Abstract
Most fir species in the Mediterranean have small to medium-sized distributions, are often endemic and endangered, and are mainly found in relict areas, except for Abies alba. The IUCN Red List of Threatened Species identified Abies nebrodensis as the rarest conifer in the world, with only 30 adult trees remaining. Additionally, Abies pinsapo is threatened and limited to five fragmented locations in Spain and Morocco. This study aimed to characterize the seed terpene profiles of Mediterranean Abies species, such as A. nebrodensis, A. pinsapo, and A. alba, since morphological results showed minimal variation among the Abies populations examined. Terpenes were extracted using n-heptane and then analyzed by GC-MS. The chemical composition revealed the dominance of limonene and α-pinene as the main monoterpenes in all the species, while A. nebrodensis reported the considerable presence of germacrene D-4-ol and selina-6-en-4-ol as sesquiterpenes. The relative contents of most of the terpenes were significantly different among the species, and subsequent statistical multivariate analysis showed clear discrimination among three distinct groups. These results confirmed the suitability of the terpene profile as a potential tool to study chemotaxonomic differences between species from the same family. Moreover, the compounds identified can be interesting for further studies on plant defense against biotic stress to reduce the risk of species extinction caused by pests and diseases.
1. Introduction
Conifers are the most important source of industrial wood, supplying over 50% of the world’s timber harvest [1]. Among them, the genus Abies Miller is the second largest genus in the Pinaceae family after Pinus [2] and is considered of major ecological and economic importance for coniferous forests. [3]. It is represented by approximately 50 species [4], and is mainly distributed in northern temperate regions of Asia, Europe, North and Central America, and North Africa [5]. Several Abies species are located in the mountains of the Mediterranean region while the others occupy only limited areas [6]. Based on the morphological and anatomical data and the geographic distribution, Mediterranean Abies species are grouped into two sections [7]. The Piceaster section features A. pinsapo Boiss. in southern Spain, A. numidica Carr. in Northwest Algeria, and A. marocana Trab. and A. tazaotana Villar in Northwest Morocco, while the Abies section includes A. alba Miller and A. nebrodensis (Loyac.) Mattei in Sicily [8].
Many fir species in the Mediterranean have limited geographical ranges, are often endemic and endangered, and are found in relict areas. However, A. alba [9] is an exception. It is the most widely distributed coniferous species in the central European forests [10]. This evergreen tree, also known as silver fir [11], can grow up to 40–50 m tall and produces cones that are 9–17 cm long and 3–4 cm wide, with about 150–200 scales [12].
Among the threatened species is Abies pinsapo Boiss. (Spanish fir). A relict fir endemic to Mediterranean mountain ranges in Spain and Morocco [13], it mainly exists in a limited area at five sites, three in southern Spain and two in the north of Morocco [14], at an altitude of 500–2000 m [15]. Spanish fir is found in about 3600 ha in south Spain, while it covers about 3000 ha in northern Morocco [16]. Over the centuries, this species has been intensely damaged due to unsustainable practices such as overgrazing, uncontrolled logging, and pollarding [17]. Thus, it is listed as an endangered tree by the International Union for Conservation of Nature (IUCN, https://www.iucnredlist.org/fr/species/42295/10679577; accessed on 24 February 2025), and forms pure woodlands that have special ecological importance for many organisms [18].
According to the IUCN Red List of Threatened Species, A. nebrodensis (Lojac.) Mattei, also known as Sicilian fir, is another conifer species and is classified as the most endangered tree in the Mediterranean region. Currently, only 30 trees remain in the Madonie Regional Park located in Northern Sicily, Italy; this endemic species is limited to an area of 84 ha, at an altitude of 1400–1650 m above sea level [19], although in the past it had a wider distribution [20]. The species is at heightened risk due to high genetic erosion, slow growth (only 50 cm in height over 10 years), and poor natural regeneration. Though it produces good seeds every 3–4, many of these seeds are empty, which leads to a low germination rate. Also, excessive grazing by wild herbivores is considered a serious problem for the growth of both adult trees and their new natural regeneration [21].
Conifers have evolved many physical and chemical defenses [22]. According to Lewinsohn et al. [23], terpenes are conifers’ most significant chemical defenses of a volatile nature, and they largely affect insect orientation and herbivory, as noted by Lundborg et al. [24]. Plants produce these substances as secondary metabolites and have a crucial protective role against stress, herbivores, and pathogenic microorganisms [25]. Forests are recognized as a major source of terpenes [26]; in particular, these volatiles are abundant and diverse in the family Pinaceae [27]. Their content and type are distinctive to each species and even vary between individual trees [28,29], and so terpenes can be a powerful tool for studying chemotaxonomic differences between species from the same family or genus [30]. Indeed, the relative percentages of constitutive terpene in mature tissue are often used in chemosystematic studies to characterize species, provenances, and clones since these compounds are under strong genetic control and are altered little by abiotic factors [31,32,33]. Further, these substances are used as ecological and biochemical markers in many coniferous taxa [34,35,36,37] to evaluate species-specific composition responses to the environment [38], to estimate the geographic variability of conifer species [39,40], and to determine trees’ resistance to pests or diseases because they are involved in the defense mechanisms of plants against insects and fungal infestations [41].
The relationships between the quantity and composition of terpenes were largely studied in coniferous species on different parts of the plant [37,38,39,40,42], and less often in seeds, to explain the chemotypes of and chemical variation in terpenes in trees of different geographical origins. In the literature, there is no information on the terpene profiles isolated from empty seeds collected from critically endangered Sicilian fir, endangered Spanish fir, and the common European silver fir species.
Therefore, this study aimed to study morphological seed features and characterize variation in seed terpene profiles of different Abies species grown in Mediterranean areas such as A. nebrodensis, A. alba, and A. pinsapo.
2. Results and Discussion
2.1. Seed Characteristics
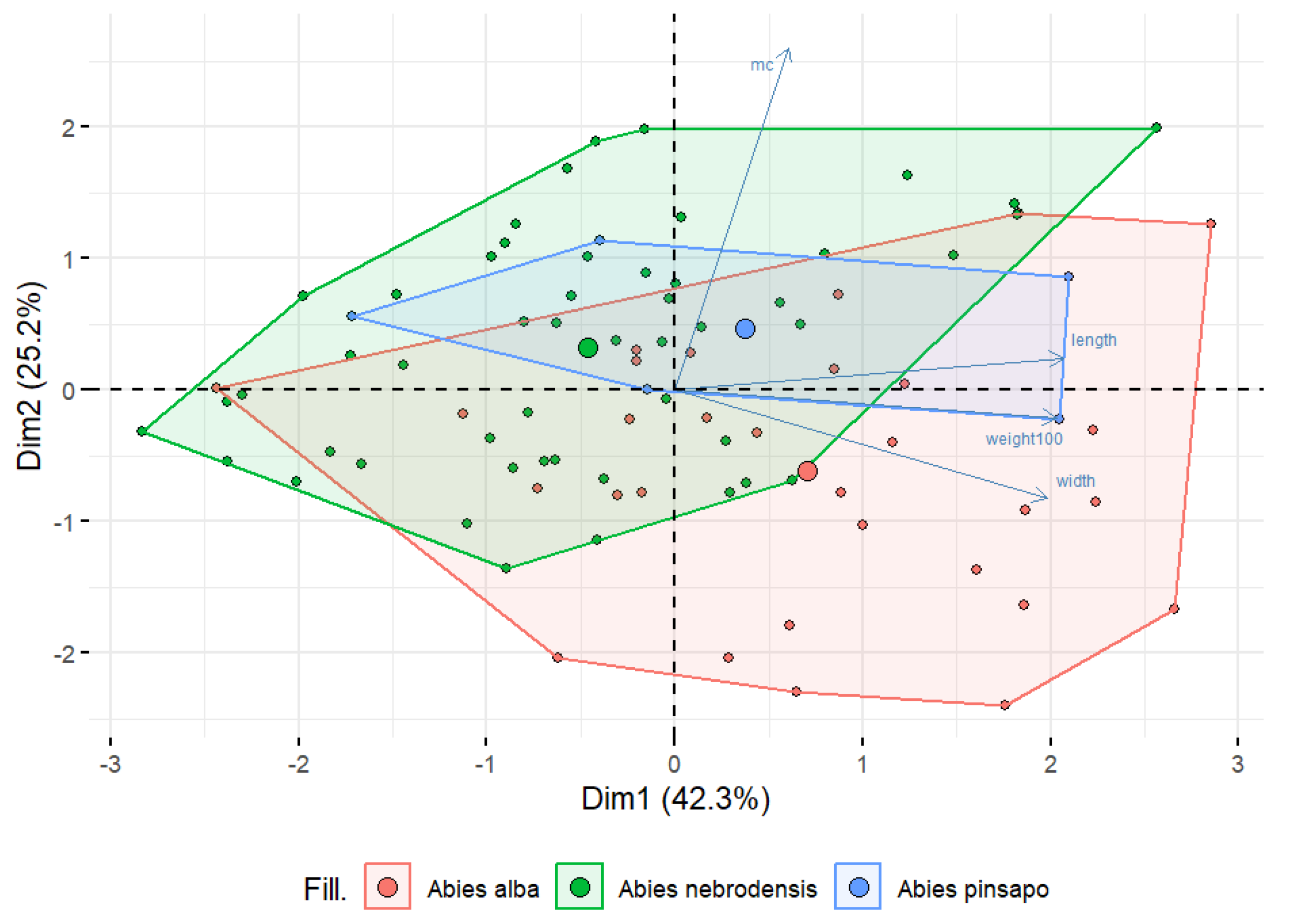
Seed features play a crucial role in species identification, classification, and understanding ecological adaptations [43]. Therefore, the moisture content (MC) percentage, 100-seed weight, length, and width of seeds from A. nebrodensis (AN: 6, 7, 8, 10, 12, 13, 19, 21, 22, and 27), A. alba (AA: 39, 64, 70, 106, and 120), and A. pinsapo (AP42) (Figure 1) were measured. In the present study, all the seed characters studied revealed significant variations among the species (Table 1). The highest coefficient of variation was recorded for moisture content (26.85%), followed by 100-seed weight (18.51%), seed width (15.10%), and seed length (9.12%).

Figure 1.
Representative empty seeds of A. nebrodensis, A. pinsapo, and A. alba. The numbers are the identification (ID) numbers of seeds collected from each population.

Table 1.
Moisture content (%) and morphological traits of empty seeds from different populations of Abies species.
The results indicated a highly significant difference in moisture content among the three Abies species (p < 0.001). The moisture content ranged from 6.66 ± 3.74% in A. alba to 18.99 ± 3.35% in A. nebrodensis (Table 1). The percentage of MC is an important seed trait that affects viability, desiccation tolerance, and storage potential. Seeds collected from Sicilian fir populations exhibited the largest variation, with MC ranging from 7.20 ± 1.61% to 18.99 ± 3.35%. In contrast, seeds from silver fir showed less variability, with moisture contents ranging from 6.66 ± 3.74% to 13.47 ± 6.42%. The significantly higher MC percentage found in A. nebrodensis compared to the other two species may indicate differences in seed desiccation tolerance, potentially influencing dormancy and germination strategies.
