complex gene interaction networks affecting flowering time in a worldwide barley collection. J. Exp. Bot. 2019, 70, 5603–5616. [Google Scholar] [PubMed]
- Tran, L.M.H. The effect of phytohormones on the flowering of plants. Plant Sci. Today 2023, 10, 138–142. [Google Scholar]
- Wang, X.; Zhao, F.; Wu, Q.; Xing, S.; Yu, Y.; Qi, S. Physiological and transcriptome analyses to infer regulatory networks in flowering transition of Rosa rugosa. Ornam. Plant Res. 2023, 3, 4. [Google Scholar]
- Goh, C.; Yang, A. Effects of growth regulators and decapitation on flowering of Dendrobium orchid hybrids. Plant Sci. Lett. 1978, 12, 287–292. [Google Scholar] [CrossRef]
- Bhalerao, R.P.; Bennett, M.J. The case for morphogens in plants. Nat. Cell Biol. 2003, 5, 939–943. [Google Scholar]
- Zoulias, N.; Duttke, S.H.; Garcês, H.; Spencer, V.; Kim, M. The role of auxin in the pattern formation of the Asteraceae flower head (capitulum). Plant Physiol. 2019, 179, 391–401. [Google Scholar]
- Blanchard, M.G.; Runkle, E.S. Benzyladenine promotes flowering in Doritaenopsis and Phalaenopsis orchids. J. Plant Growth Regul. 2008, 27, 141–150. [Google Scholar]
- Su, W.-R.; Chen, W.-S.; Koshioka, M.; Mander, L.N.; Hung, L.-S.; Chen, W.-H.; Fu, Y.-M.; Huang, K.-L. Changes in gibberellin levels in the flowering shoot of Phalaenopsis hybrida under high temperature conditionswhen flower development is blocked. Plant Physiol. Biochem. 2001, 39, 45–50. [Google Scholar]
- Hew, C.; Clifford, P. Plant growth regulators and the orchid cut-flower industry. Plant Growth Regul. 1993, 13, 231–239. [Google Scholar] [CrossRef]
- Li, J.; Jiang, J.; Qian, Q.; Xu, Y.; Zhang, C.; Xiao, J.; Du, C.; Luo, W.; Zou, G.; Chen, M. Mutation of rice BC12/GDD1, which encodes a kinesin-like protein that binds to a GA biosynthesis gene promoter, leads to dwarfism with impaired cell elongation. Plant Cell 2011, 23, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Ye, T.; Lu, Y.; Chen, X.; Wu, Y. The inhibitory effect of ABA on floral transition is mediated by ABI5 in Arabidopsis. J. Exp. Bot. 2013, 64, 675–684. [Google Scholar] [CrossRef]
- Bouwmeester, H.J.; Matusova, R.; Zhongkui, S.; Beale, M.H. Secondary metabolite signalling in host–parasitic plant interactions. Curr. Opin. Plant Biol. 2003, 6, 358–364. [Google Scholar] [CrossRef]
- Ruttink, T.; Arend, M.; Morreel, K.; Storme, V.; Rombauts, S.; Fromm, J.; Bhalerao, R.P.; Boerjan, W.; Rohde, A. A molecular timetable for apical bud formation and dormancy induction in poplar. Plant Cell 2007, 19, 2370–2390. [Google Scholar] [CrossRef]
- Tylewicz, S.; Petterle, A.; Marttila, S.; Miskolczi, P.; Azeez, A.; Singh, R.K.; Immanen, J.; Mähler, N.; Hvidsten, T.R.; Eklund, D.M. Photoperiodic control of seasonal growth is mediated by ABA acting on cell-cell communication. Science 2018, 360, 212–215. [Google Scholar] [PubMed]
- Ahmad, S.; Lu, C.; Gao, J.; Ren, R.; Wei, Y.; Wu, J.; Jin, J.; Zheng, C.; Zhu, G.; Yang, F. Genetic insights into the regulatory pathways for continuous flowering in a unique orchid Arundina graminifolia. BMC Plant Biol. 2021, 21, 587. [Google Scholar] [CrossRef]
- Wang, W.-Y.; Chen, W.-S.; Chen, W.-H.; Hung, L.-S.; Chang, P.-S. Influence of abscisic acid on flowering in Phalaenopsis hybrida. Plant Physiol. Biochem. 2002, 40, 97–100. [Google Scholar] [CrossRef]
- Burgess, E.C.; Schaeffer, R.N. The floral microbiome and its management in agroecosystems: A perspective. J. Agric. Food Chem. 2022, 70, 9819–9825. [Google Scholar] [CrossRef]
- Brysch-Herzberg, M. Ecology of yeasts in plant–bumblebee mutualism in Central Europe. FEMS Microbiol. Ecol. 2004, 50, 87–100. [Google Scholar] [CrossRef]
- Chappell, C.R.; Fukami, T. Nectar yeasts: A natural microcosm for ecology. Yeast 2018, 35, 417–423. [Google Scholar] [CrossRef]
- Figueroa, L.L.; Blinder, M.; Grincavitch, C.; Jelinek, A.; Mann, E.K.; Merva, L.A.; Metz, L.E.; Zhao, A.Y.; Irwin, R.E.; McArt, S.H. Bee pathogen transmission dynamics: Deposition, persistence and acquisition on flowers. Proc. R. Soc. B 2019, 286, 20190603. [Google Scholar] [PubMed]
- Lindow, S.E.; Suslow, T.V. Temporal dynamics of the biocontrol agent Pseudomonas fluorescens strain A506 in flowers in inoculated pear trees. Phytopathology 2003, 93, 727–737. [Google Scholar] [CrossRef] [PubMed]
- von Arx, M.; Moore, A.; Davidowitz, G.; Arnold, A.E. Diversity and distribution of microbial communities in floral nectar of two night-blooming plants of the Sonoran Desert. PLoS ONE 2019, 14, e0225309. [Google Scholar] [CrossRef]
- Junker, R.R.; Loewel, C.; Gross, R.; Dötterl, S.; Keller, A.; Blüthgen, N. Composition of epiphytic bacterial communities differs on petals and leaves. Plant Biol. 2011, 13, 918–924. [Google Scholar] [CrossRef] [PubMed]
