
Nutraceutical Profile Characterization in Apricot (Prunus armeniaca L.) Fruits

 , , , ,
, , , ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Evaluation of Pomological Traits Through Phenotyping
2.2. Evaluation of Metabolites Using Proton Nuclear Magnetic Resonance (1H NMR)
2.2.1. Amino Acids
2.2.2. Carbohydrates
2.2.3. Flavonoids and Alkaloids
2.2.4. Organic Acids
2.2.5. Polyphenol Derivatives and Alcohols
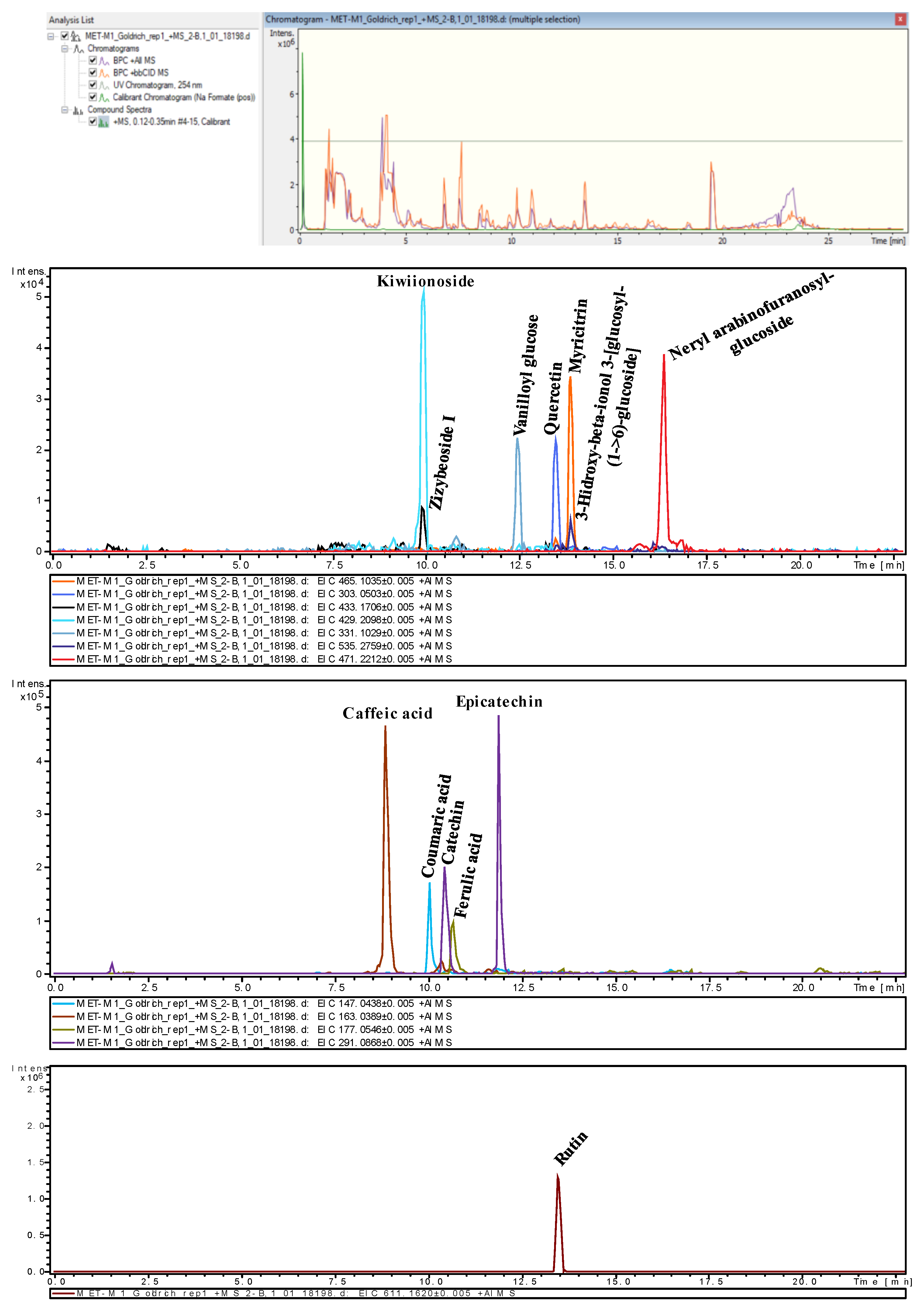
2.3. Evaluation of Metabolites Using Untargeted UPLC QToF MS/MS
2.3.1. Phenolic Acids
2.3.2. Flavonoids
2.3.3. Glycosides and Glucosylated Compounds
2.4. Non-Quantitative Area Values in Untargeted UPLC QToF MS/MS Analysis
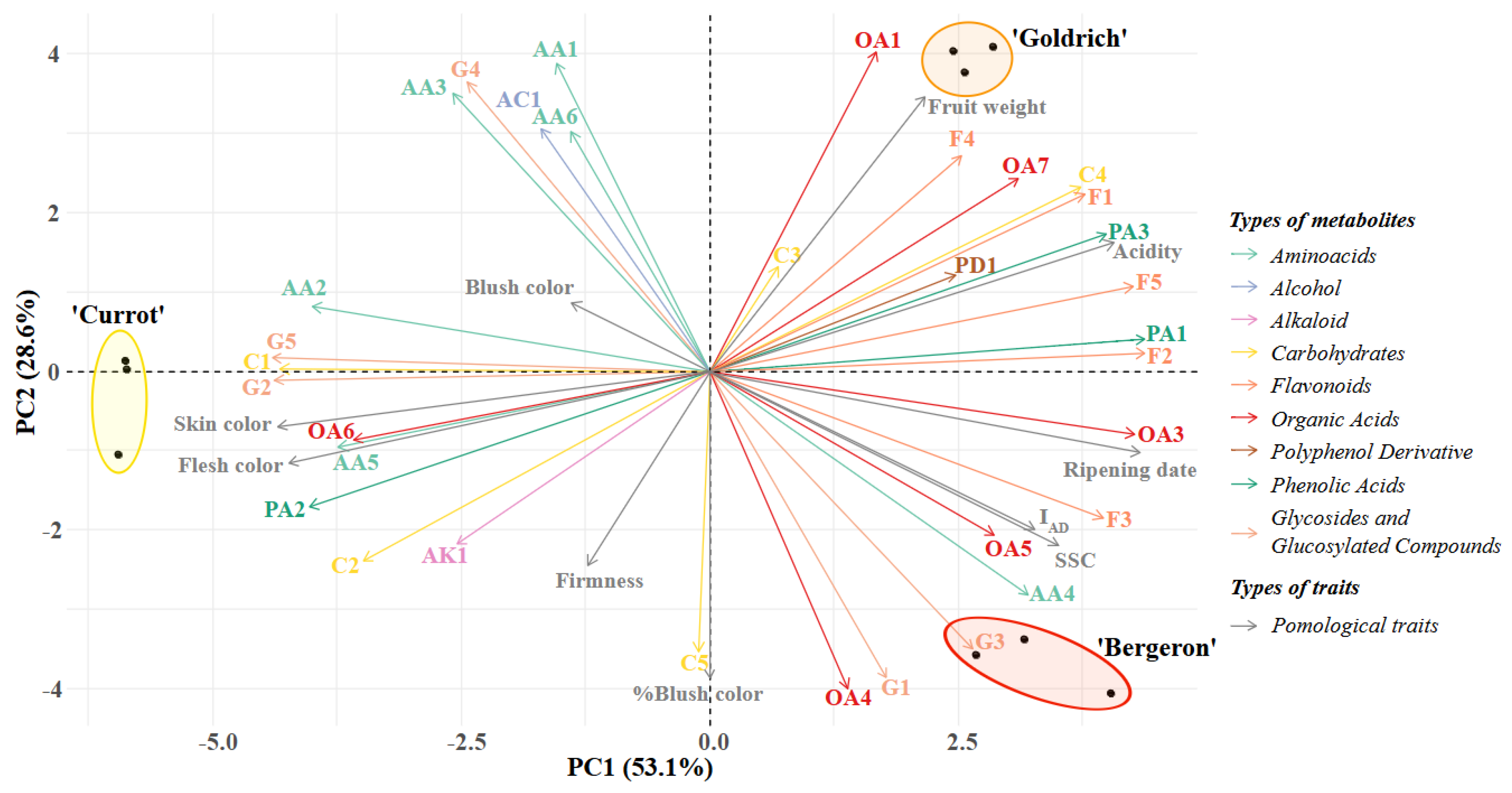
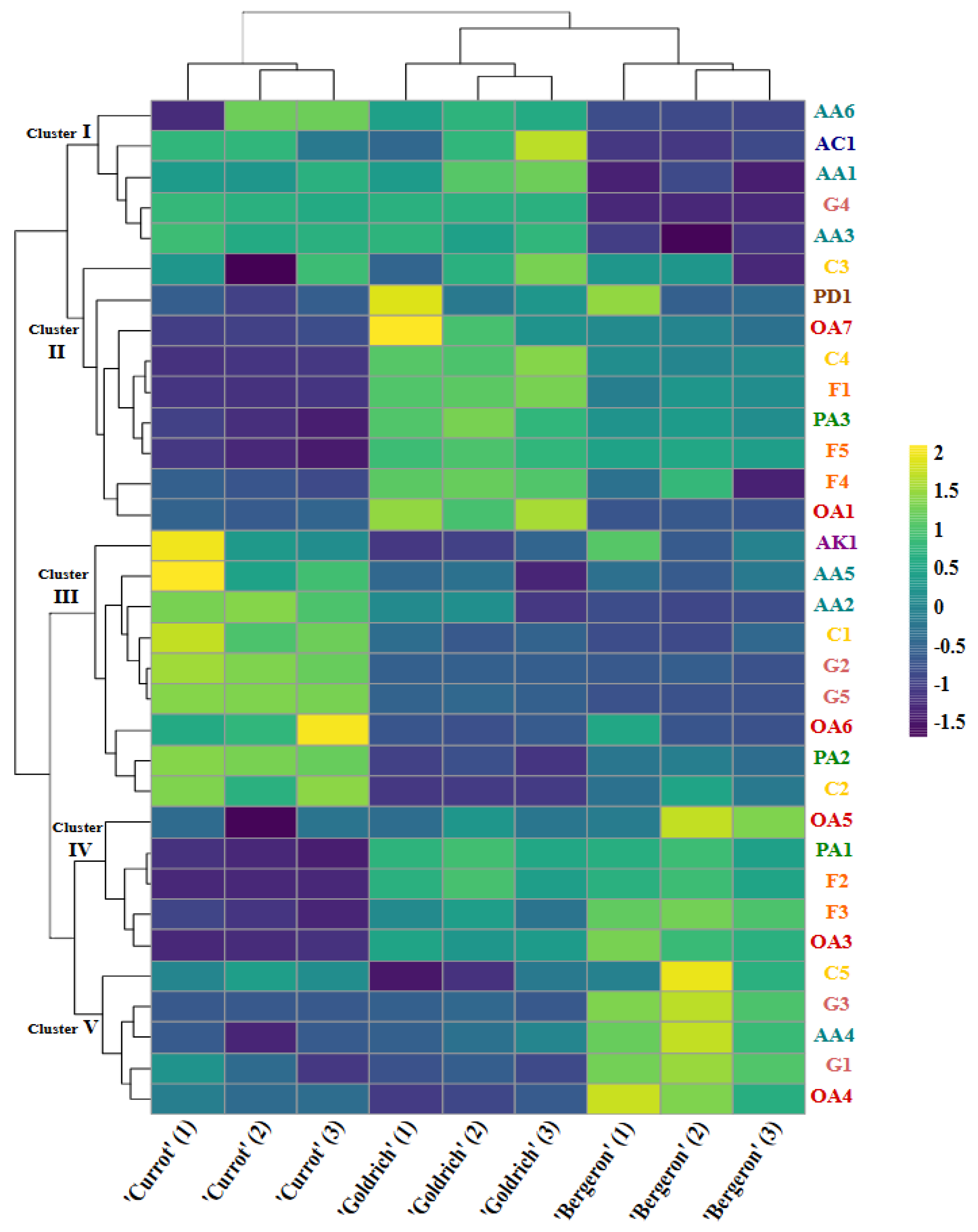
2.5. Comparative Metabolomic Profiling
2.5.1. First Cluster
2.5.2. Second Cluster
2.5.3. Third Cluster
2.5.4. Fourth Cluster
2.5.5. Fifth Cluster
3. Discussion
3.1. Primary Metabolites
3.1.1. Amino Acids
3.1.2. Carbohydrates
3.1.3. Organic Acids
3.2. Secondary Metabolites
3.2.1. Phenolic Acids
3.2.2. Flavonoids
3.2.3. Glycosides and Glucosylated Compounds
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design and Testing
4.3. Pomological Traits Analysis
4.4. Proton Nuclear Magnetic Resonance (1H NMR) Analysis
4.5. Untargeted UPLC QToF MS/MS Analysis