The seed dimensions, including length and width, that were recorded in A. nebrodensis showed significant variability, with lengths ranging from 8.7 ± 0.3 to 12 ± 1.2 mm and a width ranging from 0.49 ± 0.05 to 0.65 ± 0.1 mm (Table 1). The longest seeds were measured for A. nebrodensis. A. alba exhibited the widest seeds, while intermediate values of 10.8 mm in length and 0.59 mm in width were found in A. pinsapo. The differences in seed dimensions noted in our study are consistent with the findings by Singh et al. [44] for various populations of P. kesiya. Also, Hodžić et al. [45] indicated that there was variation among southern and northern populations of P. heldreichii, reporting 5.6 mm to 7.5 mm seed lengths and 3.0 mm to 4.4 mm seed widths. Mustafa et al. [46] reported seed widths between 3.07 mm and 3.71 mm in P. halepensis, a significantly larger size range than in Abies, but with similarly high intraspecific variation. Comparatively, Ghimire et al. [47] noted that A. koreana seeds measured 5.57 ± 0.35 mm in length and 2.92 ± 0.22 mm in width, while A. holophylla seeds were approximately 1 cm in length and 5 mm in width. These comparisons illustrate the extensive range of seed sizes among genotypes [48] and shapes across different species and provenances [49]. Since the seeds were collected from various locations, variances observed in seed parameters may be attributed to genetic variability and environmental conditions that occurred during seed development [50].
Significant variations were observed in the weight of 100 seeds from the investigated Abies species (Table 1). A. alba had the highest average weight at 6.42 ± 1.41 g, while the lowest weight of 2.93 ± 0.07 g was recorded in A. nebrodensis. A similar finding was noted by Skrzyszewska and Chłanda [49], who reported that seed weights for A. alba ranged from 38.92 g to 53.27 g per 1000 seeds collected from various provenances. Also, the seed weight was more influenced by individual trees, as noted by Trujillo-Ríos et al. [51]. While seed weight is commonly used in taxonomy and ecological studies due to its correlation with reproductive strategies and environmental adaptation, our results indicated no significant intraspecific or interspecific variation (Table 1).
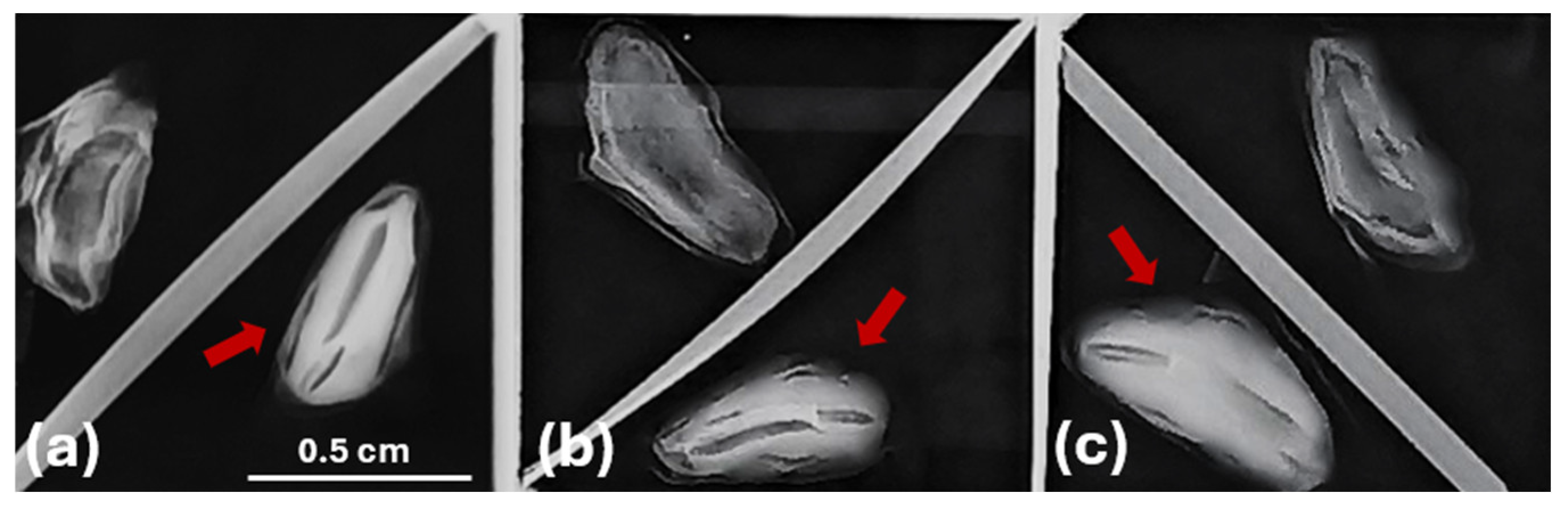
X-ray radiography techniques are used to estimate seed quality, mainly by assessing the development stages of seeds [49]. Their application in this study supports the selection between fully developed seeds, containing a well-formed embryo and endosperm, and empty (Figure 2) ones [52]. Such insights into internal seed characteristics are crucial for assessing seed quality and understanding the variability in seed traits observed among the populations from the three Abies species.
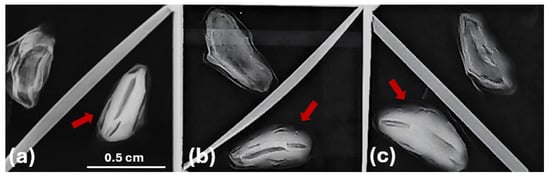
Figure 2.
X-ray image analysis of empty and full seeds of Abies species, with seeds containing zygotic embryos indicated by red arrows: (a) A. nebrodensis, (b) A. alba, (c) A. pinsapo.
To compare the differences among seeds from the three Abies species, morphological features and moisture content were analyzed using PCA, as shown in Figure 3. PCA was performed on a dataset of 17 Abies populations and 4 different variables to reduce the data dimensionality and to reveal the potential relationships among the parameters examined. The results showed that the first two PCs accounted for 76.5% of the total variability, explaining 42.3% for PC1 and 25.2% for PC2. However, there was overlap and a close distance among the populations, and so the tested features could not completely distinguish A. alba, A. nebrodensis, and A. pinsapo. From Figure 3, it can be seen that most of the samples of A. alba compared with the others were located on the right side of the biplot, which was positively correlated with the weight of the 100 seeds, length, and width, indicating that A. alba had bigger and heavier seeds than those from A. nebrodensis and A. pinsapo. Conversely, the major samples of A. nebrodensis were separated and located on the left side of the biplot, representing a negative correlation with all the morphological features. These observations confirm the existence of overlap among all the populations and make distinguishing seeds of one species from another quite difficult.
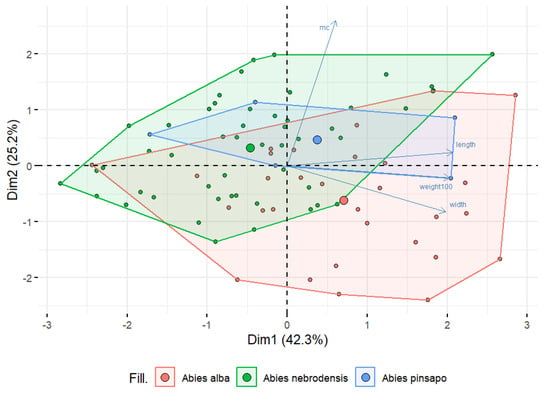
Figure 3.
A biplot of principal component analysis based on moisture content (%) and morphological characteristics in the populations of the three Abies species studied.
The morphological results indicated the existence of minimal variation among the Abies populations examined. In conifers, there is no external difference between empty and full seeds, although they have different weights [53]. Therefore, a further investigation into the chemical composition was conducted to rebut the similarity between seeds from various populations, since terpenes are known to correlate with geographic distributions [54].
2.2. Terpene Profile
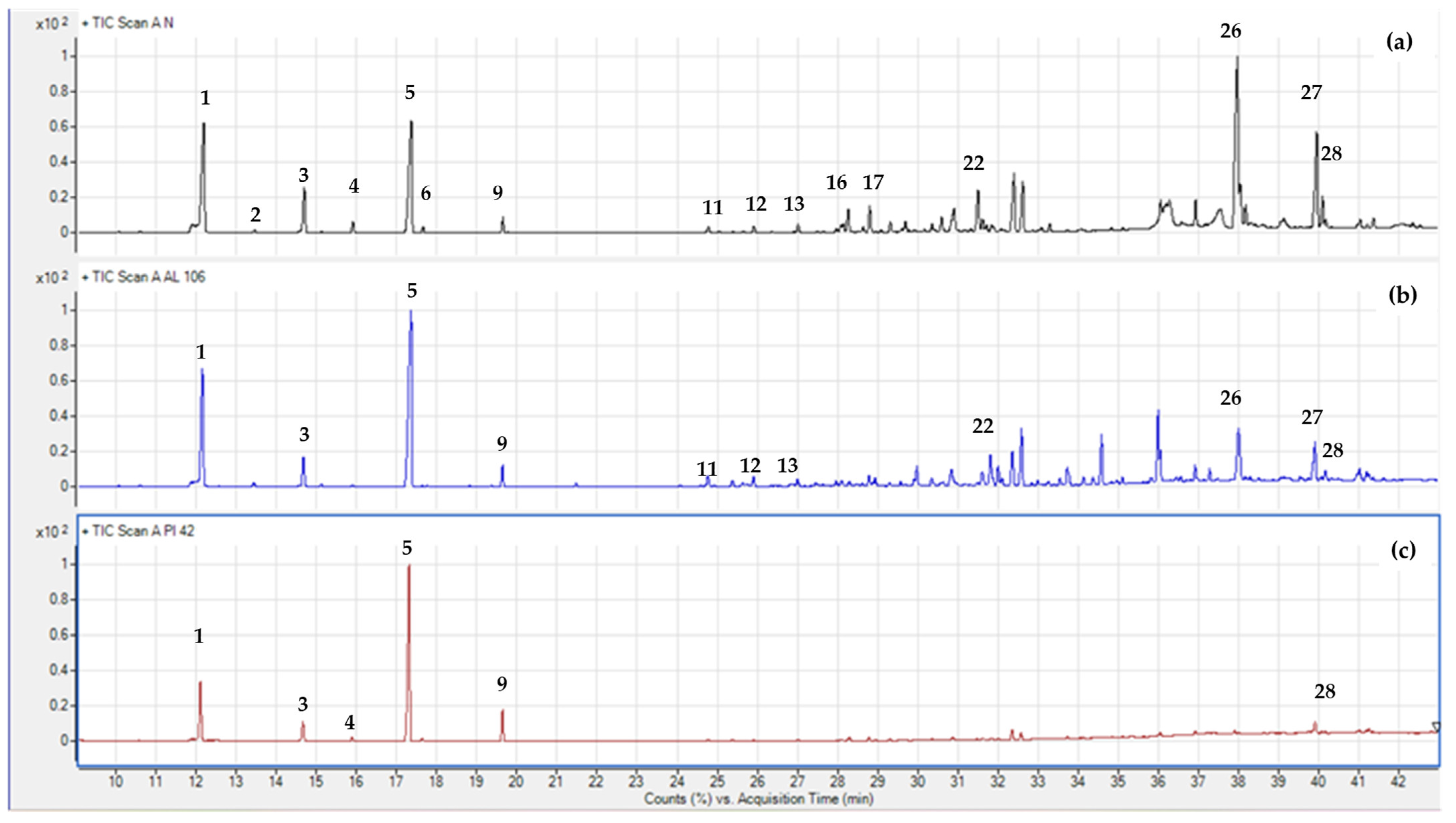
In this work, three Abies species grown in the Mediterranean basin were analyzed by GC-MS in order to characterize the overall terpene profile. Table 2 lists the volatile compounds, given in percent (%) of the total mass area (% area pct), as mean ± standard error values. In total, 28 compounds were classified into monoterpenes hydrocarbons, oxygenated monoterpenes, sesquiterpene hydrocarbons, and oxygenated sesquiterpenes. Figure 4 shows three gas chromatograms of A. nebrodensis (Sicilian fir), A. pinsapo (Spanish fir), and A. alba (silver fir), indicating some characteristic volatiles with numbers. According to the mean values of terpenes in each species (Table 2), the most dominant classes in all analyzed species were monoterpene hydrocarbons [55]. There was also a considerable presence of sesquiterpene hydrocarbons, but only in the case of A. nebrodensis. The highest percentage of monoterpenes hydrocarbons was noted in A. pinsapo (92.53%) and A. alba (82.95%), while A. nebrodensis reported the lowest presence (66.52%) in favor of sesquiterpene compounds. This result concurs with the work of Wajs-Bonikowska et al. [12] and Schicchi et al. [56], where monoterpene hydrocarbons in the seeds of A. alba and leaves of A., respectively, were the most dominant chemical group. The next most represented terpene classes in A. nebrodensis were sesquiterpene hydrocarbons (20.59%) and oxygenated sesquiterpenes (10.61%), while the least present were oxygenated monoterpenes, found in either A. pinsapo (1.14%) or A. alba (4.47%).