- Hinton, D.; Bacon, C. The distribution and ultrastructure of the endophyte of toxic tall fescue. Can. J. Bot. 1985, 63, 36–42. [Google Scholar]
- Hodgson, S.; de Cates, C.; Hodgson, J.; Morley, N.J.; Sutton, B.C.; Gange, A.C. Vertical transmission of fungal endophytes is widespread in forbs. Ecol. Evol. 2014, 4, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Peay, K.G.; Belisle, M.; Fukami, T. Phylogenetic relatedness predicts priority effects in nectar yeast communities. Proc. R. Soc. B Biol. Sci. 2012, 279, 749–758. [Google Scholar]
- Morris, M.M.; Frixione, N.J.; Burkert, A.C.; Dinsdale, E.A.; Vannette, R.L. Microbial abundance, composition, and function in nectar are shaped by flower visitor identity. FEMS Microbiol. Ecol. 2020, 96, fiaa003. [Google Scholar] [CrossRef] [PubMed]
- Boutroux, L. Sur la conservation des ferments alcooliques dans la nature. Ann. Des Sci. Nat. Série IV Bot. 1884, 17, 145–209. [Google Scholar]
- Belisle, M.; Peay, K.G.; Fukami, T. Flowers as islands: Spatial distribution of nectar-inhabiting microfungi among plants of Mimulus aurantiacus, a hummingbird-pollinated shrub. Microb. Ecol. 2012, 63, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Lachance, M.-A.; Starmer, W.T.; Rosa, C.A.; Bowles, J.M.; Barker, J.S.F.; Janzen, D.H. Biogeography of the yeasts of ephemeral flowers and their insects. FEMS Yeast Res. 2001, 1, 1–8. [Google Scholar] [PubMed]
- Samuni-Blank, M.; Izhaki, I.; Laviad, S.; Bar-Massada, A.; Gerchman, Y.; Halpern, M. The role of abiotic environmental conditions and herbivory in shaping bacterial community composition in floral nectar. PLoS ONE 2014, 9, e99107. [Google Scholar]
- Chanbusarakum, L.; Ullman, D. Characterization of bacterial symbionts in Frankliniella occidentalis (Pergande), Western flower thrips. J. Invertebr. Pathol. 2008, 99, 318–325. [Google Scholar] [CrossRef]
- Ambika Manirajan, B.; Ratering, S.; Rusch, V.; Schwiertz, A.; Geissler-Plaum, R.; Cardinale, M.; Schnell, S. Bacterial microbiota associated with flower pollen is influenced by pollination type, and shows a high degree of diversity and species-specificity. Environ. Microbiol. 2016, 18, 5161–5174. [Google Scholar]
- Russell, A.L.; Rebolleda-Gómez, M.; Shaible, T.M.; Ashman, T.L. Movers and shakers: Bumble bee foraging behavior shapes the dispersal of microbes among and within flowers. Ecosphere 2019, 10, e02714. [Google Scholar]
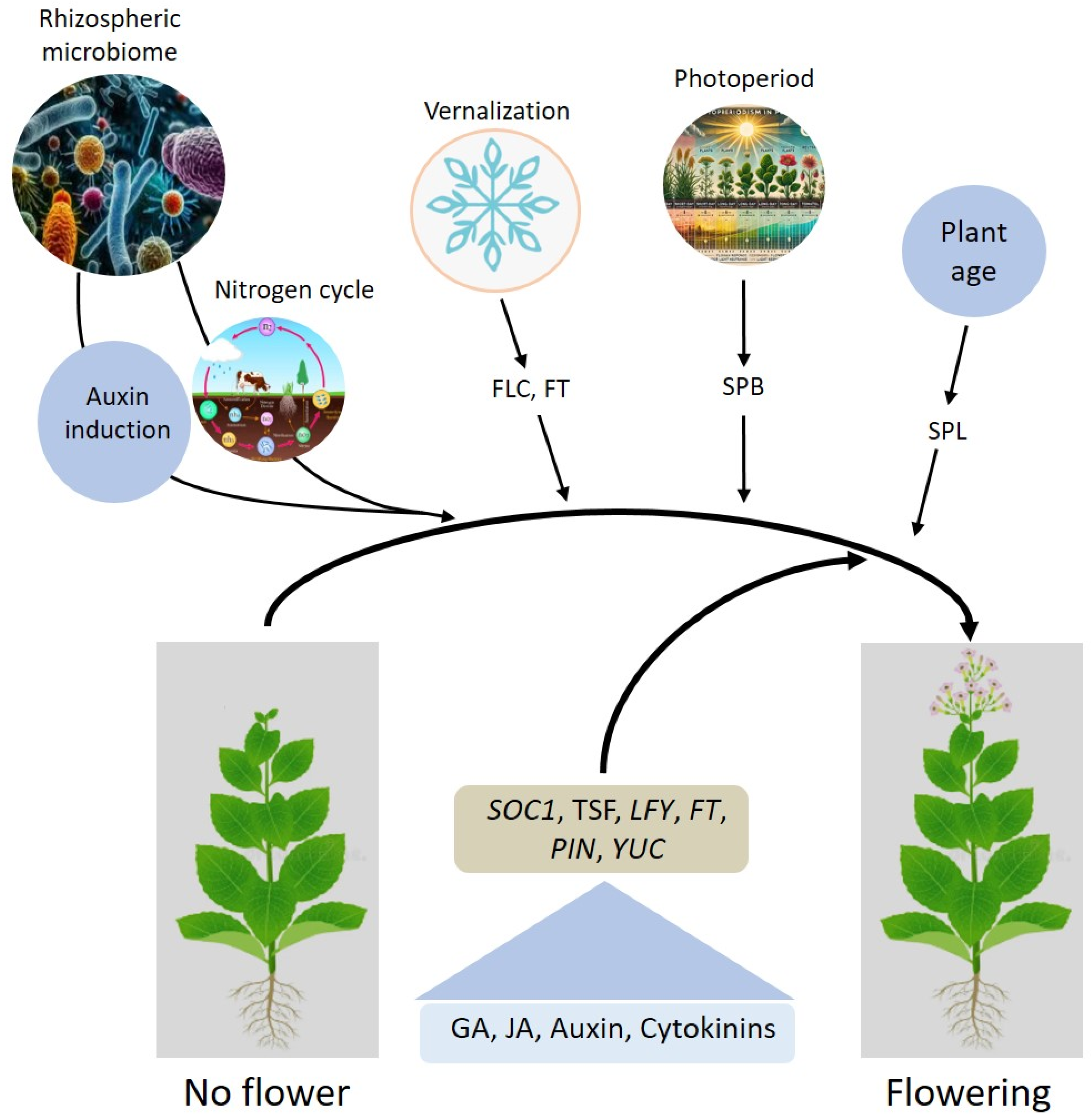
- Lu, T.; Ke, M.; Lavoie, M.; Jin, Y.; Fan, X.; Zhang, Z.; Fu, Z.; Sun, L.; Gillings, M.; Peñuelas, J. Rhizosphere microorganisms can influence the timing of plant flowering. Microbiome 2018, 6, 231. [Google Scholar]
- Putterill, J.; Laurie, R.; Macknight, R. It’s time to flower: The genetic control of flowering time. Bioessays 2004, 26, 363–373. [Google Scholar]
- Amasino, R.M.; Michaels, S.D. The timing of flowering. Plant Physiol. 2010, 154, 516–520. [Google Scholar] [PubMed]
- Colleoni, P.E.; van Es, S.W.; Winkelmolen, T.; Immink, R.G.; van Esse, G.W. Flowering time genes branching out. J. Exp. Bot. 2024, 75, 4195–4209. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, J.; Yin, H.; Li, X.; Liu, W.; Fan, Z. Function of FT in Flowering Induction in Two Camellia Species. Plants 2024, 13, 784. [Google Scholar] [CrossRef]
- Liu, B.; Woods, D.P.; Li, W.; Amasino, R.M. INDETERMINATE1-mediated expression of FT family genes is required for proper timing of flowering in Brachypodium distachyon. Proc. Natl. Acad. Sci. USA 2023, 120, e2312052120. [Google Scholar] [CrossRef]
- Jun, S.E.; Manzoor, M.A.; Kim, M.-J.; Youn, Y.; Nam, J.; Hyung, N.-I.; Kim, G.-T. Molecular cloning and functional characterization of CmSOC1 gene and its promoter region from Chrysanthemum morifolium. Sci. Hortic. 2024, 329, 112991. [Google Scholar]
- Poulet, A.; Zhao, M.; Peng, Y.; Tham, F.; Jaudal, M.; Zhang, L.; van Wolfswinkel, J.C.; Putterill, J. Gene-edited Mtsoc1 triple mutant Medicago plants do not flower. Front. Plant Sci. 2024, 15, 1357924. [Google Scholar] [CrossRef]
- Li, X.; Ping, A.; Qi, X.; Li, M.; Hou, L. Cloning, expression and functional analysis of the SOC1 homologous gene in pak choi (Brassica rapa ssp. Chinensis makino). Biotechnol. Biotechnol. Equip. 2022, 36, 848–857. [Google Scholar] [CrossRef]
- Jue, D.; Li, Z.; Zhang, W.; Tang, J.; Xie, T.; Sang, X.; Guo, Q. Identification and functional analysis of the LEAFY gene in longan flower induction. BMC Genom. 2024, 25, 308. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Li, Y.; Yang, Y.; Zhou, Y.; Zhao, K.; Zhang, Q. Isolation, functional characterization and evolutionary study of LFY1 gene in Prunus mume. Plant Cell Tissue Organ Cult. 2019, 136, 523–536. [Google Scholar] [CrossRef]
- Zhang, T.; Chao, Y.; Kang, J.; Ding, W.; Yang, Q. Molecular cloning and characterization of a gene regulating flowering time from Alfalfa (Medicago sativa L.). Mol. Biol. Rep. 2013, 40, 4597–4603. [Google Scholar] [CrossRef]