4.6. Metabolite Extraction Protocol from Untargeted UPLC QToF MS/MS
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Gómez, B.E.; Ruiz, D.; Salazar, J.A.; Rubio, M.; Martínez-García, P.J.; Martínez-Gómez, P. Analysis of Metabolites and Gene Expression Changes Relative to Apricot (Prunus Armeniaca L.) Fruit Quality During Development and Ripening. Front. Plant Sci. 2020, 11, 1269. [Google Scholar]
- Rampáčková, E.; Mrázová, M.; Čížková, J.; Nečas, T. Pomological Traits and Genome Size of Prunus Armeniaca L. Considering to Geographical Origin. Horticulturae 2022, 8, 199. [Google Scholar] [CrossRef]
- Ali, S.; Masud, T.; Abbasi, K.S.; Mahmood, T.; Hussain, A. Apricot: Nutritional potentials and health benefits—A review. Ann. Food Sci. Technol. 2015, 16, 175–189. [Google Scholar]
- Mesarović, J.; Trifković, J.; Tosti, T.; Fotirić Akšić, M.; Milatović, D.; Ličina, V.; Milojković-Opsenica, D. Relationship between Ripening Time and Sugar Content of Apricot (Prunus Armeniaca L.) Kernels. Acta Physiol. Plant 2018, 40, 157. [Google Scholar] [CrossRef]
- Gecer, M.K.; Kan, T.; Gundogdu, M.; Ercisli, S.; Ilhan, G.; Sagbas, H.I. Physicochemical Characteristics of Wild and Cultivated Apricots (Prunus Armeniaca L.) from Aras Valley in Turkey. Genet. Resour. Crop Evol. 2020, 67, 935–945. [Google Scholar] [CrossRef]
