
Phenotypic Diversity and Abiotic Stress Tolerance Among Vicia ervilia (L.) Willd. Accessions

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Measurements of Traits
2.3. Biochemical Analyses of Grain Quality
2.4. Germination Tests in Control and Stressful Conditions
2.5. Data Analysis
3. Results
3.1. Morphological Evaluation
3.2. Phenotypic Variation in the Studied Parameters
3.3. Correlation Between the Studied Economic, Biological, and Biochemical Traits
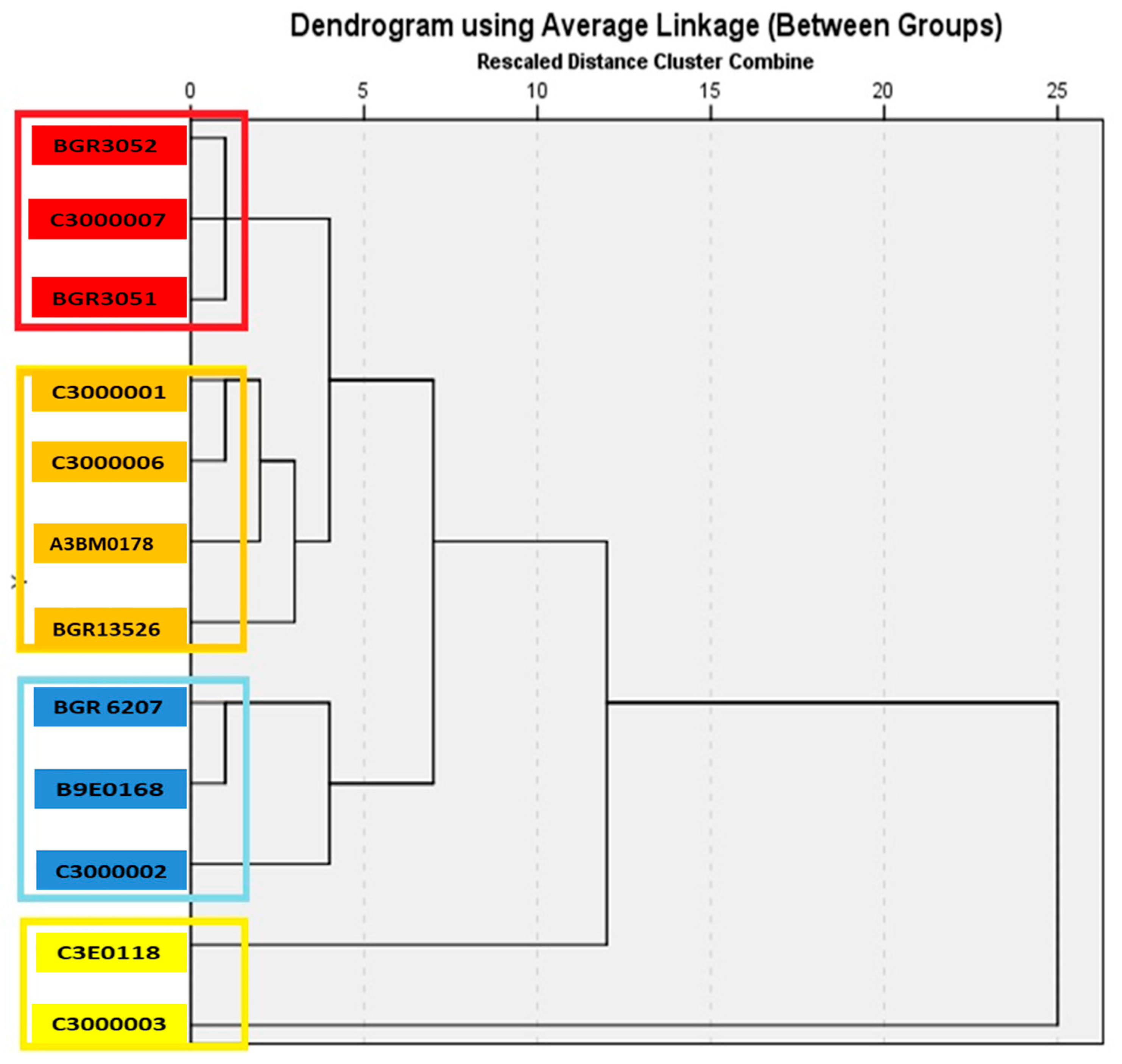
3.4. Hierarchical Cluster Analysis
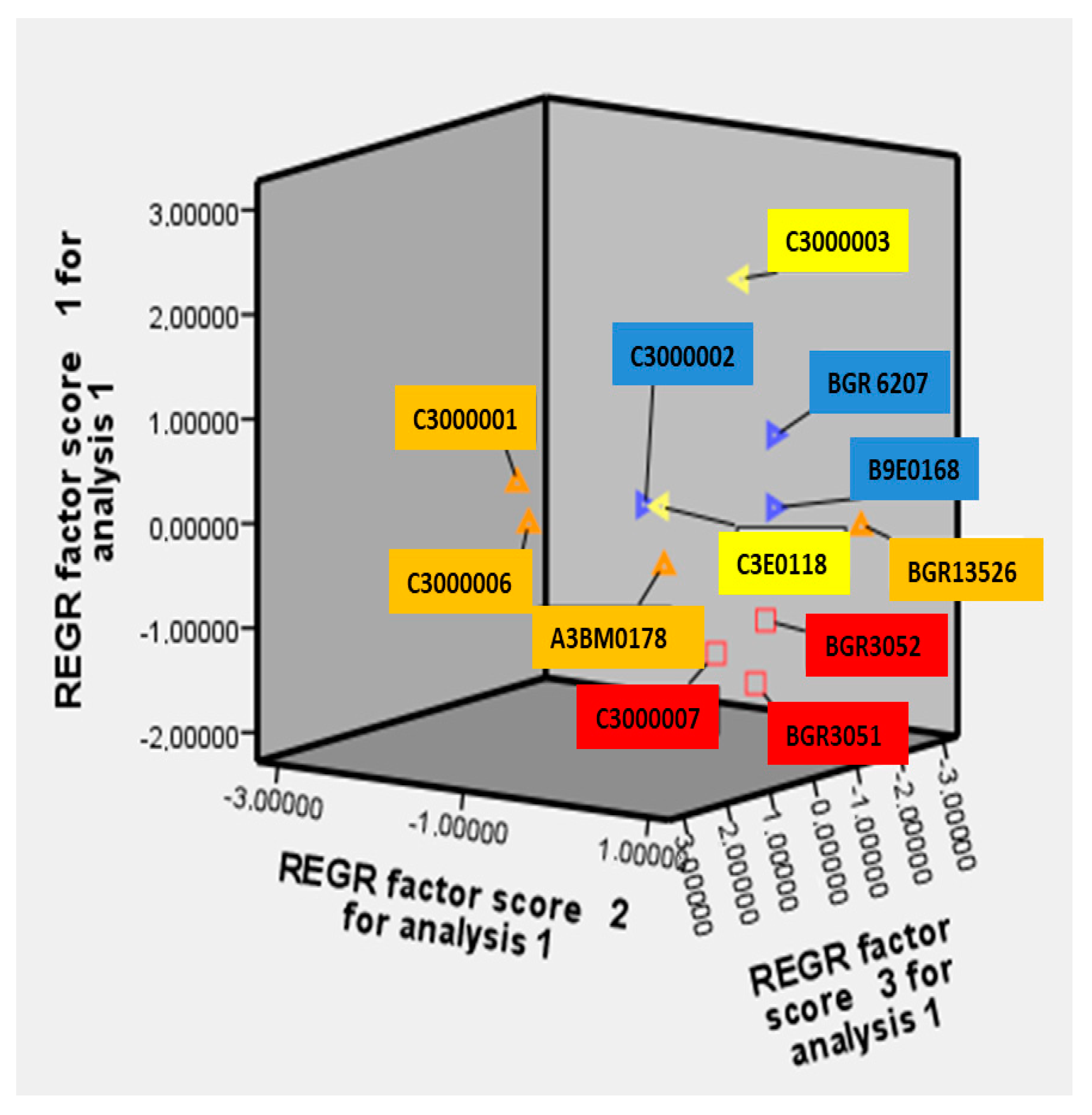
3.5. Principal Component Analysis (PCA)
3.6. Path Analysis of Productivity and Economic, Biological, and Biochemical Traits
3.7. Relative Stress Tolerance at Germination Stage
4. Discussion
4.1. Phenotypic Variations Among the Studied Bitter Vetch Accessions
4.2. Yield Structure of the Studied Bitter Vetch Accessions
4.3. Stress Tolerance of the Studied Bitter Vetch Accessions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Champ, M.M.J. Non-nutrient bioactive substances of pulses. Brit. J. Nutr. 2002, 88 (Suppl. S3), 307–319. [Google Scholar] [CrossRef]
- Alkin, R.; Goyder, D.J.; Bisby, F.A.; White, R.J. Names and Synonyms of Species and Subspecies in the Viceae: Issue 3; University of Southampton: Hampshire, UK, 1986. [Google Scholar]
- Maxted, N. An Ecogeographical Study of Vicia subgenus Vicia. In Systematic and Ecogeographic Studies on Crop Genepools 8; Plant Genetic Resources Institute: Rome, Italy, 1995. [Google Scholar]
- Saoub, M.H.; Akash, W.M. Variations among two vetch landrace species in Jordan. J. Food Agricult. Environ. 2012, 10, 763–767. [Google Scholar]
- Christiansen, S.; Bounejmate, M.; Bahhady, F.; Thomson, E.; Mawlawi, B.; Singh, M. On-farm trials with forage legume-barley compared with fallow-barley rotation and continuous barley in north-west Syria. Exp. Agric. 2000, 36, 195–204. [Google Scholar] [CrossRef]
- Jones, M.J.; Singh, M. Long-term yield patterns in barley-based cropping systems in Northern Syria. 2. The role of feed legumes. J. Agric. Sci. 2000, 135, 237–249. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World; Oxford University Press: Oxford, UK, 2012; 264p, ISBN 9780199549061. [Google Scholar]
- Okba, M.M.; Abdel Jaleel, G.A.; Yousif, M.F.; El Deeb, K.S.; Soliman, F.M. Vicia ervilia L. seeds newly explored biological activities. Cogent Biol. 2017, 3, 1299612. [Google Scholar] [CrossRef]
- Vioque, J.; Giron-Calle, J.; Torres-Salas, V.; Elamine, Y.; Alaiz, M. Characterizion of Vicia ervilia (bitter vetch) seed protein, free amino acide, and polyphenols. J. Food Biochem. 2020, 44, e13271. [Google Scholar] [CrossRef]