- Mahima; Chatterjee, S.; Singh, S.; Sarkar, A.K. LDL1 and LDL2 histone demethylases interact with FVE to regulate flowering in Arabidopsis. bioRxiv 2022. [Google Scholar] [CrossRef]
- Rédei, G.P. Supervital mutants of Arabidopsis. Genetics 1962, 47, 443. [Google Scholar] [PubMed]
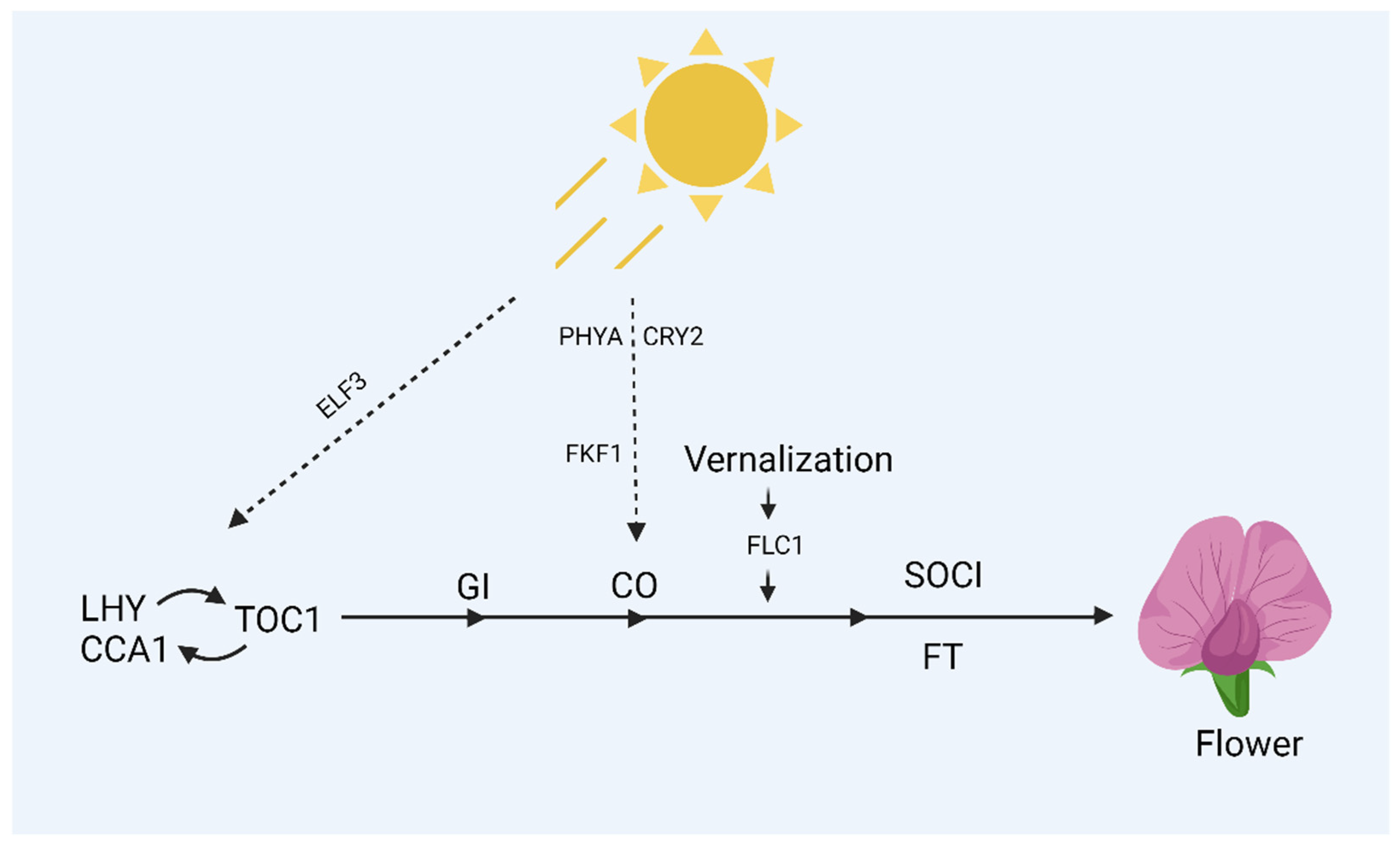
- Harmer, S.L.; Hogenesch, J.B.; Straume, M.; Chang, H.-S.; Han, B.; Zhu, T.; Wang, X.; Kreps, J.A.; Kay, S.A. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 2000, 290, 2110–2113. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-Y.; Tobin, E.M. Constitutive expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) gene disrupts circadian rhythms and suppresses its own expression. Cell 1998, 93, 1207–1217. [Google Scholar] [PubMed]
- Alabadí, D.; Oyama, T.; Yanovsky, M.J.; Harmon, F.G.; Más, P.; Kay, S.A. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 2001, 293, 880–883. [Google Scholar] [CrossRef]
- Mizoguchi, T.; Wheatley, K.; Hanzawa, Y.; Wright, L.; Mizoguchi, M.; Song, H.-R.; Carré, I.A.; Coupland, G. LHY and CCA1 are partially redundant genes required to maintain circadian rhythms in Arabidopsis. Dev. Cell 2002, 2, 629–641. [Google Scholar] [CrossRef]
- Hicks, K.A.; Albertson, T.M.; Wagner, D.R. EARLY FLOWERING3 encodes a novel protein that regulates circadian clock function and flowering in Arabidopsis. Plant Cell 2001, 13, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Peng, D.; Zhou, Y.; Zhao, K. The genetic and hormonal inducers of continuous flowering in orchids: An emerging view. Cells 2022, 11, 657. [Google Scholar] [CrossRef] [PubMed]
- Amasino, R.M. Vernalization and flowering time. Curr. Opin. Biotechnol. 2005, 16, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-H. Current understanding of flowering pathways in plants: Focusing on the vernalization pathway in Arabidopsis and several vegetable crop plants. Hortic. Environ. Biotechnol. 2020, 61, 209–227. [Google Scholar] [CrossRef]
- Choi, K.; Kim, J.; Hwang, H.-J.; Kim, S.; Park, C.; Kim, S.Y.; Lee, I. The FRIGIDA complex activates transcription of FLC, a strong flowering repressor in Arabidopsis, by recruiting chromatin modification factors. Plant Cell 2011, 23, 289–303. [Google Scholar] [CrossRef]
- Henderson, I.R.; Shindo, C.; Dean, C. The need for winter in the switch to flowering. Annu. Rev. Genet. 2003, 37, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Bastow, R.; Mylne, J.S.; Lister, C.; Lippman, Z.; Martienssen, R.A.; Dean, C. Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 2004, 427, 164–167. [Google Scholar] [CrossRef]
- Molitor, A.; Shen, W.-H. The polycomb complex PRC1: Composition and function in plants. J. Genet. Genom. 2013, 40, 231–238. [Google Scholar] [CrossRef]
- Wood, C.C.; Robertson, M.; Tanner, G.; Peacock, W.J.; Dennis, E.S.; Helliwell, C.A. The Arabidopsis thaliana vernalization response requires a polycomb-like protein complex that also includes VERNALIZATION INSENSITIVE 3. Proc. Natl. Acad. Sci. USA 2006, 103, 14631–14636. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.B.; Sung, S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 2011, 331, 76–79. [Google Scholar] [CrossRef]
- Li, M.; Wen, Z.; Meng, J.; Cheng, T.; Zhang, Q.; Sun, L. The genomics of ornamental plants: Current status and opportunities. Ornam. Plant Res. 2022, 2, 6. [Google Scholar] [CrossRef]
- Shahri, W.; Gul, F.; Tahir, I. Ornamental Plants: Some Molecular Aspects. In The Global Floriculture Industry; Apple Academic Press: Palm Bay, FL, USA, 2020; pp. 59–81. [Google Scholar]
- Mekapogu, M.; Song, H.-Y.; Lim, S.-H.; Jung, J.-A. Genetic Engineering and Genome Editing Advances to Enhance Floral Attributes in Ornamental Plants: An Update. Plants 2023, 12, 3983. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Zhang, F.; Guan, J.; Ma, Y. Genetic and epigenetic regulation of disease resistance in horticultural plants. Front. Genet. 2023, 14, 1277571. [Google Scholar] [CrossRef] [PubMed]
- Meijón, M.; Jesús Cañal, M.; Valledor, L.; Rodríguez, R.; Feito, I. Epigenetic and physiological effects of gibberellin inhibitors and chemical pruners on the floral transition of azalea. Physiol. Plant. 2011, 141, 276–288. [Google Scholar] [CrossRef]
- Gallego-Giraldo, L.; García-Martínez, J.L.; Moritz, T.; López-Díaz, I. Flowering in tobacco needs gibberellins but is not promoted by the levels of active GA1 and GA4 in the apical shoot. Plant Cell Physiol. 2007, 48, 615–625. [Google Scholar]
- Sandoval-Oliveros, R.; Guevara-Olvera, L.; Beltrán, J.; Gómez-Mena, C.; Acosta-García, G. Developmental landmarks during floral ontogeny of jalapeño chili pepper (Capsicum annuum L.) and the effect of gibberellin on ovary growth. Plant Reprod. 2017, 30, 119–129. [Google Scholar]