- Saeed, I.; Guo, X.; Azeem, M.; Elshikh, M.S.; Zainab, B.; Ayaz, Z.; You, L.; Alwahibi, M.S.; Mehmood Abbasi, A. Comparative Assessment of Polyphenolics’ Content, Free Radicals’ Scavenging and Cellular Antioxidant Potential in Apricot Fruit. J. King Saud. Univ. Sci. 2021, 33, 101459. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant Activity of Polyphenols, Flavonoids, Anthocyanins and Carotenoids: Updated Review of Mechanisms and Catalyzing Metals. Phytotherapy Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Fatima, T.; Bashir, O.; Gani, G.; Bhat, T.; Jan, N. Nutritional and Health Benefits of Apricots. Int. J. Unani Integr. Med. 2018, 2, 5–9. [Google Scholar]
- Siddiqui, S.A.; Anwar, S.; Yunusa, B.M.; Nayik, G.A.; Mousavi Khaneghah, A. The Potential of Apricot Seed and Oil as Functional Food: Composition, Biological Properties, Health Benefits & Safety. Food Biosci. 2023, 51, 102336. [Google Scholar] [CrossRef]
- Saini, D.; Rawat, N.; Negi, T.; Barthwal, R.; Sharma, S.K. Utilization, valorization and functional properties of wild apricot kernels. J. Pharmacogn. Phytochem. 2021, 10, 119–126. [Google Scholar]
- FAOSTAT. Global Crops Production Quantity of Apricots. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 25 April 2024).