Table 2.
The terpene compounds of three Abies species, their mean relative percentage with SE and significance (p), and outcomes of Kruskal–Wallis test.
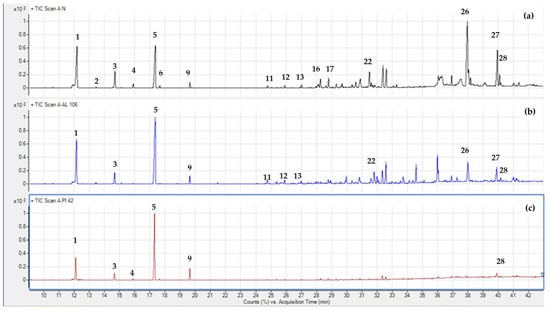
Figure 4.
Gas chromatograms of A. nebrodensis (a), A. pinsapo (b), and A. alba (c). Compounds were identified by retention indices, mass spectra (NIST library), and standard comparisons. The compounds identified were (1) α-pinene; (2) camphene; (3) β-pinene; (4) myrcene; (5) limonene; (6) β-phellandrene; (9) terpinolene; (11) α-cubebene; (12) α-copaene; (13) β-copaene; (16) β-elemene; (17) terpinen-4-ol; (22) germacrene D; (26) germacrene D-4-ol; (27) α-epi-cadinol; (28) selina-6-en-4-ol.
The seeds from all Abies species are characterized by the major presence of individual monoterpenes and dominated by a large proportion of monoterpenes [42]. Table 2 shows that the seeds contained the following monoterpenes: α-pinene, β-pinene, myrcene, limonene, and trans-verbenol. The most expressed monoterpene in Abies seeds was limonene, followed by α-pinene; however, the abundance of these monoterpenes differed noticeably among species.
This study is the first report on the composition of empty seeds from some endangered Mediterranean firs. In A. nebrodensis, the major compound was limonene (49.57%), while limonene and α-pinene represented the most abundant volatiles in A. pinsapo (66.03 and 18.85%, respectively) and A. alba (62.42 and 15.43%, respectively). The presence of sesquiterpenes was limited to germacrene D-4-ol (9.27%) and selina-6-en-4-ol (9.83%) in A. nebrodensis, representing 19.10% of the total identified compounds. For A. nebrodensis, previously published results by Schicchi et al. [56] on the leaves stated that other sesquiterpene hydrocarbons like longifolene (0.1%) and δ-cadinene (<0.05%) were present in minimal amounts. In addition, β-pinene was another monoterpene compound detected in all three fir taxa, with expression ranging from 5.89% (A. pinsapo) to 3.05% (A. nebrodensis). Thus, the phytochemical profile of each species could be recognized as follows:
A. nebrodensis: limonene > α-pinene > selina-6-en-4-ol > germacrene-D-4-ol, representing 79.76% of the total terpenes.
A. pinsapo: limonene > α-pinene > β-pinene, representing 90.76% of the total known terpenes.
A. alba: limonene > α-pinene, representing 77.85% of the total terpenes.
The present research has identified limonene and α-pinene as the primary compounds in the seeds of A. nebrodensis. A previous study by Schicchi et al. [56] examined the chemical composition of the leaves and reported the absence of limonene in favor of a higher content of β-pinene (48.8%), followed by α-pinene (17.5%). The authors attributed the lack of limonene in the leaf profile to genetic factors rather than environmental conditions. Among the volatile compounds found in the seeds (Table 2), camphene was detected in a much lower concentration (0.24%), while in the essential oil from the leaves, camphene was the third most dominant terpene, with 12.7% [56]. This highlights a notable difference in terpene profiles between the seeds and leaves of A. nebrodensis.
Our results demonstrated that the most abundant terpenes in the seeds of silver fir were limonene (62.42%) and α-pinene (15.43%), representing 77.85% of the total identified compounds, whilst Wajs-Bonikowska et al. [12] only reported limonene (>70%) as the major volatile in the seeds of this species from Poland. Similarly, limonene dominated the chemical profile of Italian A. alba seeds, representing >75% of the total identified compounds [57]. The empty seeds of A. alba contained considerable concentrations of β-pinene and verbenone [10], at 3.21 and 2.16% (Table 2). Additionally, selina-6-en-4-ol was another terpene found in silver fir seeds (Table 2), with a small concentration of 3.16%. This finding differed from a previous study by Wajs-Bonikowska et al. [10], which reported a significantly higher percentage (51.7%) of this compound in an A. alba seed hydrolate.
A. pinsapo was another species investigated in this study. Its chemical seed profile was dominated by 66.03% limonene and 18.85% α-pinene, followed by 5.89% β-pinene and 2.55% selina-6-en-4-ol (Table 2). These are the first published data on the terpene profile of Spanish fir seeds, considering that all previous studies widely focused on the phytochemical composition of the wood. Barrero et al. [58] isolated the first sesquiterpenoids from the acid fraction of the hexane extract, while later Barrero et al. [59] obtained α-cubebene, longifolene, α-murolene, and δ-cadinene from the neutral fraction and γ-cadinene and germacrene D via the usual spectroscopic techniques. Among these compounds, only α-cubebene and germacrene D appeared as traces (0.18 and 0.42%, respectively) in this study (Table 2).
According to the literature, the primary terpenes identified in Abies species include α-pinene, β-pinene, camphene, limonene, and δ-3-carene [60]. However, the terpene compositions of the seeds of A. nebrodensis and A. pinsapo, growing in their natural habitats, have not been studied. In this paper, the most prevalent compounds found were limonene and α-pinene, while the amounts of β-pinene and camphene varied depending on the tested specific species. It was noted that santene and tricyclene [56], as known compounds of A. nebrodensis, as well as bornyl acetate [61], found in A. pinsapo, and δ-3-carene [62], found in A. alba, were completely absent from the chemical profiles of seeds (Table 2). Compared to the current study, Kshatriya et al. revealed distinct monoterpenes in the seeds of other Abies species [42]. α-pinene and β-myrcene, found in A. amabilis, α-pinene and β-pinene, found in A. balsamea, limonene, found in A. grandis, and β-phellandrene, found in A. lasiocarpa, were the most predominant compounds. Such variation in the chemical profiles in conifer terpenes is attributed to the complex biosynthetic systems encoded by a large number of terpene synthase (TPS) genes and known terpene-modifying cytochrome (P450) [63,64]. Thus, genetic relationships between populations can be illustrated based on the content of monoterpenes and sesquiterpenes [65].
The highest number of terpenes was found in silver fir (28), followed by Sicilian fir (27) and Spanish fir (24), and many common compounds were identified in varying ratios (Table 2). Indeed, statistical analysis revealed variation in the mean values of the detected compounds. The different terpene distributions found among the three Abies species were verified with the Kruskal–Wallis test, as a nonparametric method, to check if multiple groups followed the same distribution. In most cases, there were notable differences (p < 0.05, p < 0.01, and p < 0.001) in the proportions of volatile compounds according to the fir species (Table 2).
Statistically significant differences were found in most of the detected monoterpenes and sesquiterpenes, and only 3 (α-pinene, limonene, and boranyl acetate) out of 25 compounds showed no statistically differences between species (Table 2). For example, the contents of germacrene D-4-ol (9.27%) and selina-6-en-4-ol (9.83%) in A. nebrodensis were greater compared to A. pinsapo (0.16 and 2.55%, respectively) and A. alba (0.69 and 3.16%, respectively), whereas verbenone was more abundant in A. alba (2.16%) than in A. nebrodensis (0.43%) and A. pinsapo (0.01%).
To determine which groups contribute to significant differences, we employed Dunn’s multiple comparisons as the post hoc procedure for each statistically significant variable. Dunn’s tests showed the highest number of nineteen differing compounds between A. nebrodensis and A. alba, whereas there were sixteen differing compounds each between A. nebrodensis and A. pinsapo, as well as between A. alba and A. pinsapo (Table 3). The most frequent terpenes that showed significant differences between each analyzed pair species were α-pinene, camphene, myrcene, p-cymene, verbenone, and α-copaene. However, terpinolene, bornyl acetate, and trans-verbenol were the only terpenes with no significantly different values for the three analyzed groups.
Table 3.
Terpene compounds revealed statistically significant differences (indicated with asterisks) between Abies species (* p < 0.05, ** p < 0.01, *** p < 0.001); p values from post hoc analysis with Dunn’s multiple comparisons test (n = 2).