- Zhang, D.; Ren, L.; Yue, J.-H.; Wang, L.; Zhuo, L.-H.; Shen, X.-H. GA4 and IAA were involved in the morphogenesis and development of flowers in Agapanthus praecox ssp. orientalis. J. Plant Physiol. 2014, 171, 966–976. [Google Scholar] [PubMed]
- Cheng, Y.; Zhao, Y. A role for auxin in flower development. J. Integr. Plant Biol. 2007, 49, 99–104. [Google Scholar]
- Yamaguchi, N.; Jeong, C.W.; Nole-Wilson, S.; Krizek, B.A.; Wagner, D. AINTEGUMENTA and AINTEGUMENTA-LIKE6/PLETHORA3 induce LEAFY expression in response to auxin to promote the onset of flower formation in Arabidopsis. Plant Physiol. 2016, 170, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Enomoto, K.; Fukunaga, Y.; Kuo, C. Regeneration of tepals, stamens and ovules in explants from perianth of Hyacinthus orientalis L. importance of explant age and exogenous hormones. Planta 1988, 175, 478–484. [Google Scholar] [PubMed]
- Shu, K.; Chen, Q.; Wu, Y.; Liu, R.; Zhang, H.; Wang, S.; Tang, S.; Yang, W.; Xie, Q. ABSCISIC ACID-INSENSITIVE 4 negatively regulates flowering through directly promoting Arabidopsis FLOWERING LOCUS C transcription. J. Exp. Bot. 2016, 67, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Renau-Morata, B.; Nebauer, S.G.; García-Carpintero, V.; Canizares, J.; Minguet, E.G.; De los Mozos, M.; Molina, R.V. Flower induction and development in saffron: Timing and hormone signalling pathways. Ind. Crops Prod. 2021, 164, 113370. [Google Scholar]
- Mutasa-Göttgens, E.; Hedden, P. Gibberellin as a factor in floral regulatory networks. J. Exp. Bot. 2009, 60, 1979–1989. [Google Scholar] [PubMed]
- Griffiths, J.; Murase, K.; Rieu, I.; Zentella, R.; Zhang, Z.-L.; Powers, S.J.; Gong, F.; Phillips, A.L.; Hedden, P.; Sun, T.-P. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell 2006, 18, 3399–3414. [Google Scholar]
- Rieu, I.; Ruiz-Rivero, O.; Fernandez-Garcia, N.; Griffiths, J.; Powers, S.J.; Gong, F.; Linhartova, T.; Eriksson, S.; Nilsson, O.; Thomas, S.G. The gibberellin biosynthetic genes AtGA20ox1 and AtGA20ox2 act, partially redundantly, to promote growth and development throughout the Arabidopsis life cycle. Plant J. 2008, 53, 488–504. [Google Scholar] [PubMed]
- Nester, J.E.; Zeevaart, J.A. Flower development in normal tomato and a gibberellin-deficient (ga-2) mutant. Am. J. Bot. 1988, 75, 45–55. [Google Scholar]
- Plack, A. Effect of gibberellic acid on corolla size. Nature 1958, 182, 610. [Google Scholar] [CrossRef]
- Hu, J.; Mitchum, M.G.; Barnaby, N.; Ayele, B.T.; Ogawa, M.; Nam, E.; Lai, W.-C.; Hanada, A.; Alonso, J.M.; Ecker, J.R. Potential sites of bioactive gibberellin production during reproductive growth in Arabidopsis. Plant Cell 2008, 20, 320–336. [Google Scholar]
- Cheng, H.; Qin, L.; Lee, S.; Fu, X.; Richards, D.E.; Cao, D.; Luo, D.; Harberd, N.P.; Peng, J. Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function. Development 2004, 131, 1055–1064. [Google Scholar] [PubMed]
- Song, S.; Qi, T.; Huang, H.; Xie, D. Regulation of stamen development by coordinated actions of jasmonate, auxin, and gibberellin in Arabidopsis. Mol. Plant 2013, 6, 1065–1073. [Google Scholar]
- Yari, V.; Roein, Z.; Sabouri, A. Exogenous 5-azaCitidine accelerates flowering and external GA3 increases ornamental value in Iranian Anemone accessions. Sci. Rep. 2021, 11, 7478. [Google Scholar]
- Brooking, I.; Jamieson, P. Temperature and photoperiod response of vernalization in near-isogenic lines of wheat. Field Crops Res. 2002, 79, 21–38. [Google Scholar]
- Izhaki, A.; Borochov, A.; Zamski, E.; Weiss, D. Gibberellin regulates post-microsporogenesis processes in petunia anthers. Physiol. Plant. 2002, 115, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Chhun, T.; Aya, K.; Asano, K.; Yamamoto, E.; Morinaka, Y.; Watanabe, M.; Kitano, H.; Ashikari, M.; Matsuoka, M.; Ueguchi-Tanaka, M. Gibberellin regulates pollen viability and pollen tube growth in rice. Plant Cell 2007, 19, 3876–3888. [Google Scholar] [CrossRef] [PubMed]
- Barendse, G.; Rodrigues Pereira, A.; Berkers, P.; Driessen, F.; van Eyden-Emons, A.; Linskens, H. Growth hormones in pollen, styles and ovaries of Petunia hybrida and of Lilium species. Acta Bot. Neerl. 1970, 19, 175–186. [Google Scholar] [CrossRef]
- Fuchs, E.; Atsmon, D.; Halevy, A.H. Adventitious staminate flower formation in gibberellin treated gynoecious cucumber plants. Plant Cell Physiol. 1977, 18, 1193–1201. [Google Scholar] [CrossRef]
- Galun, E. Effects of gibberellic acid and naphthalene-acetic acid on sex expression and some morphological characters in the cucumber plant. Phyton 1959, 13, 1–8. [Google Scholar]
- Banarjee, S.; Basu, P. Hormonal regulators of flowering and fruit development: Effect of GA and ethereal on fruit setting and development of Momordica charinata L. Biol. Plant 1992, 34, 63–70. [Google Scholar]