- Abd El-Moneim, A.M.; Ryan, J. Forage legumes for dryland agriculture in Central and West Asia and North Africa. In Challenges and Strategies for Dryland Agriculture; Rao, S.C., Ryan, J., Eds.; Book series CSSA Special Publications, 32; Crop Science Society of America. American Society of Agronomy: Madison, WI, USA, 2004; pp. 243–256. [Google Scholar] [CrossRef]
- Haddad, S.G. Bitter vetch grains as a substitute for soybean meal for growing lambs. Livest. Sci. 2006, 99, 221–225. [Google Scholar] [CrossRef]
- Abdullah, Y.; Muwalla, M.M.; Qudsieh, R.I.; Titi, H.H. Effect of bitter vetch (Vicia ervilia) seeds as a replacement protein source of soybean meal on performance and carcass characteristics of finishing Awassi lambs. Trop. Anim. Health Prod. 2010, 42, 293–300. [Google Scholar] [CrossRef]
- Sadeghi, G.H.; Mohammadi, L.; Ibrahim, S.A.; Gruber, K.J. Use of bitter vetch (Vicia ervilia) as feed ingredient for poultry. World Poult. Sci. J. 2009, 65, 51–63. [Google Scholar] [CrossRef]
- Petkova, Z.Y.; Antova, G.A.; Angelova-Romova, M.Y.; Petrova, A.; Stoyanova, M.; Petrova, S.; Stoyanova, A. Bitter vetch seeds (Vicia ervilia L.)—A valuable source of nutrients. Bulg. Chem. Commun. 2020, 52, 12–15. [Google Scholar] [CrossRef]
- Russi, L.; Acuti, G.; Trabalza-Marinucci, M.; Porta, R.; Rubini, A.; Damiani, F.; Cristiani, S.; Dal Bosco, A.; Martuscelli, G.E.; Bellucci, M.; et al. Genetic characterisation and agronomic and nutritional value of bitter vetch (Vicia ervilia), an under-utilised species suitable for low-input farming systems. Crop Past. Sci. 2019, 70, 606–614. [Google Scholar] [CrossRef]
- Parisi, G.; Tulli, F.; Fortina, R.; Marino, R.; Bani, P.; Dalle Zotte, A.; De Angelis, A.; Piccolo, G.; Pinotti, L.; Schiavone, A.; et al. Protein hunger of the feed sector: The alternatives offered by the plant world. Ital. J. Anim. Sci. 2020, 19, 1204–1225. [Google Scholar] [CrossRef]
- Boukrouh, S.; Noutfia, A.; Moula, N.; Avril, C.; Louvieaux, J.; Hornick, J.L.; Chentouf, M.; Cabaraux, J.F. Characterisation of bitter vetch (Vicia ervilia (L.) Willd) ecotypes: An ancient and promising legume. Exp. Agricult. 2024, 60, e19. [Google Scholar] [CrossRef]
- Salehi, B.; Abu-Reidah, I.M.; Sharopov, F.; Karazhan, N.; Sharifi-Rad, J.; Akram, M.; Daniyal, M.; Khan, F.S.; Abbaass, W.; Zainab, R.; et al. Vicia plants—A comprehensive review on chemical composition and phytopharmacology. Phytother. Res. 2021, 35, 790–809. [Google Scholar] [CrossRef]
- Basheer-Salimia, R.; Aloweidat, M.Y.; Al-Salimiya, M.A.; Hamdan, Y.A.S.; Sayara, T.A.S. Comparative study of five legume species under drought conditions. Legume Res. 2021, 44, 712–717. [Google Scholar] [CrossRef]
- Abdi, H.; Taheri, M.T.; Alizadeh, K.; Shahbazi, S. Investigating the Phenotypic Diversity of Vetch (Vicia L.) Genotypes from Different Species Under the Rain-fed Conditions of Maragheh City. Taxon. Biosystemat. 2023, 15, 19–34. Available online: https://tbj.ui.ac.ir/article_27554.html (accessed on 13 February 2025). (In Persian).