Terpene profiles were studied as biochemical markers in coniferous trees from various geographical origins to define chemotypes and explain their diversity [66]. Indeed, notable variation is reported in species from the genus Abies, corresponding to whether the individuals are grown naturally in their native habitat or artificially cultivated [60]. β-pinene was the most abundant compound in the natural populations of Abies species from Romania, Serbia, North Macedonia, and Bulgaria, followed by limonene-β- phellandrene [37] or α-pinene and camphene [67]. On the other hand, α-pinene was the main volatile in the A. cephalonica population from Greece [37,67]. Bornyl acetate was found in the artificial plantations of A. alba [61], and δ-3-carene prevailed in the naturally distributed A. nordmanniana from Turkey [62].
Many studies have demonstrated that monoterpenes are effective tools for chemotaxonomic surveys, helping to determine intraspecific variation and define certain species [35,37,38]. To overcome the challenges associated with classifying species based solely on morphological data, it was advised to use gas chromatography–mass spectrometry (GC-MS) techniques to identify potential terpene markers with multivariate analyses [68].
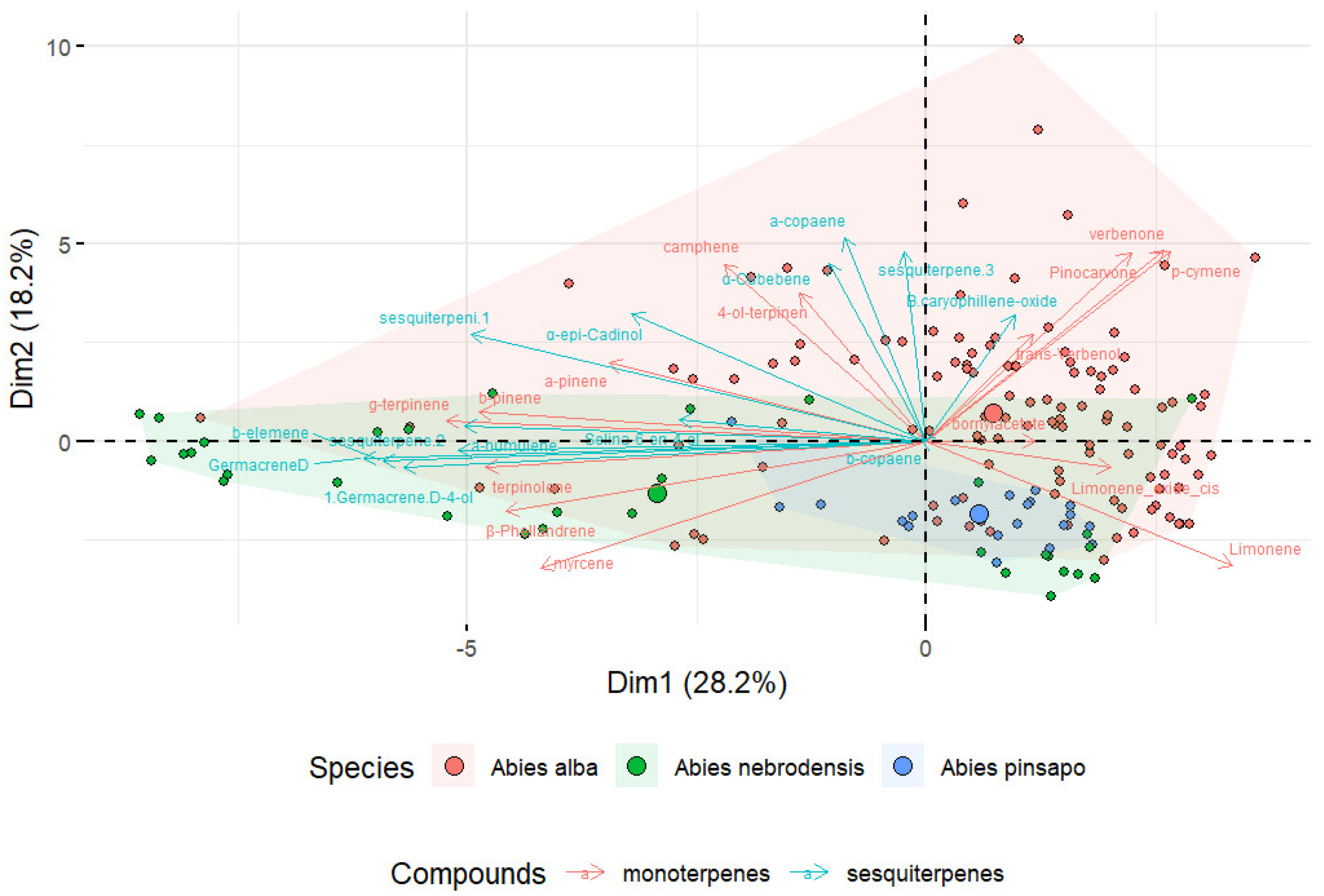
For this reason, PCA of matrix data was performed to investigate the variability among the three Abies species. The first and second components of the PCA of the collected terpenes explained 46.4% of the variation in the data, 28.2% of the variation in PC1, and 18.2% of the variation in PC2 (Figure 5). Moreover, as shown in the PCA biplot, the Abies had different terpenoid profiles that could be visually distinguished according to the species. The length of the arrow indicates the contribution of each compound to the first two components in the PCA. Longer arrows show a higher contribution by the variables, while shorter arrows denote a lower contribution by the variables. This is a useful tool for data reduction to visualize the similarities among complex datasets. An acceptable degree of separation was found between the samples from A. nebrodensis, A. alba, and A. pinsapo, suggesting that the different terpenes contribute to the distinctive terpene profile of Abies samples. Although the results of PCA depicted the separation of the Abies species encouraged to some extent, it is important to note that PCA only provides visual information, with no specific index to verify the actual differences.
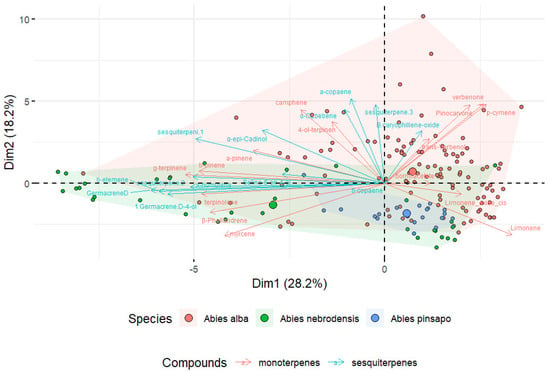
Figure 5.
Principal component analysis (PCA) biplot of three Abies species based on the variance in the terpene compounds.
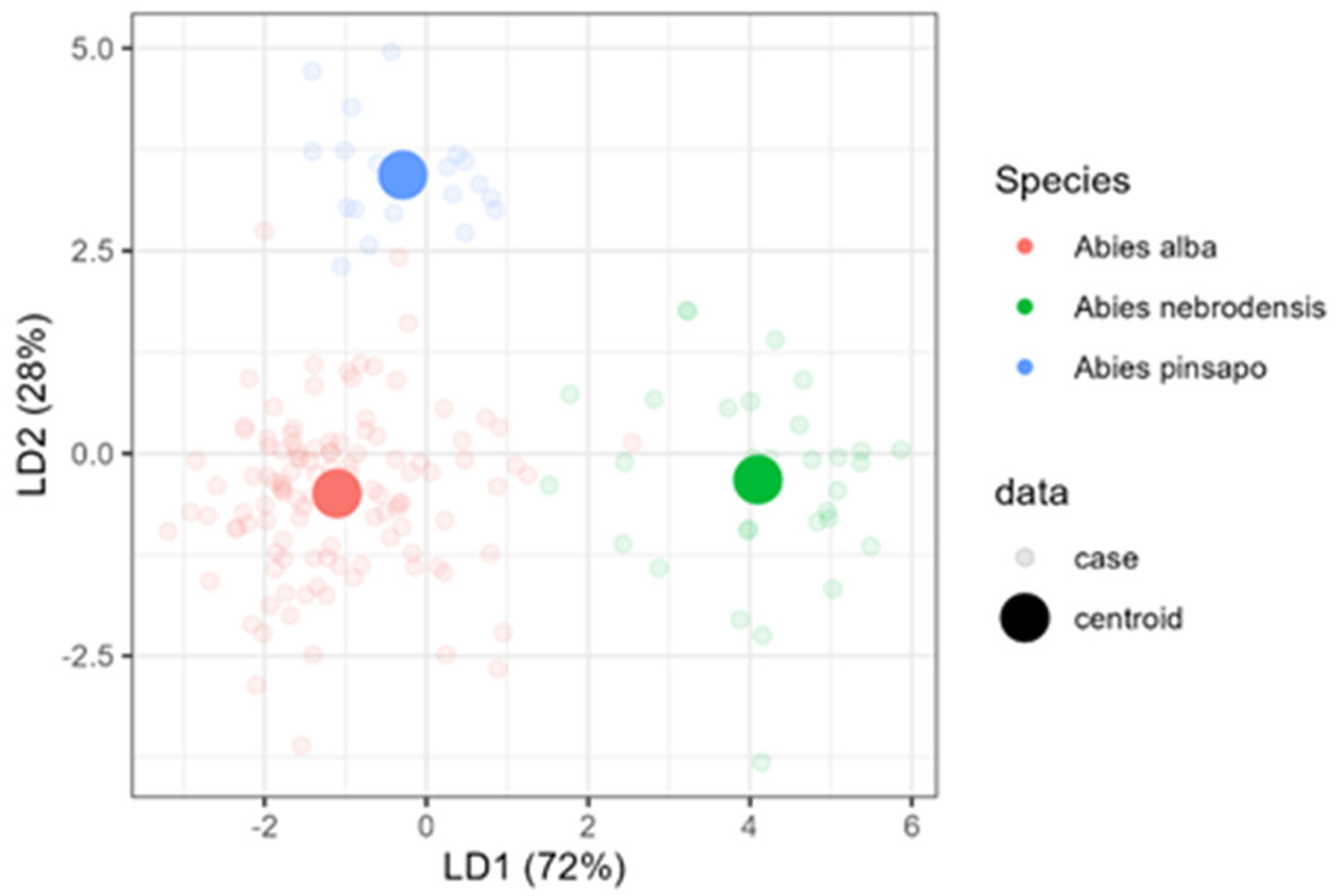
Therefore, to build a tool for Abies species classification using a relative terpene abundance, linear discriminant analysis (LDA) was used on the selected data matrix. The previous data were chosen to avoid collinearity, as mentioned in the Materials and Methods Section, and the subsequent compounds, β-elemene, germacrene D, and germacrene-D-4-ol, were excluded. In Figure 6, an LDA biplot of data with species centroids is presented. The LD1 and LD2 functions explained 96.30% of the total variance (Wilks’ λ = 0.086134, p < 2.2 × 10−16). LDA, unlike PCA, is a supervised ordination method that maximizes the separation among the data classes and minimizes distance within the class. LDA separated all three species well; indeed, A. alba and A. nebrodensis were discriminated by LD1, while LD2 enabled the separation of A. pinsapo from the other two species (Figure 6).