- Lee, Z.; Kim, S.; Choi, S.J.; Joung, E.; Kwon, M.; Park, H.J.; Shim, J.S. Regulation of flowering time by environmental factors in plants. Plants 2023, 12, 3680. [Google Scholar] [CrossRef]
- Dong, X.; Li, Y.; Guan, Y.; Wang, S.; Luo, H.; Li, X.; Li, H.; Zhang, Z. Auxin-induced AUXIN RESPONSE FACTOR4 activates APETALA1 and FRUITFULL to promote flowering in woodland strawberry. Hortic. Res. 2021, 8, 115. [Google Scholar] [CrossRef]
- Liang, Y.; Jiang, C.; Liu, Y.; Gao, Y.; Lu, J.; Aiwaili, P.; Fei, Z.; Jiang, C.-Z.; Hong, B.; Ma, C. Auxin regulates sucrose transport to repress petal abscission in rose (Rosa hybrida). Plant Cell 2020, 32, 3485–3499. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Shan, N.; Liu, H.; Yao, X.; Wang, Z.; Bai, R.; Guo, Y.; Duan, Y.; Wang, C.; Sui, X. Transcriptional control of local auxin distribution by the CsDFB1-CsPHB module regulates floral organogenesis in cucumber. Proc. Natl. Acad. Sci. USA 2021, 118, e2023942118. [Google Scholar] [CrossRef]
- Liu, X.; Dinh, T.T.; Li, D.; Shi, B.; Li, Y.; Cao, X.; Guo, L.; Pan, Y.; Jiao, Y.; Chen, X. AUXIN RESPONSE FACTOR 3 integrates the functions of AGAMOUS and APETALA 2 in floral meristem determinacy. Plant J. 2014, 80, 629–641. [Google Scholar] [CrossRef]
- D’Aloia, M.; Bonhomme, D.; Bouché, F.; Tamseddak, K.; Ormenese, S.; Torti, S.; Coupland, G.; Périlleux, C. Cytokinin promotes flowering of Arabidopsis via transcriptional activation of the FT paralogue TSF. Plant J. 2011, 65, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Zieslin, N.; Mor, Y.; Khayat, E.; Levy, M. The use of cytokinins for promotion of flower production in roses. In Proceedings of the II Symposium on Growth Regulators in Floriculture 167, Skierniewice, Poland, 30 July–4 August 1984; pp. 433–434. [Google Scholar]
- Eshghi, S.; Tafazoli, E. Possible role of cytokinins in flower induction in strawberry. Am. J. Plant Physiol. 2007, 2, 167–174. [Google Scholar] [CrossRef]
- Qiu, Y.; Guan, S.C.; Wen, C.; Li, P.; Gao, Z.; Chen, X. Auxin and cytokinin coordinate the dormancy and outgrowth of axillary bud in strawberry runner. BMC Plant Biol. 2019, 19, 528. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Sharma, S.; Jose-Santhi, J.; Kalia, D.; Singh, R.K. Hormones regulate the flowering process in saffron differently depending on the developmental stage. Front. Plant Sci. 2023, 14, 1107172. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, C.; Yang, H.; Jiao, Y. Cytokinin pathway mediates APETALA1 function in the establishment of determinate floral meristems in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 6840–6845. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Suh, S.S.; Lee, H.; Choi, K.R.; Hong, C.B.; Paek, N.C.; Kim, S.G.; Lee, I. The SOC1 MADS-box gene integrates vernalization and gibberellin signals for flowering in Arabidopsis. Plant J. 2003, 35, 613–623. [Google Scholar] [CrossRef]
- Hu, J.; Liu, Y.; Tang, X.; Rao, H.; Ren, C.; Chen, J.; Wu, Q.; Jiang, Y.; Geng, F.; Pei, J. Transcriptome profiling of the flowering transition in saffron (Crocus sativus L.). Sci. Rep. 2020, 10, 9680. [Google Scholar]
- Martignago, D.; Siemiatkowska, B.; Lombardi, A.; Conti, L. Abscisic acid and flowering regulation: Many targets, different places. Int. J. Mol. Sci. 2020, 21, 9700. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, J.; Ohashi, Y.; Takahashi, R.; Nakai, K.; Takahashi, Y. DELLA degradation by gibberellin promotes flowering via GAF1-TPR-dependent repression of floral repressors in Arabidopsis. Plant Cell 2021, 33, 2258–2272. [Google Scholar]
- Zhang, S.; Gottschalk, C.; van Nocker, S. Genetic mechanisms in the repression of flowering by gibberellins in apple (Malus x domestica Borkh.). BMC Genom. 2019, 20, 747. [Google Scholar]
- King, R.W.; Hisamatsu, T.; Goldschmidt, E.E.; Blundell, C. The nature of floral signals in Arabidopsis. I. Photosynthesis and a far-red photoresponse independently regulate flowering by increasing expression of FLOWERING LOCUS T (FT). J. Exp. Bot. 2008, 59, 3811–3820. [Google Scholar]
- Lawson, R.H. Economic importance and trends in ornamental horticulture. In Proceedings of the IX International Symposium on Virus Diseases of Ornamental Plants 432, Herzliya, Israel, 17–22 March 1996; pp. 226–237. [Google Scholar]
- Noman, A.; Aqeel, M.; Deng, J.; Khalid, N.; Sanaullah, T.; Shuilin, H. Biotechnological advancements for improving floral attributes in ornamental plants. Front. Plant Sci. 2017, 8, 530. [Google Scholar]
- Magray, M.M. Plant growth regulators and their role in horticultural crop production and development. Res. Manag. Hortic. Crops 2021, 1, 31–51. [Google Scholar]