- Iezzoni, A.F.; McFerson, J.; Luby, J.; Gasic, K.; Whitaker, V.; Bassil, N.; Yue, C.; Gallardo, K.; McCracken, V.; Coe, M.; et al. RosBREED: Bridging the Chasm between Discovery and Application to Enable DNA-Informed Breeding in Rosaceous Crops. Hortic. Res. 2020, 7, 177. [Google Scholar] [CrossRef] [PubMed]
- Bassi, D.; Rizzo, M.; Foschi, S. Breeding Apricot in Northern Italy. Acta Hortic. 2010, 862, 151–158. [Google Scholar] [CrossRef]
- Bircan, M.; Pinar, H.; Yilmaz, C.; Yildiz, A.; Paydaş, S.; Kaşka, N. The Apricot Breeding Program among Some Turkish and Foreign Cultivars. Acta Hort. 2010, 862, 103–108. [Google Scholar] [CrossRef]
- Sun, H.Y.; Zhang, J.H.; Yang, L.; Jiang, F.C.; Wang, Y.Z. An Overview of Apricot Breeding in China. In Proceedings of the XVI International Symposium on Apricot Breeding and Culture 1214, Shenyang, China, 29 June–3 July 2015; pp. 211–216. [Google Scholar]
- Ortuño-Hernández, G.; Sánchez, M.; Ruiz, D.; Martínez-Gómez, P.; Salazar, J.A. Monitoring Fruit Growth and Development in Apricot (Prunus Armeniaca L.) through Gene Expression Analysis. Int. J. Mol. Sci. 2024, 25, 9081. [Google Scholar] [CrossRef]
- García-Gómez, B.E.; Salazar, J.A.; Dondini, L.; Martínez-Gómez, P.; Ruiz, D. Identification of QTLs Linked to Fruit Quality Traits in Apricot (Prunus Armeniaca L.) and Biological Validation through Gene Expression Analysis Using QPCR. Mol. Breed. 2019, 39, 28. [Google Scholar] [CrossRef]
- Ortuño-Hernández, G.; Fernández, M.; Martínez-Gómez, P.; Ruiz, D.; Salazar, J.A. Ripening-Related Gene Expression Analysis Revealed the Molecular Impact of 1-MCP Application on Apricot Fruit Softening, Color, Aroma, and Antioxidant Capacity. Postharvest Biol. Technol. 2024, 216, 113037. [Google Scholar] [CrossRef]
- Ortuño-Hernández, G.; Ruiz, D.; Martínez-Gómez, P.; Salazar, J.A. Differentially Methylated DNA Regions in Apricot (Prunus Armeniaca L.) and Japanese Plum (Prunus Salicina L.) during Fruit Ripening after Ethylene-Related Treatments. Sci. Hortic. 2024, 330, 113052. [Google Scholar] [CrossRef]
- Hong, Y.; Wang, Z.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. High-Throughput Screening and Characterization of Phenolic Compounds in Stone Fruits Waste by LC-ESI-QTOF-MS/MS and Their Potential Antioxidant Activities. Antioxidants 2021, 10, 234. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, J.; Pu, X.; Shi, X.; Cheng, W.; Wang, B. Volatile Compounds Analysis and Biomarkers Identification of Four Native Apricot (Prunus Armeniaca L.) Cultivars Grown in Xinjiang Region of China. Foods 2022, 11, 2297. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Sharif, Y.; Zafar, M.H.; Ali, H.; Khan, K.A. Role of Primary Metabolites in Plant Defense against Pathogens. Microb. Pathog. 2019, 137, 103728. [Google Scholar] [CrossRef]
- Salam, U.; Ullah, S.; Tang, Z.-H.; Elateeq, A.A.; Khan, Y.; Khan, J.; Khan, A.; Ali, S. Plant Metabolomics: An Overview of the Role of Primary and Secondary Metabolites against Different Environmental Stress Factors. Life 2023, 13, 706. [Google Scholar] [CrossRef] [PubMed]
- Keutgen, A.J.; Pawelzik, E. Contribution of Amino Acids to Strawberry Fruit Quality and Their Relevance as Stress Indicators under NaCl Salinity. Food Chem. 2008, 111, 642–647. [Google Scholar] [CrossRef]
- Beauvoit, B.; Belouah, I.; Bertin, N.; Cakpo, C.B.; Colombié, S.; Dai, Z.; Gautier, H.; Génard, M.; Moing, A.; Roch, L.; et al. Putting Primary Metabolism into Perspective to Obtain Better Fruits. Ann. Bot. 2018, 122, 1–21. [Google Scholar] [CrossRef]
- Famiani, F.; Bonghi, C.; Chen, Z.-H.; Drincovich, M.F.; Farinelli, D.; Lara, M.V.; Proietti, S.; Rosati, A.; Vizzotto, G.; Walker, R.P. Stone Fruits: Growth and Nitrogen and Organic Acid Metabolism in the Fruits and Seeds—A Review. Front. Plant Sci. 2020, 11, 1–16. [Google Scholar]
- Chong, H.Y.; Pan, S.Z.; Li, B.J.; Liu, G.H.; Wang, D.B. Content changes of main organic acids and amino acids in Jiuqian plum during its fruit developing stage. Hubei Agric. Sci. 2022, 61, 92–95. [Google Scholar] [CrossRef]