- Çöçü, S.; Uzun, O. Germination, seedling growth and ion accumulation of bitter vetch (Vicia ervilia (L.) Willd.) lines under NaCl stress. Afr. J. Biotechnol. 2011, 10, 15869–15874. [Google Scholar] [CrossRef]
- Firincioǧlu, H.K. A comparison of six vetches (Vicia spp.) for developmental rate, herbage yield and grain yield in semi-arid central Turkey. Grass Forage Sci. 2014, 69, 303–314. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Anastasi, U.; Santonoceto, C.; Maggio, A. Effect of PEG-induced drought stress on seed germination of four lentil genotypes. J. Plant Interact. 2014, 9, 354–363. [Google Scholar] [CrossRef]
- Abbasi, A.; Mohammadi Nargesi, B.; Keshavarz Niya, R.; Ghorbanpour, E. The study of genetic variation of common vetch (Vicia sativa L.) based on morphological traits under normal and stress conditions. Iran. J. Field Crop Sci. 2014, 44, 359–370. (In Persian) [Google Scholar] [CrossRef]
- Benidire, L.; Daoui, K.; Fatemi, Z.A.; Bouarab, L.; Oufdou, K. Effect of salt stress on germination and seedling growth of Vicia faba L. J. Mat. Environ. Sci. 2015, 6, 840–851. (In French) [Google Scholar]
- Keatinge, J.D.H.; Ali, A.; Roidar Khan, B.; Abd El Moneim, A.M.; Ahmad, S. Germplasm Evaluation of Annual Sown Forage Legumes under Environmental Conditions Marginal for Crop Growth in the Highlands of West Asia. J. Agron. Crop Sci. 1991, 166, 48–57. [Google Scholar] [CrossRef]
- El Fatehi, S.; Bena, G.; Filali-Maltouf, A.; Ater, M. Variation in yield component, phenology and morphological traits among Moroccan bitter vetch landraces Vicia ervilia L. Willd. Afr. J. Agric. Res. 2014, 9, 1801–1809. [Google Scholar] [CrossRef]
- Abbasi, M.R.; Vaezi, S.; Baghaie, N. Genetic diversity of bitter vetch (Vicia ervilia) of the National Plant Gene Bank of Iran based on agro-morphological traits. Iran. J. Rangelands Forests Plant Breed. Genet. Res. 2007, 15, 113–128. (In Persian) [Google Scholar] [CrossRef]
- Livanios, I.; Lazaridi, E.; Bebeli, P.J. Assessment of phenotypic diversity in bitter vetch (Vicia ervilia (L.) Willd.) populations. Genet. Resour. Crop. Evol. 2017, 65, 355–371. [Google Scholar] [CrossRef]
- International Union for the Protection of New Varieties of Plants. Common Vetch (Vicia sativa L.) Guidelines for the Conduct of Tests for Distinctness Uniformity and Stability; International Union for the Protection of New Varieties of Plants: Geneva, Switzerland, 2011. [Google Scholar]
- López-Hidalgo, C.; Meijón, M.; Lamelas, L.; Valledor, L. The rainbow protocol: A sequential method for quantifying pigments, sugars, free amino acids, phenolics, flavonoids and MDA from a small amount of sample. Plant Cell Environ. 2021, 44, 1977–1986. [Google Scholar]
- Hansen, J.; Møller, I.B. Percolation of starch and soluble carbohydrates from plant tissue for quantitative determination with anthrone. Analyt. Biochem. 1975, 68, 87–94. [Google Scholar] [CrossRef]
- Moore, S.; Stein, W.H. A modified ninhydrin reagent for the photometric determination of amino acids and related compounds. J. Biol. Chem. 1954, 211, 907–913. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Kakade, M.L.; Rackis, J.J.; McGhee, J.E.; Puski, G. Determination of trypsin inhibitor activity of soy products: A collaborative analysis of an improved procedure. Cereal Chem. 1974, 51, 376–382. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantification of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Analyt. Biochem. 1976, 72, 248–254. [Google Scholar] [PubMed]
- Lidanski, T. Statistical Methods in Biology and Agriculture; Zemizdat: Sofia, Bulgaria, 1988; 160p. (In Bulgarian) [Google Scholar]
- Arora, J.S. Chapter 20—Additional Topics on Optimum Design. In Introduction to Optimum Design, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 731–784. ISBN 9780123813756. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Amer. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Rachovska, G.; Dimova, D. Evaluation of selection effectiveness for productivity in mutant populations of common bread wheat. Bulg. J. Agric. Sci. 2002, 8, 473–478. [Google Scholar]
- Williams, W.A.; Jones, M.B.; Demment, M.W. A concise table for path analysis statistics. Agron. J. 1990, 82, 1022–1024. [Google Scholar] [CrossRef]
- Cattel, R.B. Factor analysis: An introduction to essentials. I. The purpose and underlying models. Biometrics 1965, 21, 190–215. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, Version 19.0; IBM Corp: Armonk, NY, USA, 2009. [Google Scholar]
- Larbi, A.; Abd El Moneim, A.M.; Nakkoul, H.; Jammal, B.; Hassan, S. Intra-species variations in yield and quality determinants in Vicia species: 1. bitter vetch (Vicia ervilia L.). Anim. Feed. Sci. Technol. 2011, 65, 278–287. [Google Scholar] [CrossRef]
- Mihov, M.; Atanasova, D.; Mekhanjiev, A. Advances in productivity and stress resistance selection in chickpea (Cicer arietinum L.). Sci. Work. Nat. Acad. Sci. 2005, 3, 150–157. [Google Scholar]
- Abd El-Moneim, A.M. Agronomic potential of three vetches (Vicia spp.) under rainfed conditions. J. Agron. Crop Sci. 1993, 170, 113–120. [Google Scholar] [CrossRef]
- Seydoșoğlu, S.; Saruhan, V.; Kokten, K. Researches on determination yield and yield components of some bitter vetch (Vicia ervilia (L.) Willd.) genotypes in ecological conditions of Diyarbakır. Gaziosmanpaşa Üniversitesi Ziraat Fakültesi Derg. 2015, 32, 107–115. [Google Scholar]
- Turk, M. Effects of sowing rate and irrigation on dry biomass and grain yield of bitter vetch (Vicia ervilia) and narbon vetch (Vicia narbonensis). Indian J. Agic. Sci. 1999, 69, 438–443. Available online: https://epubs.icar.org.in/index.php/IJAgS/article/view/26807 (accessed on 13 February 2025).
- Ayan, I.; Acar, Z.; Basaran, U.; Asci, O.O.; Mut, H. Determination of forage and grain yields of some Vicia ervilia L. lines in Samsun ecological conditions. OMÜ Ziraat Fakültesi Derg. 2006, 21, 318–322. [Google Scholar]
- ICARDA. IRAN/ICARDA collaboration, Highlights of Achievements 2005–2006; ICARDA and Agricultural Research and Education Organization, Ministry of Jihad-e-eAgriculture, Islamic Republic of Iran (AEERO), Project Report; International Center for Agricultural Research in the Dry Areas: Aleppo, Syria, 2008; pp. 47–49. [Google Scholar]
- Mekonnen, F.; Mekbib, F.; Kumar, S.; Ahmed, S.; Sharma, T.R. Agromorphological traits variability of the ethiopian lentil and exotic genotypes. Adv. Agric. 2014, 2014, 870864. [Google Scholar] [CrossRef]
- Fotovat, R.; Moghadam, M.; Bozorgzadeh, M. Study of genetic diversity of bitter vetch landraces. In Proceedings of the 5th National Iranian Crop Science Congress, University of Tehran, Tehran, Iran, 31 August–4 September 1998; pp. 67–68. [Google Scholar]
- Hassanpour, F.; Sahhaf, R.S. Genetic variation in some Iranian bitter vetch (Vicia ervilia L.) landraces based on agronomic-morphological traits for use in breeding program in Rafsanjan. Genet. Resour. Crop. Evol. 2020, 67, 2087–2100. [Google Scholar] [CrossRef]
- Sachanski, S. Study on the Biological Requirements and Economic Qualities of Some Annual Leguminous Fodder Plants. Ph.D. Thesis, Publication of the NGO for Cattle and Sheep Breeding, Sofia, Bulgaria, Institute of Forages, Pleven, Bulgaria, 1983. [Google Scholar]
- Ganeva, D.; Angelova, S.; Kostov, K.; Matsov, B. Results of studying a collection of overwintering varieties and forms of fodder pea. Plant Sci. 1985, 22, 46–53. [Google Scholar]
- Dewey, D.I.; Lu, K.H. A Correlation and Path-Coefficient Analysis of Components of Crested Wheatgrass Grain Production. Agron. J. 1959, 51, 515–518. [Google Scholar] [CrossRef]
- Yücel, D.; Anlarsal, A.E.; Yücel, C. Genetic variability, correlation and path analysis of yield and yield components in chickpea (Cicer arietinum L.). Turk. J. Agric. Forestry 2006, 30, 183–188. Available online: https://journals.tubitak.gov.tr/agriculture/vol30/iss3/2/ (accessed on 13 February 2025).