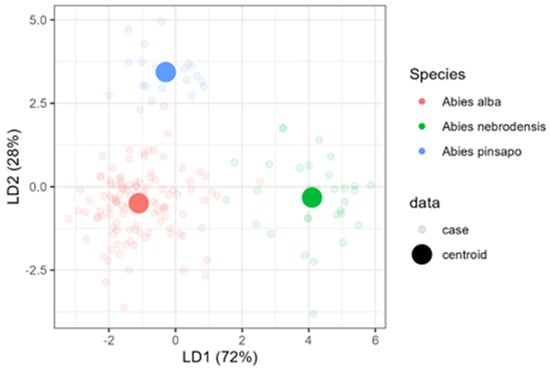
Figure 6.
Linear discriminant analysis (LDA) for the classification of three Abies species.
LDA exhibited more sharp classes among the 25 compounds. LDA was also convenient for isolating A. nebrodensis from others. A. pinsapo was also more isolated away from A. alba and A. nebrodensis. Only a few cases of A. alba seemed close to A. nebrodesis and A. pinsapo. The first two discriminant functions effectively discriminated between Abies species (Table 4); therefore, three groups representing each species were clearly observed. The first discriminant function explained 72% of the total variance and the variables with the greatest discriminant ability were sesquiterpene.1 (2.37) and p-cymene (2.11). The second discriminant function accounted for 28% of the data variation and was mainly correlated with p-cymene (1.85) and pinocarvone (1.79).
Table 4.
Coefficients of linear discriminants of Abies species in relation to terpene compounds. Significant coefficients are in bold.
Abies is one of the most explored genera for chemotaxonomic purposes [69]. Earlier, Otto and Wilde [70] marked the importance of terpene profiles to differentiate species and subspecies. In this context, α-pinene, β-pinene, bornyl acetate, and p-cymene are the most characteristic monoterpenes that may serve as chemosystematic markers in conifers. Thus, the separation of the three Abies species was confirmed by analyzing their terpene compositions. Not all compounds provide the same level of discrimination, but the most effective ones in differentiating between species include p-cymene and sesquiterpene.1, as well as terpinolene and pinocarvone. Our results indicate that sesquiterpenes could be more involved in species diversity than monoterpenes [71], even though their quantities are relatively low compared to the total identified composition.
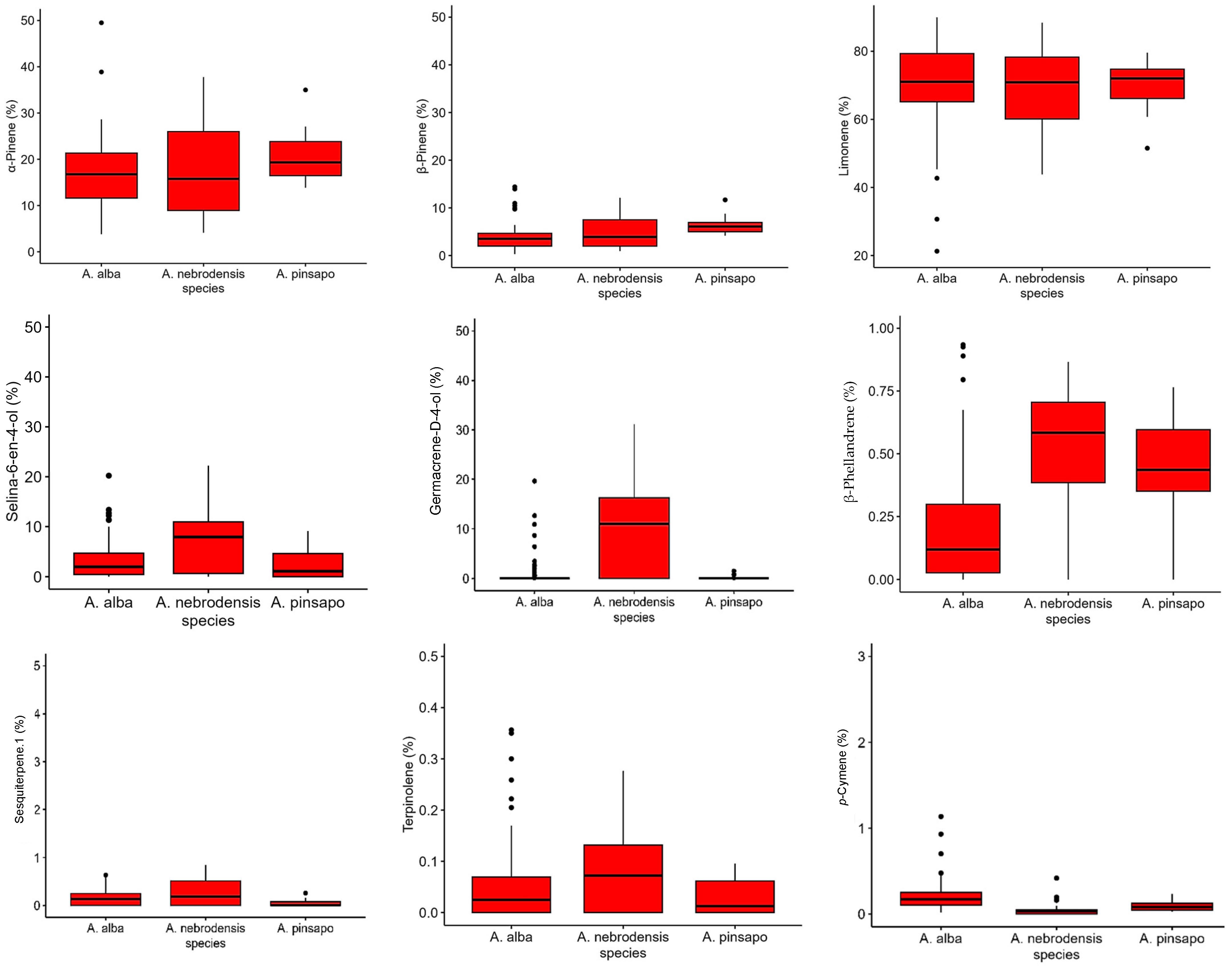
The box plots of the most important terpenes are shown in Figure 7. It was found that the relative contents of α-pinene, β-pinene, limonene, selina-6-en-4-ol, germacrene-D-4-ol, β-phellandrene, sesquiterpene.1, and terpinolene were higher in A. nebrodensis. The relative percentages of α-pinene and β-pinene in A. pinsapo were greater than those in A. alba. Moreover, the relative content of p-cymene was greater in A. alba than in other species, while the levels of sesquiterpene.1 was higher in A. alba and A. nebrodensis than A. pinsapo. The key biomarker for A. nebrodensis was germacrene-D-4-ol, and the key biomarker for A. alba was p-cymene.
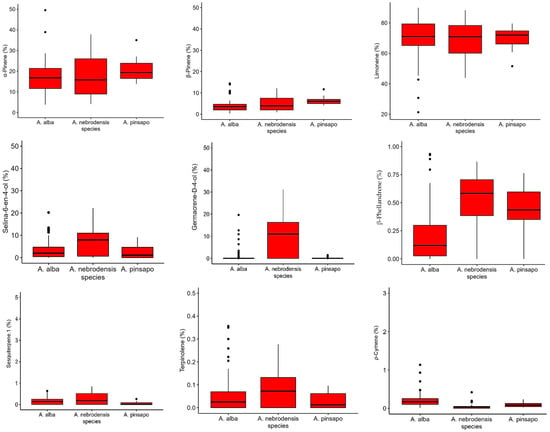
Figure 7.
The variation in the relative percentage of various terpenes detected in the empty seeds of A. nebrodensis, A. alba, and A. pinsapo grown in the Mediterranean basin.
Generally, longifolene, δ-cadinene, and α-humulene were among the sesquiterpenes most commonly used to discriminate Abies species, particularly those grown in the Mediterranean basin [65]. The comparison of our results with those of previous studies pointed out that not only could limonene and α-pinene be included in the chemotaxonomic research of the genus Abies, but so could others like camphene [72], β-pinene [73], β-phellandrene [74], and p-cymene [68].
To test the success of LDA in classifying each object in the group, classification accuracy was calculated by comparing the correctly classified samples to the total number of samples in each Abies species. In this context, the confusion matrix indicates that applying LDA to the complete dataset resulted in a recognition percentage of 96.97%. As shown in Table 5, LDA has acceptable classification accuracy for new samples regarding their geographical origins. The best results were reported for A. nebrodensis and A. pinsapo since all samples were properly classified with 100% accuracy. The classification accuracy of A. alba samples was acceptable (95.83%), as only one sample out of twenty-three was misclassified as an A. nebrodensis sample.
Table 5.
The confusion matrix of the results obtained for each species on the testing dataset using trained LDA classification model. Correct identifications are emphasized with bold font.
3. Materials and Methods
3.1. Plant Material
The current study was conducted on seeds from mature Abies spp.—trees. Seeds of Abies nebrodensis and Abies alba were collected from different sites, as detailed in Table 6, while those of Abies pinsapo were provided by the Department of Agriculture, Food, Environment and Forestry (DAGRI), University of Florence, Italy. All seeds were subjected to X-ray analysis to separate the full (with embryos) seeds from the empty seeds [52], and only the empty seeds were used for further investigation in this study.
Table 6.
Location and geographical characteristics of Mediterranean Abies spp. populations.
3.2. Seed Moisture Content and Morphological Features
After X-ray analysis, 25 empty seeds (with wings removed) were used for each tree for morphometric measurements. A stereomicroscope and camera system (Zeiss Stemi 2000 C, Jena, Germany) were used for size measurements, and ImageJ software (Rasband, W.S., ImageJ, U.S. National Institutes of Health, Bethesda, MD, USA) version 1.54g; java 1.8.0-172, was utilized to measure the width and length of the photographed seeds. Weight measurements were performed using a precision balance, with an accuracy of 0.0001 g. For the moisture content analysis of the seeds, 5 seeds (0.5 g) were used for each species, and their moisture content was determined with a Moisture Analyzer (Mettler-Toledo AG, Laboratory & Weighing Technologies, Greifensee, Switzerland). All the measured values, given in millimeters, are reported as the mean and standard deviation, as shown in Table 1.
3.3. Terpene Isolation
Terpenes were isolated from A. nebrodensis, A. alba, and A. pinsapo seeds. For each plant of Abies species, 20 empty seeds were individually analyzed. The extractions were performed in 2 mL glass vials containing one seed with 0.2 mL of sample, followed by sonication for 30 min (3 cycles of 10 min) using an Ultrasonic cleaner (5300i EP S3, Soltec, Milan, Italy). Subsequently, samples were placed on an incubating shaker (Thermoshake, Gerhardt, Milan, Italy) at a speed of 100 rpm for 24 h at 35 °C. The next day, a fraction of 100 µL of each seed extract was transferred to GC vials and stored at 4 °C until further analysis.