- Sochor, J.; Skutkova, H.; Babula, P.; Zitka, O.; Cernei, N.; Rop, O.; Krska, B.; Adam, V.; Provazník, I.; Kizek, R. Mathematical Evaluation of the Amino Acid and Polyphenol Content and Antioxidant Activities of Fruits from Different Apricot Cultivars. Molecules 2011, 16, 7428–7457. [Google Scholar] [CrossRef]
- Echeverría, G.; Cantín, C.M.; Ortiz, A.; López, M.L.; Lara, I.; Graell, J. The Impact of Maturity, Storage Temperature and Storage Duration on Sensory Quality and Consumer Satisfaction of ‘Big Top®’ Nectarines. Sci. Hortic. 2015, 190, 179–186. [Google Scholar] [CrossRef]
- Ledbetter, C.; Peterson, S.; Jenner, J. Modification of Sugar Profiles in California Adapted Apricots (Prunus Armeniaca L.) through Breeding with Central Asian Germplasm. Euphytica 2006, 148, 251–259. [Google Scholar] [CrossRef]
- Fan, X.; Zhao, H.; Wang, X.; Cao, J.; Jiang, W. Sugar and Organic Acid Composition of Apricot and Their Contribution to Sensory Quality and Consumer Satisfaction. Sci. Hortic. 2017, 225, 553–560. [Google Scholar] [CrossRef]
- Koehler, P.E.; Kays, S.J. Sweet potato flavor: Quantitative and qualitative assessment of optimum sweetness. J. Food Qual. 1991, 14, 241–249. [Google Scholar] [CrossRef]
- Naryal, A.; Acharya, S.; Kumar Bhardwaj, A.; Kant, A.; Chaurasia, O.P.; Stobdan, T. Altitudinal Effect on Sugar Contents and Sugar Profiles in Dried Apricot (Prunus Armeniaca L.) Fruit. J. Food Compos. Anal. 2019, 76, 27–32. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Ivancic, A.; Schmitzer, V.; Veberic, R.; Stampar, F. Comparison of Major Taste Compounds and Antioxidative Properties of Fruits and Flowers of Different Sambucus Species and Interspecific Hybrids. Food Chem. 2016, 200, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Ayour, J.; Sagar, M.; Harrak, H.; Alahyane, A.; Alfeddy, M.N.; Taourirte, M.; Benichou, M. Evolution of Some Fruit Quality Criteria during Ripening of Twelve New Moroccan Apricot Clones (Prunus Armeniaca L.). Sci. Hortic. 2017, 215, 72–79. [Google Scholar] [CrossRef]
- Melgarejo, P.; Calín-Sánchez, Á.; Carbonell-Barrachina, Á.A.; Martínez-Nicolás, J.J.; Legua, P.; Martínez, R.; Hernández, F. Antioxidant Activity, Volatile Composition and Sensory Profile of Four New Very-Early Apricots (Prunus Armeniaca L.). J. Sci. Food Agric. 2014, 94, 85–94. [Google Scholar] [CrossRef]
- Bumrungpert, A.; Lilitchan, S.; Tuntipopipat, S.; Tirawanchai, N.; Komindr, S. Ferulic Acid Supplementation Improves Lipid Profiles, Oxidative Stress, and Inflammatory Status in Hyperlipidemic Subjects: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients 2018, 10, 713. [Google Scholar] [CrossRef]
- Raj, N.D.; Singh, D. A Critical Appraisal on Ferulic Acid: Biological Profile, Biopharmaceutical Challenges and Nano Formulations. Health Sci. Rev. 2022, 5, 100063. [Google Scholar] [CrossRef]
- Kumar, N.; Pruthi, V. Potential Applications of Ferulic Acid from Natural Sources. Biotechnol. Rep. 2014, 4, 86–93. [Google Scholar] [CrossRef]
- Silva, T.; Oliveira, C.; Borges, F. Caffeic Acid Derivatives, Analogs and Applications: A Patent Review (2009–2013). Expert. Opin. Ther. Pat. 2014, 24, 1257–1270. [Google Scholar] [CrossRef]
- da Silva, A.P.G.; Sganzerla, W.G.; John, O.D.; Marchiosi, R.A. Comprehensive Review of the Classification, Sources, Biosynthesis, and Biological Properties of Hydroxybenzoic and Hydroxycinnamic Acids. Phytochem. Rev. 2023, 1–30. [Google Scholar] [CrossRef]
- Aldaba-Muruato, L.R.; Ventura-Juárez, J.; Perez-Hernandez, A.M.; Hernández-Morales, A.; Muñoz-Ortega, M.H.; Martínez-Hernández, S.L.; Alvarado-Sánchez, B.; Macías-Pérez, J.R. Therapeutic Perspectives of P-Coumaric Acid: Anti-Necrotic, Anti-Cholestatic and Anti-Amoebic Activities. World Acad. Sci. J. 2021, 3, 1–8. [Google Scholar] [CrossRef]
- Hewawansa, U.H.A.J.; Houghton, M.J.; Barber, E.; Costa, R.J.S.; Kitchen, B.; Williamson, G. Flavonoids and Phenolic Acids from Sugarcane: Distribution in the Plant, Changes during Processing, and Potential Benefits to Industry and Health. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13307. [Google Scholar] [CrossRef] [PubMed]