- Tu, B.; Zhang, Q.; Liu, X.; Yu, S.; Xu, N.; Liu, J.; Liu, C. Agronomic and pod traits in relation to pod shattering in cultivated soybeans. Czech J. Genet. Plant Breed. 2025, 61, 2. [Google Scholar] [CrossRef]
- Podolska, G. Plant lodging, effects, and control. In Encyclopedia of Agrophysics; Glin’ski, J., Horabik, J., Lipiec, J., Eds.; Part of the series encyclopedia of Earth Sciences Series; Springer: Berlin/Heidelberg, Germany, 2014; pp. 609–610. [Google Scholar] [CrossRef]
- Warkentin, T.; Smykal, P.; Coyne, C.J.; Weeden, N.; Domoney, C.; Bing, D.; Leonforte, T.; Zong, X.; Dixit, G.; Boros, L.; et al. Pea (Pisum sativum L.). In Handbook of Plant Breeding 10: Grain Legumes; De Ron, A., Ed.; Springer Science and Business Media: New York, NY, USA, 2015; pp. 37–83. [Google Scholar] [CrossRef]
- Kujur, A.; Upadhyaya, H.D.; Bajaj, D.; Gowda, C.L.L.; Sharma, S.; Tyagi, A.K.; Parida, S.K. Identification of candidate genes and natural allelic variants for QTLs governing plant height in chickpea. Sci. Rep. 2016, 6, 27968. [Google Scholar] [CrossRef]
- Kuzbakova, M.; Khassanova, G.; Oshergina, I.; Ten, E.; Jatayev, S.; Yerzhebayeva, R.; Bulatova, K.; Khalbayeva, S.; Schramm, C.; Anderson, P.; et al. Height to first pod: A review of genetic and breeding approaches to improve combined harvesting in legume crops. Front. Plant Sci. 2022, 13, 948099. [Google Scholar] [CrossRef]
- Siwale, J.; Labuschagne, M.; Gerrano, A.S.; Mbuma, N.W. Phenotypic Diversity and Characterization of the Southern African Bambara Groundnut Germplasm Collection for Grain Yield and Yield Components. Agronomy 2022, 12, 1811. [Google Scholar] [CrossRef]
- Gil, J.; Cubero, J.I. Multivariate analysis of the Vicia sativa L. aggregate. Bot. J. Linn. Soc. 1993, 113, 389–400. [Google Scholar] [CrossRef]
- Terzopoulos, P.J.; Kaltsikes, P.J.; Bebeli, P.J. Collection, evaluation and classification of Greek populations of faba bean (Vicia faba L.). Genet. Resour. Crop. Evol. 2003, 50, 373–381. [Google Scholar] [CrossRef]
- Firincioǧlu, H.K.; Ünal, S.; Erbektas¸, E.; Doǧruyol, S.U.L. Relationships between grain yield and yield components in common vetch (Vicia sativa ssp. sativa) populations sown in spring and autumn in central Turkey. Field Crop. Res. 2010, 116, 30–37. [Google Scholar] [CrossRef]
- Moghaddam, S.S.; Pourakbar, L.; Rahimi, A.; Jangjoo, F. Mitigation of salinity effects by salicylic acid priming on germination and physiological characteristics of bitter vetch (Vicia ervilia L.). Yuzuncu Yil Univ. J. Agricult. Sci. 2021, 31, 98–110. [Google Scholar] [CrossRef]
- Kka, N.M.; Sharif, K.N.; Mohammed, S.F.; AbdulHamid, K.A. Interactions between Halopriming and Hormopriming in Regulating the Vegetative Growth and Grain Quality of Vicia faba L. Basrah J. Agricult. Sci. 2022, 35, 1–23. [Google Scholar] [CrossRef]
- Yang, C.; Sun, R.; Lu, X.; Jin, T.; Peng, X.; Zhang, N.; Wang, J.; Wang, H.; Liu, W. Seed germination ecology of Vicia villosa Roth, a cover crop in orchards. Agronomy 2022, 12, 2488. [Google Scholar] [CrossRef]
- Perissé, P.; Arias, C.V.; Nogués, M.S. Germination, hydrotime and isotopic analysis of Vicia villosa Roth. under water and salinity stress. AgriScientia 2021, 38, 89–101. [Google Scholar] [CrossRef]
- Hojjat, S.S.; Hornyak, L.G. TiO2 Nanoparticle Improve germination and seedling parameters and enhance tolerance of bitter vetch (Vicia ervilia L.) plants under salinity and drought stress. Nanotechnol. Russ. 2022, 17, 411–419. [Google Scholar] [CrossRef]
- Aydinoğlu, B.; Shabani, A.; Safavi, S.M. Impact of priming on seed germination, seedling growth and gene expression in common vetch under salinity stress. Cell. Molec. Biol. 2019, 65, 18–24. [Google Scholar] [CrossRef]
- Pratap, V.; Sharma, Y.K. Impact of osmotic stress on seed germination and seedling growth in black gram (Phaseolus mungo). J. Environ. Biol. 2010, 31, 721–726. Available online: http://jeb.co.in/journal_issues/201009_sep10/paper_29.pdf (accessed on 13 February 2025).
- Tlahig, S.; Bellani, L.; Karmous, I.; Barbieri, F.; Loumerem, M.; Muccifora, S. Response to salinity in legume species: An insight on the effects of salt stress during seed germination and seedling growth. Chem. Biodivers. 2021, 18, e2000917. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Accession | Name | Biological Status | Origin | Source |
|---|---|---|---|---|
| A3BM0178 | Rodopi | Variety | Bulgarian | IPGR genebank |
| BGR3051 | Local population | Bulgarian | IPGR genebank | |
| BGR3052 | Local population | Bulgarian | IPGR genebank | |
| BGR 6207 | Local population | Bulgarian | IPGR genebank | |
| BGR13526 | Krasnodarskaia | Variety | Russian | IPGR genebank |
| C3000001 | CPI 10385 | Breeding line | Bulgarian | USDA, USA genebank |
| C3000002 | SH B7-3-3-1 | Breeding line | Bulgarian | USDA, USA genebank |
| C3000003 | Borina | Variety | Bulgarian | USDA, USA genebank |
| C3000006 | B92-198 | Breeding line | Bulgarian | USDA, USA genebank |
| C3000007 | B92-200 | Breeding line | Bulgarian | USDA, USA genebank |
| C3E0118 | Local population | Bulgarian | Ustrem village, Topolovgrad | |
| B9E0168 | Local population | Bulgarian | IPK Gatersleben genebank |
| 7 Feb–7 March | 8 March–5 April | 6 April– 4 May | 5 May– 2 June | 3 June– 1 July | 2–31 July | |
|---|---|---|---|---|---|---|
| Year 2022 | ||||||
| Minimum temp. (°C) | −0.73 | 3.98 | 5.39 | 10.14 | 15.51 | 15.53 |