- Nambara, E.; Yan, D.; Wen, J.; Sharma, A.; Nguyen, F.; Yan, A.; Uruma, K.; Yano, K. Plant Hormones: Gene Family Organization and Homolog Interactions of Genes for Gibberellin Metabolism and Signaling in Allotetraploid Brassica napus. In Plant Omics: Advances in Big Data Biology; CABI GB: Wallingford, UK, 2022; pp. 151–171. [Google Scholar]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. Vitr. Cell Dev. Biol.-Plant 1996, 32, 272–289. [Google Scholar] [CrossRef]
- Seaton, K.; Bettin, A.; Grüneberg, H. New ornamental plants for horticulture. In Horticulture: Plants for People and Places, Volume 1: Production Horticulture; Springer: Berlin/Heidelberg, Germany, 2014; pp. 435–463. [Google Scholar]
- Serek, M.; Woltering, E.; Sisler, E.; Frello, S.; Sriskandarajah, S. Controlling ethylene responses in flowers at the receptor level. Biotechnol. Adv. 2006, 24, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Ryu, H.; Cho, Y.-G. Plant hormones in salt stress tolerance. J. Plant Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef] [PubMed]
- Mehbub, H.; Akter, A.; Akter, M.A.; Mandal, M.S.H.; Hoque, M.A.; Tuleja, M.; Mehraj, H. Tissue culture in ornamentals: Cultivation factors, propagation techniques, and its application. Plants 2022, 11, 3208. [Google Scholar] [CrossRef]
- Kuligowska, K.; Lütken, H.; Müller, R. Towards development of new ornamental plants: Status and progress in wide hybridization. Planta 2016, 244, 1–17. [Google Scholar] [CrossRef]
- Alsoufi, A.S.; Ahmed, Z.S.; Salim, A.M. The efficiency of interaction between cytokines and Auxins in Micropropagation of Chrysanthemum plant (Chrysanthemum indicum L.). In IOP Conference Series: Earth and Environmental Science; IOP Publishing Ltd.: Bristol, UK, 2021; p. 012048. [Google Scholar]
- Reid, M.S.; Wu, M.-J. Ethylene in flower development and senescence. In The Plant Hormone Ethylene; CRC Press: Boca Raton, FL, USA, 2018; pp. 215–234. [Google Scholar]
- Castro-Camba, R.; Sánchez, C.; Vidal, N.; Vielba, J.M. Plant development and crop yield: The role of gibberellins. Plants 2022, 11, 2650. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Peñuelas, J. Gardening and urban landscaping: Significant players in global change. Trends Plant Sci. 2008, 13, 60–65. [Google Scholar] [CrossRef]
- El Malahi, S.; Sbah, N.; Zim, J.; Ennami, M.; Zakri, B.; Mokhtari, W.; Taimourya, H.; Mokhtari, M.; Hassani, L.M.I. Enhancing rooting efficiency and nutrient uptake in Rosa damascena Mill. cuttings: Insights into auxin and cutting type optimization. Plant Sci. Today 2024, 11, 119–131. [Google Scholar] [CrossRef]
- Anjum, S.; Mirza, U.; Shafi, N.; Parray, J.A. Plant–microbe interactions: Perspectives in promoting plant health. In Microbiome Drivers of Ecosystem Function; Elsevier: Amsterdam, The Netherlands, 2024; pp. 79–90. [Google Scholar]
- Finkel, O.M.; Castrillo, G.; Paredes, S.H.; González, I.S.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [PubMed]
- Singh, M.; Singh, D.; Gupta, A.; Pandey, K.D.; Singh, P.; Kumar, A. Plant growth promoting rhizobacteria: Application in biofertilizers and biocontrol of phytopathogens. In PGPR Amelioration in Sustainable Agriculture; Elsevier: Amsterdam, The Netherlands, 2019; pp. 41–66. [Google Scholar]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar]
- Poupin, M.J.; Timmermann, T.; Vega, A.; Zuñiga, A.; González, B. Effects of the plant growth-promoting bacterium Burkholderia phytofirmans PsJN throughout the life cycle of Arabidopsis thaliana. PLoS ONE 2013, 8, e69435. [Google Scholar]
- Flores, A.C.; Luna, A.A.E.; Portugal, V.O. Yield and quality enhancement of marigold flowers by inoculation with Bacillus subtilis and Glomus fasciculatum. J. Sustain. Agric. 2007, 31, 21–31. [Google Scholar]
- de los Santos-Villalobos, S.; de Folter, S.; Délano-Frier, J.P.; Gómez-Lim, M.A.; Guzmán-Ortiz, D.A.; Pena-Cabriales, J.J. Growth promotion and flowering induction in mango (Mangifera indica L. cv “Ataulfo”) trees by Burkholderia and Rhizobium Inoculation: Morphometric, biochemical, and molecular events. J. Plant Growth Regul. 2013, 32, 615–627. [Google Scholar]
- Lyons, R.; Rusu, A.; Stiller, J.; Powell, J.; Manners, J.M.; Kazan, K. Investigating the association between flowering time and defense in the Arabidopsis thaliana-Fusarium oxysporum interaction. PLoS ONE 2015, 10, e0127699. [Google Scholar]
- Das, A.; Kamal, S.; Shakil, N.A.; Sherameti, I.; Oelmüller, R.; Dua, M.; Tuteja, N.; Johri, A.K.; Varma, A. The root endophyte fungus Piriformospora indica leads to early flowering, higher biomass and altered secondary metabolites of the medicinal plant, Coleus forskohlii. Plant Signal. Behav. 2012, 7, 103–112. [Google Scholar] [PubMed]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef]
- Xing, L.-J.; Wei, L.I.; Zhai, Y.-L.; Hu, X.-Y.; Guo, S.-X. Arbuscular mycorrhizal fungi promote early flowering and prolong flowering in Antirrhinum majus L. by regulating endogenous hormone balance under field-planting conditions. Not. Bot. Horti Agrobot. 2022, 50, 12503. [Google Scholar]