- Espíndola, K.M.M.; Ferreira, R.G.; Narvaez, L.E.M.; Silva Rosario, A.C.R.; da Silva, A.H.M.; Silva, A.G.B.; Vieira, A.P.O.; Monteiro, M.C. Chemical and Pharmacological Aspects of Caffeic Acid and Its Activity in Hepatocarcinoma. Front. Oncol. 2019, 9, 541. [Google Scholar] [PubMed]
- Ruiz, D.; Egea, J.; Gil, M.I.; Tomás-Barberán, F.A. Characterization and Quantitation of Phenolic Compounds in New Apricot (Prunus Armeniaca L.) Varieties. J. Agric. Food Chem. 2005, 53, 9544–9552. [Google Scholar] [CrossRef]
- Sochor, J.; Zitka, O.; Skutkova, H.; Pavlik, D.; Babula, P.; Krska, B.; Horna, A.; Adam, V.; Provaznik, I.; Kizek, R. Content of Phenolic Compounds and Antioxidant Capacity in Fruits of Apricot Genotypes. Molecules 2010, 15, 6285–6305. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, H.J. Effects of Apricot and Apricot Kernels on Human Health and Nutrition: A Review of Recent Human Research. Technium BioChemMed 2021, 2, 139–162. [Google Scholar]
- Battistoni, B.; Ibáñez, J.; Ballesta, P.; Fiol, A.; Ahumada, S.; Meisel, L.A.; Morales, H.; Peña-Neira, A.; Lin-Wang, K.; Allan, A.C.; et al. The R2R3 MYB700 activates the expression of biosynthetic genes involved in the accumulation of (+)-catechin but not (−)-epicatechin in Chinese plum fruits (Prunus salicina Lindl.). Sci. Hortic. 2024, 338, 113798. [Google Scholar] [CrossRef]
- Jang, G.H.; Kim, H.W.; Lee, M.K.; Jeong, S.Y.; Bak, A.R.; Lee, D.J.; Kim, J.B. Characterization and Quantification of Flavonoid Glycosides in the Prunus Genus by UPLC-DAD-QTOF/MS. Saudi J. Biol. Sci. 2018, 25, 1622–1631. [Google Scholar] [CrossRef]
- Liang, Z.; Fang, Z.; Pai, A.; Luo, J.; Gan, R.; Gao, Y.; Lu, J.; Zhang, P. Glycosidically Bound Aroma Precursors in Fruits: A Comprehensive Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 215–243. [Google Scholar] [CrossRef]
- Rashid, F.; Ahmed, R.; Mahmood, A.; Ahmad, Z.; Bibi, N.; Kazmi, S.U. Flavonoid Glycosides FromPrunus Armeniaca and the Antibacterial Activity of a Crude Extract. Arch. Pharm. Res. 2007, 30, 932–937. [Google Scholar] [CrossRef]
- Johnson, J.B.; Mani, J.S.; Broszczak, D.; Prasad, S.S.; Ekanayake, C.P.; Strappe, P.; Valeris, P.; Naiker, M. Hitting the Sweet Spot: A Systematic Review of the Bioactivity and Health Benefits of Phenolic Glycosides from Medicinally Used Plants. Phytotherapy Res. 2021, 35, 3484–3508. [Google Scholar] [CrossRef]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Kathuria, D.; Barthwal, R.; Joshi, R. Metabolomics of Chemical Constituents as a Tool for Understanding the Quality of Fruits during Development and Processing Operations. Int. J. Food Sci. Technol. 2024, 59, 4169–4184. [Google Scholar] [CrossRef]
- Brown, G.S.; Walker, T.D. Indicators of Maturity in Apricots Using Biplot Multivariate Analysis. J. Sci. Food Agric. 1990, 53, 321–331. [Google Scholar] [CrossRef]
- van der Sar, S.; Kim, H.K.; Meissner, A.; Verpoorte, R.; Choi, Y.H. Nuclear Magnetic Resonance Spectroscopy for Plant Metabolite Profiling. In The Handbook of Plant Metabolomics; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 57–76. ISBN 9783527669882. [Google Scholar]
- Gil Solsona, R.; Boix, C.; Ibáñez, M.; Sancho, J. V The Classification of Almonds (Prunus Dulcis) by Country and Variety Using UHPLC-HRMS-Based Untargeted Metabolomics. Food Addit. Contam. Part A 2018, 35, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Guillamón, J.G.; Prudencio, Á.S.; Yuste, J.E.; Dicenta, F.; Sánchez-Pérez, R. Ascorbic Acid and Prunasin, Two Candidate Biomarkers for Endodormancy Release in Almond Flower Buds Identified by a Nontargeted Metabolomic Study. Hortic. Res. 2020, 7, 203. [Google Scholar] [CrossRef]
- KEGG: Kyoto Encyclopedia of Genes and Genomes. Available online: https://www.genome.jp/kegg/ (accessed on 18 June 2024).
- HMDB. Human Metabolome Database. Available online: https://hmdb.ca/ (accessed on 18 June 2024).
- Matthiesen, R. Mass Spectrometry Data Analysis in Proteomics; Springer: Berlin/Heidelberg, Germany, 2007; Volume 367, ISBN 158829563X. [Google Scholar]