| Maximum temp. (°C) | 9.10 | 11.36 | 18.27 | 25.47 | 27.79 | 31.32 |
| Mean temp. (°C) | 4.95 | 6.71 | 14.01 | 19.53 | 23.00 | 25.65 |
| Rainfall (mm) | 62.80 | 9.50 | 53.10 | 69.80 | 116.50 | 116.50 |
| Year 2023 | ||||||
| Minimum temp. (°C) | −0.51 | 2.69 | 6.70 | 10.44 | 14.42 | 18.09 |
| Maximum temp. (°C) | 11.19 | 14.51 | 16.57 | 20.26 | 26.77 | 32.81 |
| Mean temp. (°C) | 6.33 | 9.55 | 12.54 | 16.33 | 22.07 | 27.20 |
| Rainfall (mm) | 17.00 | 44.00 | 31.20 | 78.80 | 80.20 | 62.50 |
| Trait | Abbreviation | Classes |
|---|---|---|
| Vegetative phase | ||
| Serration of lower leaves | SLL | 1: serrate; 2: dentate; 3: entire |
| Stem anthocyanins | SA | 1: absent; 2: present |
| Plant Branching | PB | 1: strong basal; 2: only in the upper part |
| Presence of tendrils | PT | 1: absent; 2: present |
| Presence of leaves | PL | 1: absent; 2: present |
| Leaves color | LC | 1: yellow green; 2: green; 3: light green |
| Leaves shape | LSh | 1: narrow elliptic; 2: elliptic; 3: broad elliptic; 4: narrow ovate; 5: ovate; 6: broad ovate |
| Type of leaves | TL | 1: even pinnate; 2: odd pinnate; 3: acacia-like leaflets |
| Growth habit | GH | 1: bushy; 2: tall |
| Reproductive phase | ||
| Flowering duration | FD | Record at the beginning to end of flowering (days) |
| Flower: color of standard and wings | FCS/ FCW | 1: white; 2: creamy; 3: greenish; 4: pink; 5: reddish; 6: carmine; 7: light violet; 8: violet; 9: dark violet; 10: brown |
| Number of flowers in one inflorescence | NFI | 1: one; 2: one or two; 3: two; 4: two or three; 5: three or more |
| Flower size | FS | 1: small; 2: medium; 3: large |
| Pod traits | ||
| Pod shattering | PShat | 1: non-shattering; 2: shattering |
| Pod shape | PSh | 1: straight; 2: slightly curved; 3: markedly curved |
| Pod length and width | PL, PW | The observations on well-developed green pods; the width is assessed from suture to suture on unopened pods. |
| Seed traits | ||
| Ground color of testa | GCT | 1: yellow; 2: green; 3: yellow and green; 4: yellow to greenish gray; 5: green-grey to brown-grey; 6: reddish brown-grey; 7: dark brown, brownish-yellow; 8: carmine |
| Pattern of the seeds | PS | 0: absent; 1: present |
| Color of pattern of testa | CPT | 0: absent; 1: violet; 2: brown; 3: violet and brown |
| Seeds shape | SSh | 1: round, globular; 2: roundish-angular; 3: angular to compressed; 4: angular; 5: wrinkled; 6: granular surface |
| Grain yield | ||
| Height to the first pod | HFP | Record at the height of first pod/ten plants per accession |
| Number of pods/plant | NPP | Record number of harvested pods/ten plants per accession |
| Number of grains/pod | NGP | Record number of seeds per pod/ten plants per accession |
| Number of grains/plant | NGPl | Record number of grains per plant/ten plants per accession |
| Weight of grains/plant | WGPl | Record weight of grains per plant//ten plants per accession |
| 100-seed weight (g) | SW | Record the mass of 100 randomly taken seeds per accession |
| Harvest index (%) | HI | grain yield/total biomass × 100 |
| Trait | Abbreviation | Unit s |
|---|---|---|
| Economic traits—Yield structure | ||
| Plant height | PlH | cm |
| Height to the first pod | HFP | cm |
| Number of branches | NB | |
| Number of pods per plant | NPPl | |
| Number of grains per pod | NGP | |
| Number of grains per plant | NGPl | |
| Weight of grains per plants | WGPl | g |
| Length of pod | PL | mm |
| Width of pod | PW | mm |
| 100-seed weight | SW | g |
| Harvest index, | HI | % |
| Biological traits—phenology | ||
| Germination–50% flowering | G-Fl | days |
| Duration of flowering | DFl | days |
| Growth cycle | GC | days |
| Biochemical traits—seed components | ||
| Soluble protein | Pr | mg·g−1 FW |
| Soluble sugars | SS | mg·g−1 FW |
| Starch | St | mg·g−1 FW |
| Phenols | Phe | mg·g−1 FW |
| Free amino acids | AAs | mg·g−1 FW |
| Antitrypsin activity | AT | U·mg−1 FW |
| Accession | Anthocyanins | Growth Habit | Plant Branching | Leaf Color | Leaf Shape | Type of Leaf |
|---|---|---|---|---|---|---|
| A3BM0178 | no | bushy | strongly from the base | green | narrow elliptic | even pinnate |
| BGR3051 | yes | bushy | strongly from the base | green | narrow elliptic | odd pinnate |
| BGR3052 | yes | bushy | strongly from the base | green | narrow elliptic | even pinnate |
| BGR6207 | yes | bushy | strongly from the base | green | narrow elliptic | odd pinnate |
| BGR13526 | no | bushy | strongly from the base | green | narrow elliptic | even pinnate |
| C3000001 | no | tall | only in the upper part | green | elliptic | even pinnate |
| C3000002 | no | bushy | strongly from the base | green | elliptic | even pinnate |
| C3000003 | no | bushy | strongly from the base | yellow green | elliptic | even pinnate |
| C3000006 | no | bushy | strongly from the base | yellow green | elliptic | odd pinnate |
| C3000007 | no | bushy | strongly from the base | green | broad elliptic | odd pinnate |
| C3E0118 | no | bushy | strongly from the base | yellow green | elliptic | even pinnate |
| B9E0168 | no | bushy | strongly from the base | light green | narrow elliptic | even pinnate |
| Accession | Flower Size | Flowers in Inflorescence | Flowers: Color of Standard | Flowers: Color of Wings | Grain Shape | Ground Color of Testa | Testa Pattern |
|---|---|---|---|---|---|---|---|