3.4. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis
Chemical analyses were carried out using an Agilent 7820A gas chromatograph coupled to an Agilent mass spectrometer 5977E (Agilent, Palo Alto, CA, USA) with a quadruple mass selective detector (electron ionization) equipped with a data processor: we employed Mass hunter Quantitative version B 07 01 and Qualitative version B 06 00. Terpenes were separated on a capillary column known as DB-Wax Ultra Inert (Agilent, 0.25 mm × 60 m × 0.5 µm film thicknesses). The following conditions were adopted: splitless (1 min); a flow of 1.2 mL min−1; and Helium 5.5 as a carrier gas. Overall, 1 µL of extract was injected using Gerstel MPS2 XL autosampler (Gerstel, Mülheim an der Ruhr, Germany); the injector temperature was set to 250; the initial temperature was 40 °C. It was programmed to 200 °C at 5 °C min−1, and then increased to 240 °C at 10 °C min−1 (held 5 min). The total run time was set to 44 min for each sample. The mass spectrometer operated with an electron ionization of 70 eV in scan mode in the m/z range of 29–330, at 4.5 scans s−1. The compounds were identified based on matching their mass spectra with those obtained from the NIST 11 (National Institute of Standards and Technology, Gaithersburg, MD, USA) library, using standards when possible, and the Kovats index.
3.5. Statistical Analysis
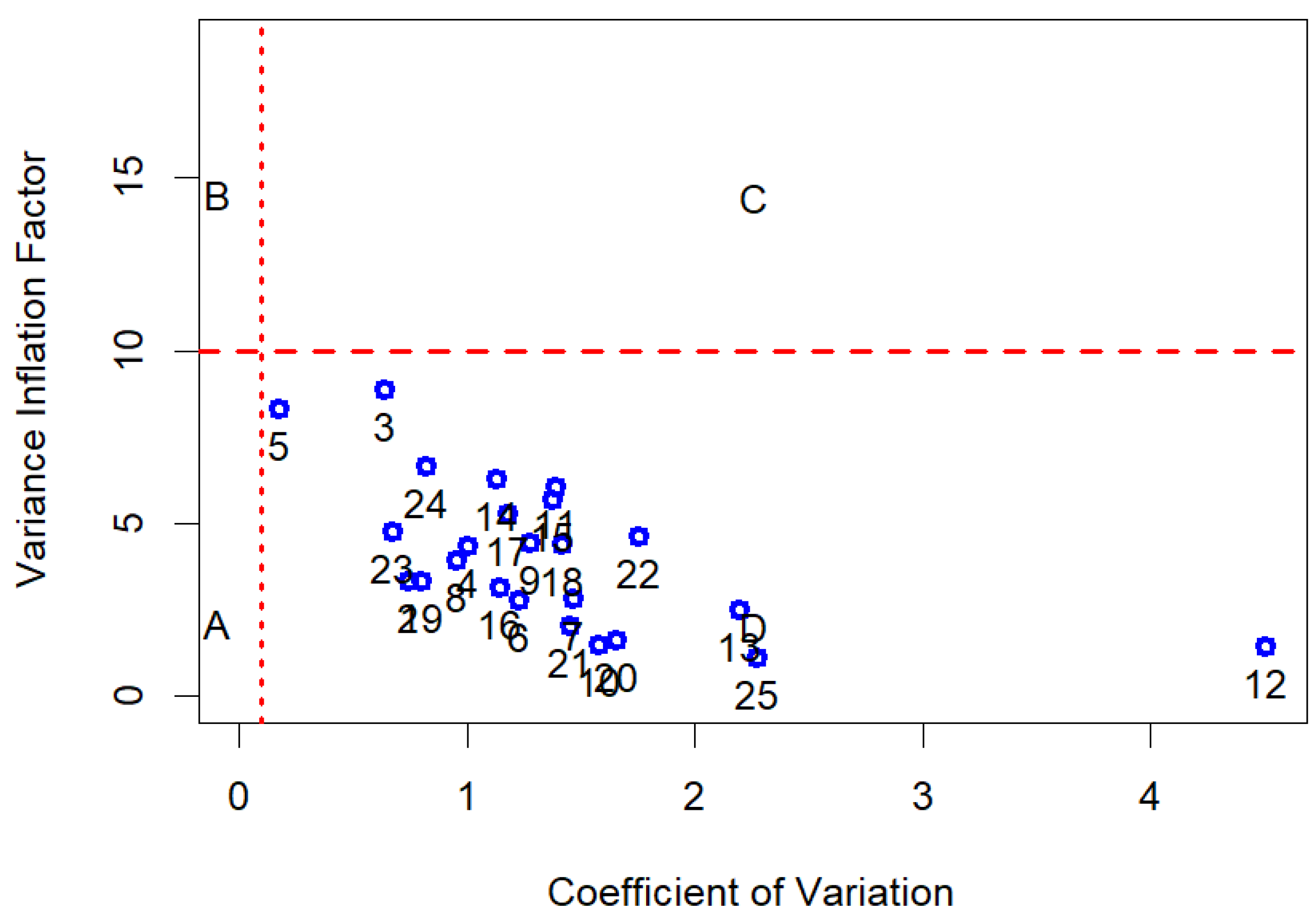
A one-way ANOVA test was performed for the moisture content (%) and morphological traits of empty seeds to determine significant differences in the means of the three Abies species. The statistical analysis of the data matrix (N = 130 observation for 28 terpenes) was carried out in 5 steps. (i) GC data were normalized as monoterpenes (C10) in relation to the total amount of monoterpene compounds, while sesquiterpenes (C15) were normalized on the sum of mono and sesquiterpenes. (ii) The second step was to verify normality, for each compound investigated, by using the Shapiro normality test [75] and the level of collinearity. The measure of the latter is given by the index of Variance Inflation Factor (VIF < 10, Figure 8) for each individual variable [76]. The variable selection to reduce collinearity was carried out using the “multiColl” R package [77] in order to perform the next step of linear discriminant analysis (LDA). (iii) The third step of the analytic framework was to evaluate if the grouping variable, represented by the tree species (A. alba, A. nebrodensis, and A. pinsapo), had shown a significant difference in each terpene’s relative content. The Kruskal–Wallis test was performed on the single terpene’s data and the related post hoc results were computed (Dunn test). The Kruskal–Wallis test is a non-parametric test, assumes no particular distribution of data, and is analogous to one-way analysis of variance (ANOVA). The results were presented in tabular form. (iv) A preliminary exploration of dataset variability was conducted effectively through PCA (principal component analysis). (v) Finally, a supervised classification was carried out using Fisher Linear Classification Analysis. LDA is a set of methods used in multivariate statistics to find a linear combination of features that characterize or separate two or more classes of objects [78] and is a tool for classification, dimension reduction, and data visualization. The training and test datasets needed to perform LDA were obtained by splitting the original dataset into two subsets thanks to the “createDataPartition” function of the R “caret” package [79]. The percentage of data used for training was equal to 80%, and the remaining data were used to test the classifier. The confusion matrix was performed on the test dataset. Overall statistics of classifier and accuracy statistics by species (Sensitivity or True Positive Rate, Specificity or False Negative Rate, the Detection Rate, and other classification errors) were presented. All analyses were conducted in the R statistical computing environment (version 4.3.0) by using the following R packages: we used “ggplot2” [80] for graphical purposes; we used “FactoMineR” [81] and “ordr” [82] for PCA and LDA plottings; “caret” to establish the accuracy statistics of an LDA classifier; “matrixTests” [83] provides the functions to test one-way stratification significance by species (the non-parametric Kruskal–Wallis test for terpene data and one-way ANOVA for seed data); and the “PMCMRplus” [84] and “postHoc” [85] were used for post hoc comparison. All data and codes are available at the web repository of the work: https://github.com/bioeconomy/mediterranean_fir_analisys; accessed on 24 February 2025).
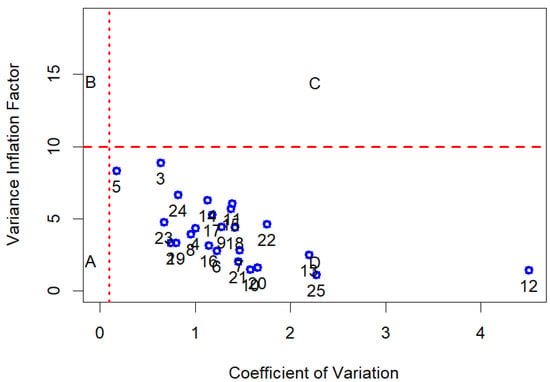
Figure 8.
The index of Variance Inflation Factor for each single variable. Letters are labels for the areas identified by VIF (<10) and coefficient of variation (CV = 0.1) indicating the contribution of each predictor to VIF.
4. Conclusions
The current study demonstrates variation in moisture content, the weight of 100 empty seeds, and seed dimensions among and within Abies populations. Although PCA results indicated overlapping among all the populations, making species distinction quite difficult, seed morphological traits can serve as reliable taxonomic markers in species identification and classification. These findings suggest the need for other work using additional parameters such as cone dimensions, the cone mass number of seeds per cone, etc., to establish the differences among and within the conifer species.
To address the difficulties of classifying species solely based on morphological data, chemical profiles obtained through GC/MS analysis, combined with multivariate techniques, proved effective for classifying and predicting the potential origin of unknown Abies samples. While the results of PCA indicated some separation among Abies species, its effectiveness for classification was limited by the absence of a specific index to confirm actual differences. In this context, LDA, as a supervised method, was particularly useful in separating the Abies species into A. nebrodensis, A. pinsapo, and A. alba.
The current findings from the analysis of seed terpene composition will enable further research on how different terpene profiles influence various biological processes. These processes include protecting seeds against pathogens, herbivores, and desiccation, as well as effects on seed viability and germination.