- Amelin, V.G.; Bol’shakov, D.S. Simultaneous Identification and Determination by LC-MS of Quaternary Ammonium Compounds with Other Active Ingredients in Drugs. Pharm. Chem. J. 2020, 54, 79–85. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Apricot Cultivar | Trait | Mean ± SD | Apricot Cultivar | Trait | Mean ± SD | ||
|---|---|---|---|---|---|---|---|
| ‘Bergeron’ | Ripening date | 167 | ‘Goldrich’ | Ripening date | 157 | ||
 | Fruit weight | 40.41 ± 4.45 |  | Fruit weight | 86.57 ± 14.39 | ||
| IAD | 0.34 ± 0.13 | IAD | 0.22 ± 0.08 | ||||
| Skin color | 77.82 ± 2.51 | Skin color | 72.61 ± 1.21 | ||||
| Blush color | 48.11 ± 8.11 | Blush color | 56.09 ± 6.23 | ||||
| % Blush color | 26.25 ± 6.78 | % Blush color | 9.28 ± 5.35 | ||||
| Flesh color | 73.83 ± 1.84 | Flesh color | 70.70 ± 0.71 | ||||
| Firmness | 47.22 ± 12.74 | Firmness | 34.44 ± 15.63 | ||||
| SSC | 14.30 ± 0.53 | SSC | 13.13 ± 0.47 | ||||
| Acidity | 2.19 ± 0.11 | Acidity | 2.64 ± 0.09 | ||||
| Apricot cultivar | Trait | Mean ± SD | |||||
| ‘Currot’ | Ripening date | 130 | |||||
 | Fruit weight | 38.84 ± 3.32 | |||||
| IAD | 0.10 ± 0.05 | ||||||
| Skin color | 98.34 ± 2.71 | ||||||
| Blush color | 58.39 ± 12.92 | ||||||
| % Blush color | 18.75 ± 7.42 | ||||||
| Flesh color | 98.14 ± 2.18 | ||||||
| Firmness | 49.63 ± 10.11 | ||||||
| SSC | 12.03 ± 0.75 | ||||||
| Acidity | 1.44 ± 0.05 | ||||||
| Class | ID | Compound | Formula | 1H (ppm) a | Multiplicity b | ‘Bergeron’ (mg/g) cd | ‘Currot’ (mg/g) cd | ‘Goldrich’ (mg/g) cd |
|---|---|---|---|---|---|---|---|---|
| Amino acids | AA1 | Alanine | C3H7NO2 | 1.49 | d | 0.47 ± 0.06 b | 0.80 ± 0.04 a | 0.88 ± 0.09 a |
| AA2 | Asparagine | C4H8N2O3 | 2.95 | dd | 8.93 ± 0.32 b | 24.44 ± 1.43 a | 13.27 ± 5.25 b | |
| AA3 | Isoleucine | C6H13NO2 | 0.98 | t | 0.11 ± 0.01 b | 0.18 ± 0.01 a | 0.17 ± 0.01 a | |
| AA4 | Phenylalanine | C6H11NO2 | 7.40 | m | 0.21 ± 0.02 a | 0.12 ± 0.02 b | 0.15 ± 0.01 b | |
| AA5 | Threonine | C4H9NO3 | 1.32 | d | 0.22 ± 0.01 b | 0.30 ± 0.05 a | 0.20 ± 0.03 b | |
| AA6 | Valine | C5H11NO2 | 1.00 | d | 0.163 ± 0.003 a | 0.23 ± 0.07 a | 0.24 ± 0.01 a | |
| Alcohol | AC1 | Choline | (C5H14NO)+ | 3.20 | s | 0.032 ± 0.002 a | 0.06 ± 0.01 a | 0.06 ± 0.02 a |
| Alkaloid | AK1 | Trigonelline | C7H7NO2 | 9.10 | s | 0.04 ± 0.01 a | 0.04 ± 0.01 a | 0.029 ± 0.003 a |
| Carbohydrates | C1 | Fructose | C6H12O6 | 4.11 | m | 54.26 ± 3.08 b | 79.93 ± 4.48 a | 56.72 ± 1.86 b |
| C2 | Glucose | C6H12O6 | 5.22 | d | 139.68 ± 15.47 b | 180.83 ± 14.24 a | 100.75 ± 1.00 c | |
| C3 | Myo-Inositol | C6H12O6 | 3.30 | t | 2.57 ± 0.20 a | 2.58 ± 0.30 a | 2.74 ± 0.20 a | |
| C4 | Sucrose | C12H22O11 | 5.40 | d | 464.65 ± 4.89 b | 370.64 ± 3.66 c | 546.98 ± 14.85 a | |
| C5 | Xylose | C5H10O5 | 5.10 | d | 2.46 ± 0.34 a | 2.22 ± 0.07 ab | 1.85 ± 0.22 b | |
| Flavonoid | F1 | Epicatechin | C15H14O6 | 6.05 | d | 0.44 ± 0.06 b | 0.03 ± 0.01 c | 0.79 ± 0.05 a |
| Organic Acids | OA1 | Citrate | (C6H5O7)−3 | 2.74 | dd | 64.12 ± 1.86 b | 73.98 ± 4.79 b | 202.27 ± 22.37 a |
| OA2 | Formate | (CHO2)− | 8.45 | s | Not detected | Not detected | Not detected | |
| OA3 | Fumarate | (C4H2O4)−2 | 6.53 | s | 0.025 ± 0.002 a | 0.0126 ± 0.0003 c | 0.0218 ± 0.0007 b | |
| OA4 | Malate | (C4H4O5)−2 | 2.39 | dd | 109.16 ± 13.25 a | 71.34 ± 4.00 b | 57.56 ± 5.22 b | |
| OA5 | Quinate | (C7H11O6)− | 1.95 | dd | 49.33 ± 3.95 a | 42.32 ± 3.04 a | 45.03 ± 1.30 a | |
| OA6 | Succinate | (C4H4O4)−2 | 2.49 | s | 0.04 ± 0.03 ab | 0.09 ± 0.03 a | 0.025 ± 0.003 b | |
| OA7 | Tartrate | (C4H4O6)−2 | 7.10 | s | 0.91 ± 0.05 ab | 0.66 ± 0.03 b | 1.23 ± 0.26 a | |
| Polyphenol Derivative | PD1 | Chlorogenate | (C16H17O9)− | 7.63 | d | 1.64 ± 0.87 a | 0.95 ± 0.16 a | ± 0.84 a |
| Class | ID | Tentative Compound | Formula | Theoretical Mass (m/z) | Measured Mass (m/z) | RT (min) | Error | mSigma | Adduct | Database |
|---|---|---|---|---|---|---|---|---|---|---|
| Phenolic Acids | PA1 | Caffeic acid | C9H8O4 | 163.0390 | 163.0389 | 9.03 | 0.2 | 8.7 | [M−H2O+H]+ | KEGG |