| A3BM0178 | medium | 1–2 | creamy | creamy | roundish-angular | reddish brown | no |
| BGR3051 | medium | 2–3 | pink | pink | roundish-angular | reddish gray | no |
| BGR3052 | medium | 2–3 | creamy brown streaks | creamy | roundish-angular | reddish gray | no |
| BGR6207 | medium | 1–2 | creamy brown streaks | creamy | roundish-angular | reddish brown | no |
| BGR13526 | small | 1–2 | pink | pink | round-globular | reddish brown | no |
| C3000001 | medium | 1–2 | creamy brown streaks | creamy | round-globular | reddish brown | dots |
| C3000002 | medium | 2–3 | light violet | creamy | round-globular | reddish gray | no |
| C3000003 | medium | 2–3 | light violet | creamy | round-globular | reddish green brown | no |
| C3000006 | medium | 1–2 | creamy brown streaks | creamy | round-globular | brownish gray | dots |
| C3000007 | large | 3 or more | light violet | creamy | round-globular | reddish gray | no |
| C3E0118 | medium | 2–3 | creamy brown streaks | creamy | round-globular | reddish gray | no |
| B9E0168 | medium | 3 or more | creamy | creamy | roundish-angular | reddish brown | no |
| Accession | Plant Height, cm | Height to First pod, cm | Number of Branches | Number of Pods/Plant | Number of Grains/Pod | Number of Grains/Plant | Weight of Grains/Plant | Size of Pod | 100-Seed Weight, g | Harvest Index, % | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Length, mm | Width, mm | |||||||||||||||||||||
| M | DM | M | DM | M | DM | M | DM | M | DM | M | DM | M | DM | M | DM | M | DM | M | DM | M | DM | |
| A3BM0178 | 41.3 | 19.8 | 1.0 | 73.2 | 2.8 | 137.2 | 4.0 | 14.7 | 4.3 | 3.3 | 37.2 | |||||||||||
| BGR3051 | 50.7 | 9.4*** | 30.7 | 10.8*** | 1.2 | 0.2 | 32.8 | −40.3*** | 2.1 | −0.7*** | 55.0 | −82.2*** | 1.1 | −2.9*** | 12.5 | −2.3*** | 3.9 | −0.4*** | 2.2 | −1.1*** | 11.3 | −26.0*** |
| BGR3052 | 50.0 | 8.7*** | 28.1 | 8.3*** | 3.6 | 2.6*** | 25.5 | −47.7*** | 2.8 | 0.0 | 49.6 | −87.6*** | 1.2 | −2.9*** | 14.2 | −0.5 | 3.9 | −0.4*** | 2.6 | −0.8*** | 19.8 | −17.5*** |
| BGR 6207 | 57.0 | 15.8*** | 24.0 | 4.2** | 2.0 | 1.0** | 105 | 31.3*** | 3.3 | 0.5*** | 226 | 89.2*** | 9.0 | 5.0*** | 18.8 | 4.1*** | 4.4 | 0.1 | 4.0 | 0.7*** | 41.1 | 3.9* |
| BGR13526 | 46.8 | 5.6** | 24.6 | 4.8** | 3.4 | 2.4*** | 41.4 | −31.8*** | 2.8 | 0.0 | 63.3 | −73.9** | 2.3 | −1.7 | 17.6 | 2.8*** | 4.5 | 0.2 | 3.8 | 0.5*** | 24.1 | −13.1*** |
| B9E0168 | 64.2 | 22.9*** | 30.8 | 11.0*** | 1.7 | 0.7* | 97.5 | 24.3* | 2.9 | 0.1 | 194 | 57.0** | 6.2 | 2.1* | 16.3 | 1.5** | 4.2 | −0.1 | 4.1 | 0.8*** | 26.4 | −10.8*** |
| C3000002 | 43.3 | 2.1 | 17.7 | −2.2 | 1.3 | 0.3 | 65.3 | −7.8 | 3.3 | 0.5*** | 180 | 42.8 | 6.2 | 2.2* | 18.3 | 3.6*** | 4.5 | 0.2 | 3.9 | 0.6*** | 42.0 | 4.8* |
| C3000003 | 45.7 | 4.4* | 20.0 | 0.2 | 4.0 | 3.0*** | 159 | 86.2*** | 4.0 | 1.2*** | 367 | 230.2*** | 13 | 9.4*** | 22.4 | 7.6*** | 5.0 | 0.7*** | 4.1 | 0.8*** | 31.8 | −5.4** |
| C3000001 | 24.7 | −16.6*** | 7.7 | −12.2*** | 1.3 | 0.3 | 50.3 | −22.8 | 3.2 | 0.4*** | 131 | −6.2 | 6.7 | 2.7** | 20.0 | 5.3*** | 4.5 | 0.2 | 4.4 | 1.0*** | 57.2 | 20.0*** |
| C3000007 | 47.3 | 6.1** | 27.3 | 7.5*** | 1.0 | 0.0 | 39.0 | −34.2*** | 2.2 | −0.6*** | 76.3 | −60.8** | 2.2 | −1.8 | 13.1 | −1.6*** | 3.8 | −0.5*** | 3.1 | −0.2 | 20.8 | −16.4*** |
| C3000006 | 28.3 | −12.9*** | 10.3 | −9.5*** | 0.7 | −0.3 | 30.0 | −43.2*** | 3.3 | 0.5*** | 83.7 | −53.5** | 4.3 | 0.3 | 18.5 | 3.7*** | 4.6 | 0.3** | 4.6 | 1.2*** | 41.6 | 4.4** |
| C3E0118 | 48.3 | 7.0** | 21.7 | 1.9 | 1.3 | 0.3 | 99.7 | 26.5* | 3.3 | 0.5*** | 169 | 31.5 | 6.9 | 2.9*** | 16.2 | 1.5** | 4.1 | −0.2 | 3.5 | 0.2 | 38.1 | 0.9 |
| average | 45.6 | 21.9 | 1.9 | 68.2 | 3.0 | 144.4 | 5.3 | 16.9 | 4.3 | 3.6 | 32.6 | |||||||||||
| min | 24.7 | 7.7 | 0.7 | 25.5 | 2.1 | 49.6 | 1.1 | 12.5 | 3.8 | 2.2 | 11.3 | |||||||||||
| max | 64.2 | 30.8 | 4.0 | 159.3 | 4.0 | 367.3 | 13.4 | 22.4 | 5.0 | 4.6 | 57.2 | |||||||||||
| R | 39.50 | 23.1 | 3.3 | 133.8 | 1.9 | 317.7 | 12.3 | 9.9 | 1.2 | 2.4 | 45.9 | |||||||||||
| CV, % | 23.8 | 33.7 | 60.5 | 59.2 | 17.3 | 63.6 | 67.7 | 17.5 | 8.2 | 20.0 | 39.0 | |||||||||||
| Accession | Germination to 50% Flowering, Days | Duration of Flowering, Days | Growth Cycle, Days | Soluble Protein, mg·g−1 FW | Sugars, mg·g−1 FW | Starch, mg·g−1 FW | Phenols, mg·g−1 FW | Amino Acids, mg·g−1 FW | TIA, Units mg−1 FW | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | MD | M | MD | M | MD | M | MD | M | MD | M | MD | M | MD | M | MD | M | MD | |
| A3BM0178 | 73 | 14 | 112 | 105.4 | 28.73 | 571.13 | 0.219 | 0.030 | 1.967 | |||||||||
| BGR3051 | 79 | 6 | 12 | −2 | 116 | 4 | 100.8 | 4.60 | 35.55 | −6.82** | 561.75 | 9.38 | 0.210 | −0.009 | 0.030 | 0.001 | 2.224 | −0.590*** |
| BGR3052 | 66 | −7 | 8 | −6 | 101 | −11 | 86.05 | 19.35*** | 34.6 | −5.87** | 525.63 | 45.5 | 0.256 | 0.037 | 0.060 | −0.022*** | 2.387 | −1.293*** |
| BGR 6207 | 80 | 7 | 11 | 3 | 108 | −4 | 92.2 | 13.20*** | 33.55 | −4.82 | 615.00 | −43.87 | 0.260 | 0.041 | 0.040 | −0.005 | 1.699 | −0.502*** |
| BGR13526 | 59 | −14 | 6 | −8 | 94 | −18 | 86.05 | 19.35*** | 49.68 | −20.95*** | 627.75 | −56.62 | 0.329 | 0.110*** | 0.040 | −0.011 | 1.22 | 0.502*** |
| B9E0168 | 83 | 10 | 11 | −3 | 162 | 3 | 105.8 | −0.40 | 35.43 | −6.70** | 565.50 | 5.63 | 0.206 | −0.014 | 0.050 | −0.001 | 2.178 | −0.053 |
| C3000002 | 54 | −29 | 8 | −6 | 84 | −28 | 95.9 | 9.50*** | 38.40 | −9.67*** | 506.73 | 64.4** | 0.197 | −0.022 | 0.060 | −0.027*** | 2.550 | −0.308*** |