Author Contributions
Conceptualization, W.T., M.M. and A.C.; methodology, W.T., G.C. and M.M.; software, W.T., G.C. and A.C.; validation, W.T., C.B. and A.C.; formal analysis, A.C.; investigation, W.T., T.İ. and G.C.; resources, C.B., G.C. and M.M.; data curation, W.T., G.C., A.C.; writing—original draft preparation, W.T., T.İ., G.C., A.C.; writing—review and editing, W.T., T.İ., C.B., G.C., M.M. and A.C.; visualization, W.T., C.B., G.C. and A.C.; supervision, W.T.; M.M. and A.C.; project administration, A.C.; funding acquisition, A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors thank Alberto Pierguidi from Dipartimento di Scienze e Tecnologie Agrarie, Alimentari, Ambientali e Forestali, Università degli Studi di Firenze, for providing A. pinsapo seeds used in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cooper, R.J. World markets for coniferous forest products: Recent trends and future prospects. Acta Hortic. 2003, 615, 349–353. [Google Scholar] [CrossRef]
- Farjon, A. A Handbook of the World’s Conifers; BRILL: Leiden, The Netherlands, 2010; Volume 1. [Google Scholar]
- Liu, T.S. A Monograph of the Genus Abies; Department of Forestry, College of Agriculture, National Taiwan University: Taipei, China, 1971. [Google Scholar]
- Ran, J.H.; Shen, T.T.; Wu, H.; Gong, X.; Wang, X.Q. Phylogeny and evolutionary history of Pinaceae updated by transcriptomic analysis. Mol. Phylogenet. Evol. 2018, 129, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Padalia, R.C.; Verma, R.S.; Chauhan, A.; Goswami, P.; Chanotiya, C.S. Chemical analysis of volatile oils from West Himalayan Pindrow Fir Abies pindrow. Nat. Prod. Commun. 2014, 9, 1181–1184. [Google Scholar] [CrossRef]
- Sękiewicz, K.; Sękiewicz, M.; Jasińska, A.K.; Boratyńska, K.; Iszkuło, G.; Romo, A.; Boratyński, A. Morphological diversity and structure of West Mediterranean Abies species. Plant Biosyst. 2013, 147, 125–134. [Google Scholar] [CrossRef]
- Liepelt, S.; Mayland-Quellhorst, E.; Lahme, M.; Ziegenhagen, B. Contrasting geographical patterns of ancient and modern genetic lineages in Mediterranean Abies species. Plant Syst. Evol. 2010, 284, 141–151. [Google Scholar] [CrossRef]
- Terrab, A.; Talavera, S.; Arista, M.; Paun, O.; Stuessy, T.F.; Tremetsberger, K. Genetic diversity at chloroplast microsatellites (cpSSRs) and geographic structure in endangered West Mediterranean firs (Abies spp., Pinaceae). Taxon 2007, 56, 409–416. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W. Abies_spp. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the EU: Luxembourg, 2016; p. e015be7+. [Google Scholar]
- Wajs-Bonikowska, A.; Sienkiewicz, M.; Stobiecka, A.; Maciąg, A.; Szoka, Ł.; Karna, E. Chemical Composition and Biological Activity of Abies alba and A. Koreana Seed and Cone Essential Oils and Characterization of Their Seed Hydrolates. Chem. Biodivers. 2015, 12, 407–418. [Google Scholar] [CrossRef]
- Pokorska, O.; Dewulf, J.; Amelynck, C.; Schoon, N.; Šimpraga, M.; Steppe, K.; Van Langenhove, H. Isoprene and terpenoid emissions from Abies alba: Identification and emission rates under ambient conditions. Atmos. Environ. 2012, 59, 501–508. [Google Scholar] [CrossRef]
- Wajs, A.; Urbańska, J.; Zaleśkiewicz, E.; Bonikowski, R. Composition of essential oil from seeds and cones of Abies alba. Nat. Prod. Commun. 2010, 5, 1291–1294. [Google Scholar] [CrossRef]
- Peguero-Pina, J.J.; Sancho-Knapik, D.; Flexas, J.; Galmés, J.; Niinemets, Ü.; Gil-Pelegrín, E. Light acclimation of photosynthesis in two closely related firs (Abies pinsapo Boiss. and Abies alba Mill.): The role of leaf anatomy and mesophyll conductance to CO2. Tree Physiol. 2016, 36, 300–310. [Google Scholar] [CrossRef]
- Sanchez-Morales, J.; Pardo-Igúzquiza, E.; Rodríguez-Tovar, F.J.; Dowd, P.A. A new method for reconstructing past-climate trends using tree-ring data and kernel smoothing. Dendrochronologia 2019, 55, 1–15. [Google Scholar] [CrossRef]
- De Vita, P.; Serrano, M.S.; Luchi, N.; Capretti, P.; Trapero, A.; Sánchez, M.E. Susceptibility of Abies pinsapo and its tree cohort species to Heterobasidion abietinum. For. Pathol. 2010, 40, 129–132. [Google Scholar] [CrossRef]
- Linares, J.C.; Carreira, J.A. Temperate-like stand dynamics in relict Mediterranean-fir (Abies pinsapo, Boiss.) forests from southern Spain. Ann. For. Sci. 2009, 66, 610. [Google Scholar] [CrossRef]
- Linares, J.C.; Camarero, J.J.; Carreira, J.A. Interacting effects of changes in climate and forest cover on mortality and growth of the southernmost European fir forests. Glob. Ecol. Biogeogr. 2009, 18, 485–497. [Google Scholar] [CrossRef]
- Sánchez-Robles, J.M.; Balao, F.; García-Castaño, J.L.; Terrab, A.; Navarro-Sampedro, L.; Talavera, S. Nuclear microsatellite primers for the endangered relict fir, abies pinsapo (Pinaceae) and cross-amplification in related Mediterranean species. Int. J. Mol. Sci. 2012, 13, 14243–14250. [Google Scholar] [CrossRef] [PubMed]
- Frascella, A.; Della Rocca, G.; Barberini, S.; Emiliani, G.; Secci, S.; Lambardi, M.; Benelli, C.; Tarraf, W.; Izgu, T.; Schicchi, R.; et al. Innovative In Situ and Ex Situ Conservation Strategies of the Madonie Fir Abies nebrodensis. Sustainability 2022, 14, 12643. [Google Scholar] [CrossRef]
- Pasta, S.; Sala, G.; La Mantia, T.; Bondì, C.; Tinner, W. The past distribution of Abies nebrodensis (Lojac.) Mattei: Results of a multidisciplinary study. Veg. Hist. Archaeobot. 2020, 29, 357–371. [Google Scholar] [CrossRef]
- Tarraf, W.; Izgu, T.; Jouini, N. Strategies for the conservation by biotechnological approaches of Abies nebrodensis, a relict conifer of Sicily. Acta Hortic. 2023, 1359, 215–221. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef]
- Lewinsohn, E.; Gijzen, M.; Croteau, R. Mechanisms of conifers. Plant Physiol. 1991, 96, 44–49. [Google Scholar] [CrossRef]
- Lundborg, L.; Sampedro, L.; Borg-Karlson, A.K.; Zas, R. Effects of methyl jasmonate on the concentration of volatile terpenes in tissues of Maritime pine and Monterey pine and its relation to pine weevil feeding. Trees 2019, 33, 53–62. [Google Scholar] [CrossRef]
- Kim, T.; Song, B.; Cho, K.S.; Lee, I.S. Therapeutic potential of volatile terpenes and terpenoids from forests for inflammatory diseases. Int. J. Mol. Sci. 2020, 21, 2187. [Google Scholar] [CrossRef] [PubMed]
- Šimpraga, M.; Ghimire, R.P.; Van Der Straeten, D.; Blande, J.D.; Kasurinen, A.; Sorvari, J.; Holopainen, T.; Adriaenssens, S.; Holopainen, J.K.; Kivimäenpää, M. Unravelling the functions of biogenic volatiles in boreal and temperate forest ecosystems. Eur. J. For. Res. 2019, 138, 763–787. [Google Scholar] [CrossRef]
- Zulak, K.G.; Bohlmann, J. Terpenoid biosynthesis and specialized vascular cells of conifer defense. J. Integr. Plant Biol. 2010, 52, 86–97. [Google Scholar] [CrossRef]
- Kopaczyk, J.M.; Warguła, J.; Jelonek, T. The variability of terpenes in conifers under developmental and environmental stimuli. Environ. Exp. Bot. 2020, 180, 104197. [Google Scholar] [CrossRef]
- Zorić, M.; Kostić, S.; Kladar, N.; Božin, B.; Vasić, V.; Kebert, M.; Orlović, S. Phytochemical screening of volatile organic compounds in three common coniferous tree species in terms of forest ecosystem services. Forests 2021, 12, 928. [Google Scholar] [CrossRef]
- Raber, A.G.; Peachey-Stoner, R.J.; Cessna, S.G.; Siderhurst, M.S. Headspace GC-MS analysis of differences in intra- and interspecific Terpene profiles of Picea pungens Engelm. and P. abies (L.) Karst. Phytochemistry 2021, 181, 112541. [Google Scholar] [CrossRef]
- Hanover, J.W. Applications of terpene analysis in forest genetics. New For. 1992, 6, 159–178. [Google Scholar] [CrossRef]
- Plomion, C.; Yani, A.; Marpeau, A. Genetic determinism of δ3-carene in maritime pine using RAPD markers. Genome 1996, 39, 1123–1127. [Google Scholar] [CrossRef]
- Casano, S.; Grassi, G.; Martini, V.; Michelozzi, M. Variations in Terpene Profiles of Different Strains of Cannabis sativa L. CRA-CIN Consiglio per la Ricerca e la Sperimentazione in Agricoltura Centro di Ricerca per le Colture Industriali Rovigo Italy. Acta Hortic. 2011, 925, 115–122. [Google Scholar] [CrossRef]
- Nikolić, B.; Ristić, M.; Tešević, V.; Marin, P.D.; Bojović, S. Terpene chemodiversity of relict conifers picea omorika, pinus heldreichii, and pinus peuce, endemic to balkan. Chem. Biodivers. 2011, 8, 2247–2260. [Google Scholar] [CrossRef] [PubMed]
- Mitić, Z.S.; Jovanović, S.; Zlatković, B.K.; Nikolić, B.M.; Stojanović, G.S.; Marin, P.D. Needle terpenes as chemotaxonomic markers in Pinus: Subsections Pinus and Pinaster. Chem. Biodivers. 2017, 14, e1600453. [Google Scholar] [CrossRef] [PubMed]
- Mukrimin, M.; Kovalchuk, A.; Ghimire, R.P.; Kivimäenpää, M.; Sun, H.; Holopainen, J.K.; Asiegbu, F.O. Evaluation of potential genetic and chemical markers for Scots pine tolerance against Heterobasidion annosum infection. Planta 2019, 250, 1881–1895. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, J.S.; Zlatković, B.K.; Jovanović, S.; Stojanović, G.S.; Marin, P.D.; Mitić, Z.S. Needle volatiles as chemophenetic markers in differentiation of natural populations of Abies alba, A. x borisii-regis, and A. cephalonica. Phytochemistry 2021, 183, 112612. [Google Scholar] [CrossRef]
- Rajčević, N.; Nikolić, B.; Marin, P.D. Different responses to environmental factors in terpene composition of pinus heldreichii and P. peuce: Ecological and chemotaxonomic considerations. Arch. Biol. Sci. 2019, 71, 629–637. [Google Scholar] [CrossRef]
- Sadeghi, H.; Tahery, Y.; Moradi, S. Intra- and inter-specific variation of turpentine composition in Eldar pine (Pinus eldarica Medw.) and black pine (Pinus nigra Arnold). Biochem. Syst. Ecol. 2013, 48, 189–193. [Google Scholar] [CrossRef]