| PA2 | Coumaric acid | C9H8O3 | 147.0441 | 147.0438 | 10.06 | 2.0 | 6.8 | [M−H2O+H]+ | KEGG | |
| PA3 | Ferulic acid | C10H10O4 | 177.0546 | 177.0546 | 10.76 | 0.0 | 2.9 | [M−H2O+H]+ | KEGG | |
| Flavonoids | F2 | Catechin | C15H14O6 | 291.0863 | 291.0868 | 10.43 | −1.8 | 6.2 | [M+H]+ | HMDB |
| F3 | Myricitrin | C21H20O12 | 465.1028 | 465.1035 | 13.92 | −1.6 | 1.2 | [M+H]+ | HMDB | |
| F4 | Quercetin | C15H10O7 | 303.0499 | 303.0503 | 13.50 | −1.1 | 13.9 | [M+H]+ | HMDB | |
| F5 | Rutin | C27H30O16 | 611.1607 | 611.1620 | 13.54 | −2.2 | 16.0 | [M+H]+ | HMDB | |
| Glycosides and Glucosylated Compounds | G1 | Kiwiionoside | C19H34O9 | 429.2095 | 429.2098 | 10.00 | −0.8 | 5.3 | [M+Na]+ | HMDB |
| G2 | Neryl arabinofuranosyl-glucoside | C21H36O10 | 471.2201 | 471.2212 | 16.37 | −2.4 | 6.4 | [M+Na]+ | HMDB | |
| G3 | Vanilloyl glucose | C14H18O9 | 331.1024 | 331.1029 | 12.54 | −1.5 | 3.0 | [M+H]+ | HMDB | |
| G4 | Zizybeoside I | C19H28O11 | 433.1704 | 433.1706 | 9.97 | −0.3 | 29.9 | [M+H]+ | HMDB | |
| G5 | 3-Hydroxy-beta-ionol 3-[glucosyl-(1->6)-glucoside] | C25H42O12 | 535.2749 | 535.2759 | 13.94 | −1.9 | 11.2 | [M+H]+ | HMDB |
| Class | ID | Tentative Compound | ‘Bergeron’ (ra) | ‘Currot’ (ra) | ‘Goldrich’ (ra) |
|---|---|---|---|---|---|
| Phenolic Acids | PA1 | Caffeic acid | 4,609,160 ± 463,508 a | 532,861 ± 219,791 b | 4,781,477 ± 368,002 a |
| PA2 | Coumaric acid | 1,040,945 ± 90,857 b | 2,140,665 ± 76,633 a | 508,567 ± 119,475 c | |
| PA3 | Ferulic acid | 917,099 ± 21,049 b | 640,206 ± 38,056 c | 1,067,681 ± 50,773 a | |
| Flavonoids | F2 | Catechin | 2,424,284 ± 249,228 a | 17,918 ± 2567 b | 2,400,544 ± 330,572 a |
| F3 | Myricitrin | 362,724 ± 12,724 a | 138,008 ± 19,084 c | 256,125 ± 32,223 b | |
| F4 | Quercetin | 94,999 ± 67,109 ab | 67,531 ± 9353 b | 180,915 ± 5506 a | |
| F5 | Rutin | 9,012,800 ± 178,088 b | 4,324,298 ± 458,177 c | 10,174,290 ± 289,308 a | |
| Glycosides and Glucosylated Compounds | G1 | Kiwiionoside | 833,782 ± 31,998 a | 591,256 ± 96,070 b | 544,706 ± 19,783 b |
| G2 | Neryl arabinofuranosyl-glucoside | 293,059 ± 44,975 b | 1,559,538 ± 104,179 a | 364,621 ± 18,820 b | |
| G3 | Vanilloyl glucose | 1,100,944 ± 183,073 a | 46,613 ± 5633 b | 105,084 ± 60,922 b | |
| G4 | Zizybeoside I | Not detected b | 77,605 ± 4379 a | 77,824 ± 301 a | |
| G5 | 3-Hydroxy-beta-ionol 3-[glucosyl-(1->6)-glucoside] | Not detected c | 128,088 ± 2514 a | 14,932 ± 3828 b |
| Cultivar | Pedigree | Self-Compatibility | Sharka Resistance (PPV-D) | Flowering | Ripening | Skin Color |
|---|---|---|---|---|---|---|
| ‘Bergeron’ | Unknown | Self-compatible | No | Late | Late | Light Orange |
| ‘Currot’ | Unknown | Self-compatible | No | Early | Early | Light Yellow/Pink |
| ‘Goldrich’ | ‘Sunglo × Perfection’ | Self-incompatible | Yes | Late | Late | Orange |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortuño-Hernández, G.; Silva, M.; Toledo, R.; Ramos, H.; Reis-Mendes, A.; Ruiz, D.; Martínez-Gómez, P.; Ferreira, I.M.P.L.V.O.; Salazar, J.A. Nutraceutical Profile Characterization in Apricot (Prunus armeniaca L.) Fruits. Plants 2025, 14, 1000. https://doi.org/10.3390/plants14071000
Ortuño-Hernández G, Silva M, Toledo R, Ramos H, Reis-Mendes A, Ruiz D, Martínez-Gómez P, Ferreira IMPLVO, Salazar JA. Nutraceutical Profile Characterization in Apricot (Prunus armeniaca L.) Fruits. Plants. 2025; 14(7):1000. https://doi.org/10.3390/plants14071000
Chicago/Turabian StyleOrtuño-Hernández, Germán, Marta Silva, Rosa Toledo, Helena Ramos, Ana Reis-Mendes, David Ruiz, Pedro Martínez-Gómez, Isabel M. P. L. V. O. Ferreira, and Juan Alfonso Salazar. 2025. "Nutraceutical Profile Characterization in Apricot (Prunus armeniaca L.) Fruits" Plants 14, no. 7: 1000. https://doi.org/10.3390/plants14071000
APA StyleOrtuño-Hernández, G., Silva, M., Toledo, R., Ramos, H., Reis-Mendes, A., Ruiz, D., Martínez-Gómez, P., Ferreira, I. M. P. L. V. O., & Salazar, J. A. (2025). Nutraceutical Profile Characterization in Apricot (Prunus armeniaca L.) Fruits. Plants, 14(7), 1000. https://doi.org/10.3390/plants14071000