| C3000003 | 67 | −6 | 13 | −1 | 83 | −29 | 88.1 | 17.30*** | 36.20 | −7.47*** | 591.77 | −20.64 | 0.145 | −0.074*** | 0.077 | −0.044*** | 2.370 | −0.133 |
| C3000001 | 41 | −32 | 10 | −4 | 83 | −29 | 92.7 | 12.70*** | 26.07 | 2.66 | 598.90 | −27.77 | 0.180 | −0.040 | 0.068 | −0.035*** | 2.390 | −0.153 |
| C3000007 | 67 | −6 | 8 | −6 | 83 | −29 | 85.7 | 19.70*** | 41.33 | −12.6*** | 502.63 | 68.5** | 0.142 | −0.078*** | 0.056 | −0.023*** | 2.140 | 0.100 |
| C3000006 | 41 | −32 | 12 | −2 | 83 | −29 | 93.85 | 11.55*** | 21.67 | 7.06** | 581.70 | −10.57 | 0.211 | −0.009 | 0.061 | −0.0280*** | 2.080 | 0.158** |
| C3E0118 | 162 | 121 | 14 | 0 | 210 | 98 | 89.2 | 16.20*** | 29.87 | −1.14 | 509.73 | 61.4** | 0.147 | −0.072*** | 0.072 | −0.039*** | 2.500 | −0.263*** |
| average | 64.5 | 10.3 | 97.5 | 93.48 | 34.26 | 563.19 | 0.209 | 0.054 | 2.142 | |||||||||
| min | 41 | 6 | 83 | 85.70 | 21.67 | 502.63 | 0.142 | 0.030 | 1.220 | |||||||||
| max | 83 | 14 | 162 | 105.8 | 49.68 | 627.75 | 0.329 | 0.077 | 2.550 | |||||||||
| R | 42 | 8 | 33 | 20.10 | 28.01 | 125.12 | 0.187 | 0.047 | 1.330 | |||||||||
| CV, % | 22.6 | 24.3 | 14.7 | 6.8 | 21.4 | 7.7 | 26.3 | 29.4 | 17.6 | |||||||||
| PlH | HFP | NB | NPPl | NGP | NGPl | WGPl | PL | PW | SW | HI | G-Fl | DFl | GC | Pr | SS | St | Phe | AAs | AT | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PlH | 1 | 0.898** | 0.286 | 0.337 | −0.222 | 0.195 | 0.017 | −0.333 | −0.372 | −0.371 | 0.602* | 0.466 | −0.042 | 0.319 | 0.152 | 0.509 | −0.132 | 0.189 | −0.328 | −0.163 |
| HFP | 1 | 0.271 | 0.015 | −0.554 | −0.136 | −0.332 | −0.627* | −0.613* | −0.682* | −0.869** | 0.370 | −0.166 | 0.237 | 0.134 | 0.600* | −0.233 | 0.214 | −0.444 | −0.181 | |
| NB | 1 | 0.321 | 0.338 | 0.312 | 0.256 | 0.342 | 0.316 | −0.055 | −0.298 | −0.077 | −0.301 | −0.171 | −0.368 | 0.496 | 0.312 | 0.360 | 0.214 | −0.206 | ||
| NPPl | 1 | 0.695* | 0.963** | 0.910** | 0.583* | 0.538 | 0.369 | 0.233 | 0.380 | 0.513 | 0.239 | −0.074 | −0.014 | 0.178 | −0.363 | 0.327 | 0.145 | |||
| NGP | 1 | 0.782** | 0.867** | 0.907** | 0.839** | 0.719** | 0.640* | 0.021 | 0.308 | 0.007 | −0.241 | −0.304 | 0.269 | −0.191 | 0.639* | 0.209 | ||||
| NGPl | 1 | 0.965** | 0.711** | 0.661* | 0.474 | 0.342 | 0.166 | 0.435 | 0.028 | −0.089 | −0.067 | 0.191 | −0.394 | 0.412 | 0.225 | |||||
| WGPl | 1 | 0.828** | 0.734** | 0.617* | 0.521 | 0.132 | 0.431 | 0.032 | −0.171 | −0.194 | 0.273 | −0.398 | 0.517 | 0.219 | ||||||
| PL | 1 | 0.933** | 0.828** | 0.664* | −0.256 | 0.096 | −0.266 | −0.181 | −0.163 | 0.514 | −0.080 | 0.544 | 0.009 | |||||||
| PW | 1 | 0.771** | 0.560 | −0.346 | 0.165 | −0.343 | −0.029 | −0.13 | 0.581* | 0.009 | 0.349 | −0.112 | ||||||||
| SW | 1 | 0.758** | −0.275 | 0.066 | −0.236 | 0.033 | −0.295 | 0.444 | −0.064 | 0.426 | −0.105 | |||||||||
| HI | 1 | 0.140 | 0.220 | −0.036 | −0.11 | −0.576 | 0.222 | −0.200 | 0.396 | 0.178 | ||||||||||
| G-Fl | 1 | 0.472 | 0.964** | −0.07 | −0.025 | −0.369 | −0.280 | 0.099 | 0.198 | |||||||||||
| DFl | 1 | 0.496 | 0.193 | −0.671* | 0.035 | −0.486 | 0.052 | 0.318 | ||||||||||||
| GC | 1 | 0.022 | −0.167 | −0.313 | −0.190 | 0.061 | 0.19 | |||||||||||||
| Pr | 1 | −0.088 | 0.081 | 0.070 | −0.397 | 0.046 | ||||||||||||||
| SS | 1 | 0.002 | 0.367 | −0.241 | −0.443 | |||||||||||||||
| St | 1 | 0.530 | −0.288 | −0.677* | ||||||||||||||||
| Phe | 1 | −0.584* | −0.756** | |||||||||||||||||
| AAs | 1 | 0.611* | ||||||||||||||||||
| AT | 1 |
| Component | Total | Initial Eigenvalues | |
|---|---|---|---|
| % of Variance | Cumulative % | ||
| PC1 | 7.259 | 36.294 | 36.294 |
| PC2 | 3.98 | 19.899 | 56.193 |
| PC3 | 3.375 | 16.875 | 73.068 |
| Traits/Components | PC1 | PC2 | PC3 |
|---|---|---|---|
| Plant height, cm | −0.398 | 0.300 | 0.773 |
| Height to the first pod, cm | −0.706 | 0.232 | 0.633 |
| Number of branches | 0.129 | −0.323 | 0.721 |
| Number of pods per plant | 0.662 | 0.372 | 0.609 |
| Number of grains per pod | 0.936 | 0.021 | 0.191 |
| Number of grains per plant | 0.774 | 0.258 | 0.500 |
| Weight of grains per plants, g | 0.888 | 0.197 | 0.364 |
| length of pod, mm | 0.933 | −0.284 | 0.158 |
| width of pod, mm | 0.867 | −0.370 | 0.152 |
| Weight of 100 grains, g | 0.812 | −0.290 | −0.107 |
| Harvest index, % | 0.765 | −0.065 | −0.435 |
| Germination−50% flowering, days | −0.103 | 0.794 | 0.316 |
| Duration of flowering, days | 0.374 | 0.642 | −0.068 |
| Growth cycle, days | −0.107 | 0.742 | 0.146 |
| Protein,mg·g−1 FW % | −0.056 | 0.233 | −0.124 |
| Soluble sugars, mg·g−1 FW | −0.419 | −0.328 | 0.641 |
| Starch, mg·g−1 FW | 0.334 | −0.643 | 0.25 |
| Phenols, mg·g−1 FW | −0.348 | −0.701 | 0.268 |
| Amino acids, mg·g−1 FW | 0.623 | 0.265 | −0.106 |
| Antitrypsin activity, U·mg−1 FW | 0.233 | 0.673 | −0.347 |
| Extraction Method: Principal Component Analysis. | |||
| a. 3 components extracted. | |||
| Direct and Indirect Effect | Total Indirect Effect | Total Phenotypic Correlation with WGPl | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Plant Height | Height to First Pod | Number of Main Branches | Number of Pods/Plant | Number of Grains per Pod | Number of Grains per Plant | 100-Seed Weight | Harvest Index | |||
| PlH | −0.324 | −0.291 | −0.093 | −0.109 | 0.072 | −0.063 | 0.120 | 0.195 | −0.169 | 0.017 |
| HFP | 0.384 | 0.428 | 0.116 | 0.006 | −0.237 | −0.058 | −0.292 | −0.372 | −0.452 | −0.332 |
| NB | −0.003 | −0.003 | −0.010 | −0.003 | −0.003 | −0.003 | 0.001 | 0.003 | −0.012 | 0.256 |