- Toffoli, L.M.; Martínez-Zamora, M.G.; Medrano, N.N.; Fontana, C.A.; Lovaisa, N.C.; Delaporte-Quintana, P.; Elias, J.M.; Salazar, S.M.; Pedraza, R.O. Natural occurrence of Azospirillum brasilense in petunia with capacity to improve plant growth and flowering. J. Basic Microbiol. 2021, 61, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Zulueta-Rodriguez, R.; Cordoba-Matson, M.V.; Hernandez-Montiel, L.G.; Murillo-Amador, B.; Rueda-Puente, E.; Lara, L. Effect of Pseudomonas putida on growth and anthocyanin pigment in two poinsettia (Euphorbia pulcherrima) cultivars. Sci. World J. 2014, 2014, 810192. [Google Scholar] [CrossRef] [PubMed]
- Vaingankar, J.D.; Rodrigues, B. Effect of arbuscular mycorrhizal (AM) inoculation on growth and flowering in Crossandra infundibuliformis (L.) Nees. J. Plant Nutr. 2015, 38, 1478–1488. [Google Scholar] [CrossRef]
- Yanti, Y.; Warnita, W.; Reflin, R.; Nasution, C.R. Characterizations of endophytic Bacillus strains from tomato roots as growth promoter and biocontrol of Ralstonia solanacearum. Biodiversitas J. Biol. Divers. 2018, 19, 906–911. [Google Scholar] [CrossRef]
- Goicoechea; Aguirreolea; Cenoz, G.-M. Gas exchange and flowering in Verticillium-wilted pepper plants. J. Phytopathol. 2001, 149, 281–286. [Google Scholar] [CrossRef]
- Nordstedt, N.P.; Chapin, L.J.; Taylor, C.G.; Jones, M.L. Identification of Pseudomonas spp. that increase ornamental crop quality during abiotic stress. Front. Plant Sci. 2020, 10, 1754. [Google Scholar] [CrossRef]
- Sharaf-Eldin, M.; Elkholy, S.; Fernández, J.-A.; Junge, H.; Cheetham, R.; Guardiola, J.; Weathers, P. Bacillus subtilis FZB24® affects flower quantity and quality of saffron (Crocus sativus). Planta Med. 2008, 74, 1316–1320. [Google Scholar] [CrossRef]
- Ousley, M.; Lynch, J.; Whipps, J. The effects of addition of Trichoderma inocula on flowering and shoot growth of bedding plants. Sci. Hortic. 1994, 59, 147–155. [Google Scholar] [CrossRef]
- Cui, Z.; Huntley, R.B.; Zeng, Q.; Steven, B. Temporal and spatial dynamics in the apple flower microbiome in the presence of the phytopathogen Erwinia amylovora. ISME J. 2021, 15, 318–329. [Google Scholar] [CrossRef]
- Christensen, S.M.; Munkres, I.; Vannette, R.L. Nectar bacteria stimulate pollen germination and bursting to enhance microbial fitness. Curr. Biol. 2021, 31, 4373–4380.e6. [Google Scholar] [CrossRef]
- Gätjens-Boniche, O.; Jiménez-Madrigal, J.P.; Whetten, R.W.; Valenzuela-Diaz, S.; Alemán-Gutiérrez, A.; Hanson, P.E.; Pinto-Tomás, A.A. Microbiome and plant cell transformation trigger insect gall induction in cassava. Front. Plant Sci. 2023, 14, 1237966. [Google Scholar]
- Crowley-Gall, A.; Rering, C.C.; Rudolph, A.B.; Vannette, R.L.; Beck, J.J. Volatile microbial semiochemicals and insect perception at flowers. Curr. Opin. Insect Sci. 2021, 44, 23–34. [Google Scholar] [PubMed]
- Dombrowski, N.; Schlaeppi, K.; Agler, M.T.; Hacquard, S.; Kemen, E.; Garrido-Oter, R.; Wunder, J.; Coupland, G.; Schulze-Lefert, P. Root microbiota dynamics of perennial Arabis alpina are dependent on soil residence time but independent of flowering time. ISME J. 2017, 11, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.R.; Lundberg, D.S.; Coleman-Derr, D.; Tringe, S.G.; Dangl, J.L.; Mitchell-Olds, T. Natural soil microbes alter flowering phenology and the intensity of selection on flowering time in a wild Arabidopsis relative. Ecol. Lett. 2014, 17, 717–726. [Google Scholar]
- Yoo, S.K.; Chung, K.S.; Kim, J.; Lee, J.H.; Hong, S.M.; Yoo, S.J.; Yoo, S.Y.; Lee, J.S.; Ahn, J.H. Constans activates suppressor of overexpression of constans 1 through Flowering Locus T to promote flowering in Arabidopsis. Plant Physiol. 2005, 139, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annu. Rev. Plant Biol. 2008, 59, 573–594. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Ying, H.; Helliwell, C.A.; Taylor, J.M.; Peacock, W.J.; Dennis, E.S. FLOWERING LOCUS C (FLC) regulates development pathways throughout the life cycle of Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 6680–6685. [Google Scholar] [CrossRef]
- Peer, L.A.; Bhat, M.Y.; Ahmad, N.; Mir, B.A. Floral induction pathways: Decision making and determination in plants to flower-A comprehensive review. J. Appl. Biol. Biotechnol 2021, 9, 7–17. [Google Scholar]
- Izawa, T. What is going on with the hormonal control of flowering in plants? Plant J. 2021, 105, 431–445. [Google Scholar] [CrossRef]
- Wang, P.; Li, Y.; Liu, Z.; Li, X.; Wang, Y.; Liu, W.; Li, X.; Hu, J.; Zhu, W.; Wang, C. Reciprocal regulation of flower induction by ELF3α and ELF3β generated via alternative promoter usage. Plant Cell 2023, 35, 2095–2113. [Google Scholar] [CrossRef]