- Mitić, Z.S.; Jovanović, S.; Zlatković, B.K.; Milanovici, S.J.; Nikolić, B.M.; Petrović, G.M.; Stojanović, G.S.; Marin, P.D. Variation of needle volatiles in native populations of Pinus mugo–evidence from multivariate statistical analysis. Plant Biosyst. 2021, 155, 700–710. [Google Scholar] [CrossRef]
- Axelsson, K.; Zendegi-Shiraz, A.; Swedjemark, G.; Borg-Karlson, A.K.; Zhao, T. Chemical defence responses of Norway spruce to two fungal pathogens. For. Pathol. 2020, 50, e12640. [Google Scholar] [CrossRef]
- Kshatriya, K.; Whitehill, J.G.A.; Madilao, L.; Henderson, H.; Kermode, A.; Kolotelo, D.; Bohlmann, J. Histology of resin vesicles and oleoresin terpene composition of conifer seeds. Can. J. For. Res. 2018, 48, 1073–1084. [Google Scholar] [CrossRef]
- Calvo-Yuste, J.; Ruiz-Rodríguez, Á.L.; Hermosilla, B.; Agut, A.; Martínez-Ortega, M.M.; Tejero, P. Classification Importance of Seed Morphology and Insights on Large-Scale Climate-Driven Strophiole Size Changes in the Iberian Endemic Chasmophytic Genus Petrocoptis (Caryophyllaceae). Plants 2024, 13, 3208. [Google Scholar] [CrossRef]
- Singh, O.; Bordoloi, S.; Mahanta, N. Variability in cone, seed and seedling characteristics of Pinus kesiya Royle ex. Gordon. J. For. Res. 2015, 26, 331–337. [Google Scholar] [CrossRef]
- Hodžić, M.M.; Hajrudinović-Bogunić, A.; Bogunić, F.; Marku, V.; Ballian, D. Geographic variation of Pinus heldreichii Christ from the Western Balkans based on cone and seed morphology. Dendrobiology 2020, 84, 81–93. [Google Scholar] [CrossRef]
- Mustafa, E.; Tigabu, M.; Aldahadha, A.; Li, M. Variations in cone and seed phenotypic traits among and within populations of Aleppo pine in Jordan. New For. 2024, 55, 289–304. [Google Scholar] [CrossRef]
- Ghimire, B.; Yeom, D.; Jeong, M.J. Seed Atlas of Korea I. Conifers. J. Asia-Pac. Biodivers. 2019, 12, 459–466. [Google Scholar] [CrossRef]
- Shen, X.; Cho, M.J. Factors affecting seed germination and establishment of an efficient germination method in sugar pine (Pinus lambertiana Dougl.). HortScience 2021, 56, 299–304. [Google Scholar] [CrossRef]
- Skrzyszewska, K.; Chłanda, J. A study on the variation of morphological characteristics of silver fir (Abies alba Mill.) seeds and their internal structure determined by X-ray radiography in the beskid sâdecki and beskid niski mountain ranges of the Carpathians (southern Poland). J. For. Sci. 2009, 55, 403–414. [Google Scholar] [CrossRef]
- Harper, J.L.; Lovell, P.H.; Moore, K.G. The Shapes and Sizes of Seeds. Annu. Rev. Ecol. Syst. 1970, 1, 327–356. [Google Scholar] [CrossRef]
- Trujillo-Ríos, M.; Gazol, A.; Seco, J.I.; Linares, J.C. Phenotypic variation in cone scales and seeds as drivers of seedling germination dynamics of co-occurring cedar and fir species. Forests 2025, 16, 252. [Google Scholar] [CrossRef]
- Benelli, C.; Tarraf, W.; İzgü, T.; Anichini, M.; Faraloni, C.; Salvatici, M.C.; Antonietta, M.; Danti, R.; Lambardi, M. Long-term conservation for the safeguard of Abies nebrodensis: An endemic and endangered species of Sicily. Plants 2024, 13, 1682. [Google Scholar] [CrossRef]
- Arista, M.; Talavera, S. Density effect on the fruit-set, seed crop viability and seedling vigour of Abies pinsapo. Ann. Bot. 1996, 77, 187–192. [Google Scholar] [CrossRef]
- Hunt, R.S.; von Rudloff, E. Chemosystematic studies in the Genus Abies. IV. Introgression in Abies lasiocarpa and Abies bifolia. Taxon 1979, 28, 297–305. [Google Scholar] [CrossRef]
- Yang, X.W.; Li, S.M.; Shen, Y.H.; Zhang, W.D. Phytochemical and biological studies of Abies species. Chem. Biodivers. 2008, 5, 56–81. [Google Scholar] [CrossRef]
- Schicchi, R.; Geraci, A.; Rosselli, S.; Maggio, A.; Bruno, M. Chemodiversity of the essential oil from leaves of Abies nebrodensis (Lojac.) Mattei. Chem. Biodivers. 2017, 14, e1600254. [Google Scholar] [CrossRef] [PubMed]
- Rubino, F.M.; Martinoli, A.; Pitton, M.; Di Fabio, D.; Caruso, E.; Banfi, S.; Tosi, G.; Wauters, L.A.; Martinoli, A. Food choice of Eurasian red squirrels and concentrations of anti-predatory secondary compounds. Mamm. Biol. 2012, 77, 332–338. [Google Scholar] [CrossRef]
- Barrero, A.F.; Sanchez, J.F.; Alvarez-Manzaneda R., E.J.; Dorado, M.M. Sesquiterpenoids related to juvabione in Abies pinsapo. Phytochemistry 1989, 28, 2617–2619. [Google Scholar] [CrossRef]
- Barrero, A.F.; Sanchez, J.F.; Alvarez-Manzaneda, E.J.; Dorado, M.M.; Haidour, A. Terpenoids and sterols from the wood of Abies pinsapo. Phytochemistry 1993, 32, 1261–1265. [Google Scholar] [CrossRef]
- Nikolić, B.M.; Ballian, D.; Mitić, Z.S. Autochthonous conifers of family Pinaceae in Europe: Broad review of morpho-anatomical and phytochemical properties of needles and genetic investigations. Forests 2024, 15, 989. [Google Scholar] [CrossRef]
- Yang, S.A.; Jeon, S.K.; Lee, E.J.; Im, N.K.; Jhee, K.H.; Lee, S.P.; Lee, I.S. Radical scavenging activity of the essential oil of silver fir (Abies alba). J. Clin. Biochem. Nutr. 2009, 44, 253–259. [Google Scholar] [CrossRef]
- Bagci, E.; Babaç, M.T. A morphometric and chemosystematic study on the Abies Miller (Fir) species in Turkey. Acta Bot. Gall. 2003, 150, 355–367. [Google Scholar] [CrossRef]
- Warren, R.L.; Keeling, C.I.; Yuen, M.M.S.; Raymond, A.; Taylor, G.A.; Vandervalk, B.P.; Mohamadi, H.; Paulino, D.; Chiu, R.; Jackman, S.D.; et al. Improved white spruce (Picea glauca) genome assemblies and annotation of large gene families of conifer terpenoid and phenolic defense metabolism. Plant J. 2015, 83, 189–212. [Google Scholar] [CrossRef]
- Celedon, J.M.; Yuen, M.M.S.; Chiang, A.; Henderson, H.; Reid, K.E.; Bohlmann, J. Cell-type- and tissue-specific transcriptomes of the white spruce (Picea glauca) bark unmask fine-scale spatial patterns of constitutive and induced conifer defense. Plant J. 2017, 92, 710–726. [Google Scholar] [CrossRef]
- Fady, B.; Arbez, M.; Marpeau, A. Geographic variability of terpene composition in Abies cephalonica Loudon and Abies species around the Aegean: Hypotheses for their possible phylogeny from the Miocene. Trees 1992, 6, 162–171. [Google Scholar] [CrossRef]
- Fojtová, J.; Lojková, L.; Kubáň, V. Supercritical fluid extraction as a tool for isolation of monoterpenes from coniferous needles and walnut-tree leaves. Cent. Eur. J. Chem. 2010, 8, 409–418. [Google Scholar] [CrossRef]
- Mitić, Z.S.; Stojanović-Radić, Z.Z.; Jovanović, S.; Cvetković, V.J.; Nikolić, J.S.; Ickovski, J.D.; Mitrović, T.L.; Nikolić, B.M.; Zlatković, B.K.; Stojanović, G.S. Essential Oils of three Balkan Abies Species: Chemical profiles, antimicrobial activity and toxicity toward Artemia salina and Drosophila melanogaster. Chem. Biodivers. 2022, 19, e202200235. [Google Scholar] [CrossRef] [PubMed]
- Barbero, F.; Maffei, M. Biodiversity and chemotaxonomic significance of specialized metabolites. In Plant Specialized Metabolism; CRC Press: Boca Raton, FL, USA, 2016; pp. 35–76. [Google Scholar]
- Niederbacher, B.; Winkler, J.B.; Schnitzler, J.P. Volatile organic compounds as non-invasive markers for plant phenotyping. J. Exp. Bot. 2015, 66, 5403–5416. [Google Scholar] [CrossRef] [PubMed]
- Otto, A.; Volker, W. Sesqui-, di-, and triterpenoids as chemosystematic markers in extant conifers—A review. Bot. Rev. 2001, 67, 141–238. [Google Scholar] [CrossRef]
- Naydenov, K.D.; Tremblay, F.M.; Alexandrov, A.; Fenton, N.J. Structure of Pinus sylvestris L. populations in Bulgaria revealed by chloroplast microsatellites and terpenes analysis: Provenance tests. Biochem. Syst. Ecol. 2005, 33, 1226–1245. [Google Scholar] [CrossRef]
- von Rudloff, E. Volatile leaf oil analysis in chemosystematic studies of North American conifers. Biochem. Syst. Ecol. 1975, 2, 131–167. [Google Scholar] [CrossRef]
- Duquesnoy, E.; Castola, V.; Casanova, J. Composition and chemical variability of the twig oil of Abies alba Miller from Corsica Emilie. Flavour Fragr. J. 2008, 22, 293–299. [Google Scholar] [CrossRef]
- Mitsopoulos, D.J.; Panetsos, C.P. Origin of variation in Fir forests of Greece. Silvae Genet. 1987, 36, 1–7. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Salmerón-Gómez, R.; García-García, C.; García-Pérez, J. A Guide to Using the R package “multiColl” for detecting multicollinearity. Comput. Econ. 2021, 57, 529–536. [Google Scholar] [CrossRef]
- Salmeron, A.R.; Garcia, C.; Garcia, J.; Salmeron, M.R. Collinearity Detection in a Multiple Linear Regression Model. Available online: http://colldetreat.r-forge.r-project.org/ (accessed on 24 February 2025).
- Hastie, T.; Tibshirani, R.; Friedman, J.F. The Elements of Statistical Learning: Data Mining, Inference, and Prediction; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]
- Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. In Data Analysis; Wickham, H., Ed.; Springer: New York, NY, USA, 2016; pp. 189–201. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Brunson, J.C. Ordr: A Tdyverse Extension for Ordinations and Biplots. Available online: https://cran.r-project.org/web/packages/ordr/ordr.pdf (accessed on 24 February 2025).
- Karolis Koncevicius Package ‘matrixTests’. Available online: https://cran.r-project.org/web/packages/matrixTests/matrixTests.pdf (accessed on 24 February 2025).
- Pohler, T. Calculate Pairwise Multiple Comparisons of Mean Rank Sums. Available online: https://cran.r-project.org/web/packages/PMCMRplus/PMCMRplus.pdf (accessed on 24 February 2025).
- Labouriau, R. postHoc: Tools for Post-Hoc Analysis. R Package Version 0.1.3. Available online: https://tildeweb.au.dk/au33031/astatlab/software/posthoc/ (accessed on 24 February 2025).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).