| NPPl | 0.036 | 0.002 | 0.034 | 0.106 | 0.073 | 0.102 | 0.039 | 0.025 | 0.310 | 0.910** |
| NGP | −0.038 | −0.095 | 0.058 | 0.119 | 0.172 | 0.134 | 0.123 | 0.110 | 0.412 | 0.867** |
| NGPl | 0.141 | −0.099 | 0.226 | 0.698 | 0.566 | 0.724 | 0.521 | 0.248 | 2.301 | 0.965** |
| SW | −0.039 | −0.071 | −0.006 | 0.039 | 0.075 | 0.050 | 0.105 | 0.079 | 0.127 | 0.617* |
| HI | −0.140 | −0.203 | −0.069 | 0.054 | 0.149 | 0.080 | 0.177 | 0.233 | 0.048 | 0.521 |
| Direct and Indirect Effect | Total Indirect Effect | Total Phenotypic Correlation with WGPl | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Germination- 50%fl. | Flower duration | Growth Cycle | Protein | Sugars | Starch | Phenols | Amino Acids | Anti- trypsins | |||
| G-Fl | 2.262 | 1.068 | 2.181 | −0.025 | −0.057 | −0.835 | −0.633 | 0.224 | 0.448 | 2.371 | 0.132 |
| DFl | 0.128 | 0.271 | 0.134 | 0.099 | −0.182 | 0.009 | −0.132 | 0.014 | 0.086 | 0.157 | 0.431 |
| GC | −2.074 | −1.067 | −2.152 | −0.123 | 0.359 | 0.673 | 0.409 | −0.131 | −0.409 | −2.362 | 0.032 |
| Pr | −0.002 | 0.082 | 0.013 | 0.224 | −0.064 | 0.012 | −0.010 | −0.089 | 0.036 | −0.022 | −0.005 |
| SS | 0.002 | 0.053 | 0.013 | 0.023 | −0.079 | 0.000 | −0.029 | 0.019 | 0.035 | 0.116 | −0.194 |
| St | −0.233 | 0.022 | −0.198 | 0.035 | 0.001 | 0.631 | 0.334 | −0.182 | −0.427 | −0.647 | 0.273 |
| Phe | −0.053 | −0.093 | −0.036 | −0.008 | 0.070 | 0.101 | 0.191 | −0.112 | −0.144 | −0.275 | −0.398 |
| AAs | 0.065 | 0.034 | 0.040 | −0.260 | −0.158 | −0.189 | −0.383 | 0.655 | 0.400 | −0.450 | 0.517 |
| AT | 0.038 | 0.061 | 0.037 | 0.031 | −0.086 | −0.131 | −0.146 | 0.118 | 0.193 | −0.077 | 0.219 |
| Accession | Germination % | RST-G % | Seedling VI | RST-VI % | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| H2Od | PEG | NaCl | PEG | NaCl | H2Od | PEG | NaCl | PEG | NaCl | |
| A3BM0178 | 83.17 | 86.91 | 55.84 | 1.045 | 0.671 | 6.82 | 6.16 | 0.95*** | 0.903 | 0.139 |
| BGR3051 | 42.24 | 29.60* | 30.47 | 0.701 | 0.721 | 2.88 | 1.66 | 1.20*** | 0.576 | 0.417 |
| BGR3052 | 93.94 | 91.16 | 82.58 | 0.970 | 0.879 | 6.85 | 4.53** | 1.83*** | 0.661 | 0.267 |
| BGR 6207 | 71.86 | 67.59 | 44.38 | 0.941 | 0.617 | 5.28 | 3.64* | 1.36*** | 0.689 | 0.258 |
| BGR13526 | 70.91 | 72.40 | 32.07** | 1.021 | 0.452 | 5.22 | 6.18 | 2.28*** | 1.184 | 0.437 |
| C3000001 | 79.92 | 61.39 | 15.00** | 0.768 | 0.188 | 5.44 | 4.01 | 0.11*** | 0.737 | 0.020 |
| C3000002 | 95.00 | 98.04 | 63.49* | 1.032 | 0.668 | 8.26 | 5.47*** | 1.54*** | 0.662 | 0.186 |
| C3000003 | 95.96 | 73.44 | 69.34* | 0.765 | 0.723 | 7.52 | 4.30* | 2.23*** | 0.572 | 0.297 |
| C3000006 | 56.15 | 66.57 | 5.83** | 1.186 | 0.104 | 3.13 | 3.54 | nd | 1.131 | nd |
| C3000007 | 80.58 | 53.70* | 0.00*** | 0.666 | 0.000 | 7.51 | 3.09** | nd | 0.411 | nd |
| C3E0118 | 93.65 | 80.32 | 52.38* | 0.857 | 0.559 | 7.29 | 4.77* | 1.62*** | 0.743 | 0.222 |
| B9E0168 | 88.47 | 79.33 | 50.84* | 0.897 | 0.575 | 6.03 | 4.48*** | 1.17*** | 0.744 | 0.194 |
| average | 79.32 | 71.70 | 41.85 | 0.904 | 0.513 | 6.02 | 4.32 | 1.19 | 0.751 | 0.203 |
| CV% | 15.98 | 18.51 | 43.68 | 14.22 | 42.75 | 22.84 | 21.88 | 62.06 | 22.04 | 55.66 |
| Accession | Root Length (mm) | Shoot Length (mm) | Root/Shoot Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|
| H2Od | PEG | NaCl | H2Od | PEG | NaCl | H2Od | PEG | NaCl | |
| A3BM0178 | 48 | 59*** | 15*** | 37 | 30 | 6*** | 1. 511 | 2.266*** | 2.778*** |
| BGR3051 | 43 | 72** | 17** | 50 | 54 | 21** | 0.903 | 1.401* | 0.905 |
| BGR3052 | 49 | 53 | 12*** | 49 | 46 | 10*** | 1.080 | 1.279 | 1.341 |
| BGR 6207 | 36 | 41 | 14*** | 35 | 44 | 14*** | 1.032 | 1.018 | 1.235 |
| BGR13526 | 25 | 66** | 24 | 27 | 46* | 36 | 1.104 | 1.552 | 0.703 |
| C3000001 | 35 | 55** | nd | 29 | 26 | nd | 1.256 | 2.410*** | nd |
| C3000002 | 49 | 43 | 9*** | 33 | 38 | 6*** | 1.649 | 1.232* | 1.303 |
| C3000003 | 45 | 80*** | 20*** | 40 | 41 | 12*** | 1.255 | 2.233*** | 2.207* |
| C3000006 | 40 | 53 | 22 | 24 | 25 | 15 | 1.791 | 2.267 | 1.470 |
| C3000007 | 53 | 53 | nd | 37 | 24* | nd | 1.485 | 2.091* | nd |
| C3E0118 | 28 | 40** | 10*** | 30 | 23 | 14** | 1.084 | 2.045** | 0.847 |
| B9E0168 | 43 | 53 | 10*** | 42 | 44 | 10*** | 1.103 | 1.248 | 1.200 |
| average | 41 | 56 | 15 | 36 | 37 | 14 | 1.246 | 1.753 | 1.399 |
| CV% | 17 | 16 | 28 | 18 | 25 | 40 | 17.57 | 26.52 | 32.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrova, S.; Stoilova, T.; Velinov, V.; Vaseva, I.I.; Simova-Stoilova, L. Phenotypic Diversity and Abiotic Stress Tolerance Among Vicia ervilia (L.) Willd. Accessions. Plants 2025, 14, 1008. https://doi.org/10.3390/plants14071008
Petrova S, Stoilova T, Velinov V, Vaseva II, Simova-Stoilova L. Phenotypic Diversity and Abiotic Stress Tolerance Among Vicia ervilia (L.) Willd. Accessions. Plants. 2025; 14(7):1008. https://doi.org/10.3390/plants14071008
Chicago/Turabian StylePetrova, Sofiya, Tsvetelina Stoilova, Valentin Velinov, Irina I. Vaseva, and Lyudmila Simova-Stoilova. 2025. "Phenotypic Diversity and Abiotic Stress Tolerance Among Vicia ervilia (L.) Willd. Accessions" Plants 14, no. 7: 1008. https://doi.org/10.3390/plants14071008
APA StylePetrova, S., Stoilova, T., Velinov, V., Vaseva, I. I., & Simova-Stoilova, L. (2025). Phenotypic Diversity and Abiotic Stress Tolerance Among Vicia ervilia (L.) Willd. Accessions. Plants, 14(7), 1008. https://doi.org/10.3390/plants14071008