- Eichmann, R.; Richards, L.; Schäfer, P. Hormones as go-betweens in plant microbiome assembly. Plant J. 2021, 105, 518–541. [Google Scholar] [CrossRef]
- Shah, A.; Tyagi, S.; Saratale, G.D.; Guzik, U.; Hu, A.; Sreevathsa, R.; Reddy, V.D.; Rai, V.; Mulla, S.I. A comprehensive review on the influence of light on signaling cross-talk and molecular communication against phyto-microbiome interactions. Crit. Rev. Biotechnol. 2021, 41, 370–393. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Datta, S.; Ramamurthy, P.C.; Singh, J. Molecular mechanism and signaling pathways interplay between plant hormones during plant-microbe crosstalk. In Microbial Management of Plant Stresses; Elsevier: Amsterdam, The Netherlands, 2021; pp. 93–105. [Google Scholar]
- Darras, A.I. Implementation of sustainable practices to ornamental plant cultivation worldwide: A critical review. Agronomy 2020, 10, 1570. [Google Scholar] [CrossRef]
- Atal, H.L.; Srilakshmi, D.; Debbarma, K.; Jena, L.; Ichancha, M. A Review on Breeding in Ornamental Crops for Abiotic Stress Tolerance. Int. J. Plant Soil Sci. 2022, 34, 134–138. [Google Scholar] [CrossRef]
- Chandler, S.F.; Tribe, D. Modern techniques for plant breeding in ornamentals. In Floriculture and Ornamental Plants; Springer: Berlin/Heidelberg, Germany, 2022; pp. 523–555. [Google Scholar]
- Roberts, W.R.; Roalson, E.H. Co-expression clustering across flower development identifies modules for diverse floral forms in Achimenes (Gesneriaceae). PeerJ 2020, 8, e8778. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Song, A.; Zhang, X.; Li, S.; Su, J.; Xia, W.; Zhao, K.; Zhao, W.; Guan, Y.; Fang, W. The core regulatory networks and hub genes regulating flower development in Chrysanthemum morifolium. Plant Mol. Biol. 2020, 103, 669–688. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Kang, X.; Wang, M.; Haider, W.; Price, W.B.; Hajek, B.; Hanzawa, Y. Transcriptome-enabled network inference revealed the GmCOL1 feed-forward loop and its roles in photoperiodic flowering of soybean. Front. Plant Sci. 2019, 10, 1221. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbe | Type | Plant Host | Effect on Flowering | References |
|---|---|---|---|---|
| Burkholderia phytofirmans | Bacteria | Arabidopsis thaliana | Promotes flowering time | [148] |
| Bacillus subtilis | Bacteria | Marigold (Tagetes) | Flowering color | [149] |
| Rhizobium spp. | Fungi | Mangifera indica L. cv | Flowering number | [150] |
| Fusarium oxysporum | Fungi | Arabidopsis thaliana | Flowering timing | [151] |
| Piriformospora indica | Fungi | Coleus forskohlii | Promotes early flowering | [152] |
| Trichoderma harzianum | Fungi | Chrysanthemum | Flowering number | [153] |
| Arbuscular mycorrhizal fungi | Fungi | Antirrhinum majus L. | Promote flowering | [154] |
| Azospirillum brasilense | Bacteria | Petunia | Promotes flowering number | [155] |
| Pseudomonas putida | Bacteria | Euphorbia pulcherrima | Promotes flowering number | [156] |
| Glomus intraradices | Mycorrhizal fungi | Crossandra infundibuliformis L. | Promote flowering weight and number | [157] |
| Bacillus spp. | Bacteria | Solanum lycopersicum | Flowering time | [158] |
| B. acidiceler, B. subtilis and B. pumilus | Bacteria | Rosa hybrida L. | Flowering diameter | [159] |
| Pseudomonas spp. | Bacteria | Petunia plants | Flower number | [160] |
| Bacillus subtilis FZB24 | Bacteria | Crocus sativus | Flowers per corm | [161] |
| Trichoderma inocula | Fungi | Petunia plant | Promotes flowering | [162] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aizaz, M.; Lubna; Hashmi, S.S.; Khan, M.A.; Jan, R.; Bilal, S.; Kim, K.-M.; Al-Harrasi, A.; Asaf, S. Unraveling the Complexities of Flowering in Ornamental Plants: The Interplay of Genetics, Hormonal Networks, and Microbiome. Plants 2025, 14, 1131. https://doi.org/10.3390/plants14071131
Aizaz M, Lubna, Hashmi SS, Khan MA, Jan R, Bilal S, Kim K-M, Al-Harrasi A, Asaf S. Unraveling the Complexities of Flowering in Ornamental Plants: The Interplay of Genetics, Hormonal Networks, and Microbiome. Plants. 2025; 14(7):1131. https://doi.org/10.3390/plants14071131
Chicago/Turabian StyleAizaz, Muhammad, Lubna, Syed Salman Hashmi, Muhammad Aaqil Khan, Rahmatullah Jan, Saqib Bilal, Kyung-Min Kim, Ahmed Al-Harrasi, and Sajjad Asaf. 2025. "Unraveling the Complexities of Flowering in Ornamental Plants: The Interplay of Genetics, Hormonal Networks, and Microbiome" Plants 14, no. 7: 1131. https://doi.org/10.3390/plants14071131
APA StyleAizaz, M., Lubna, Hashmi, S. S., Khan, M. A., Jan, R., Bilal, S., Kim, K.-M., Al-Harrasi, A., & Asaf, S. (2025). Unraveling the Complexities of Flowering in Ornamental Plants: The Interplay of Genetics, Hormonal Networks, and Microbiome. Plants, 14(7), 1131. https://doi.org/10.3390/plants14071131