


Nitrogen and Phosphorus Stoichiometry of Bolboschoenus planiculmis Plants in Soda–Alkali Wetlands Undergoing Agricultural Drainage Water Input in a Semi-Arid Region


Abstract
1. Introduction
2. Results
2.1. Variations in N and P Stoichiometry in Organs of B. planiculmis Plants
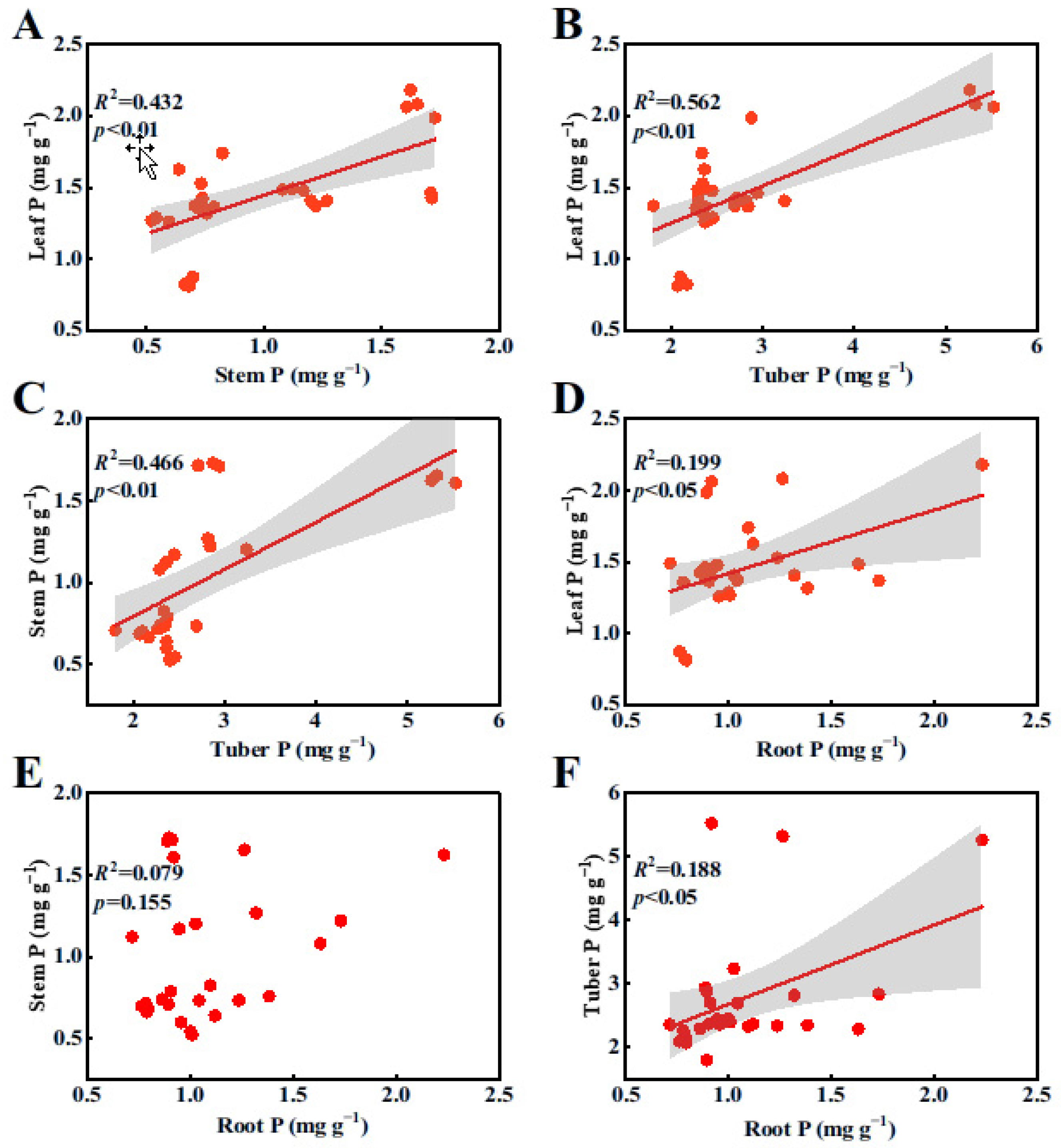
2.2. Relationships of N and P Contents in Organs of B. planiculmis Plants
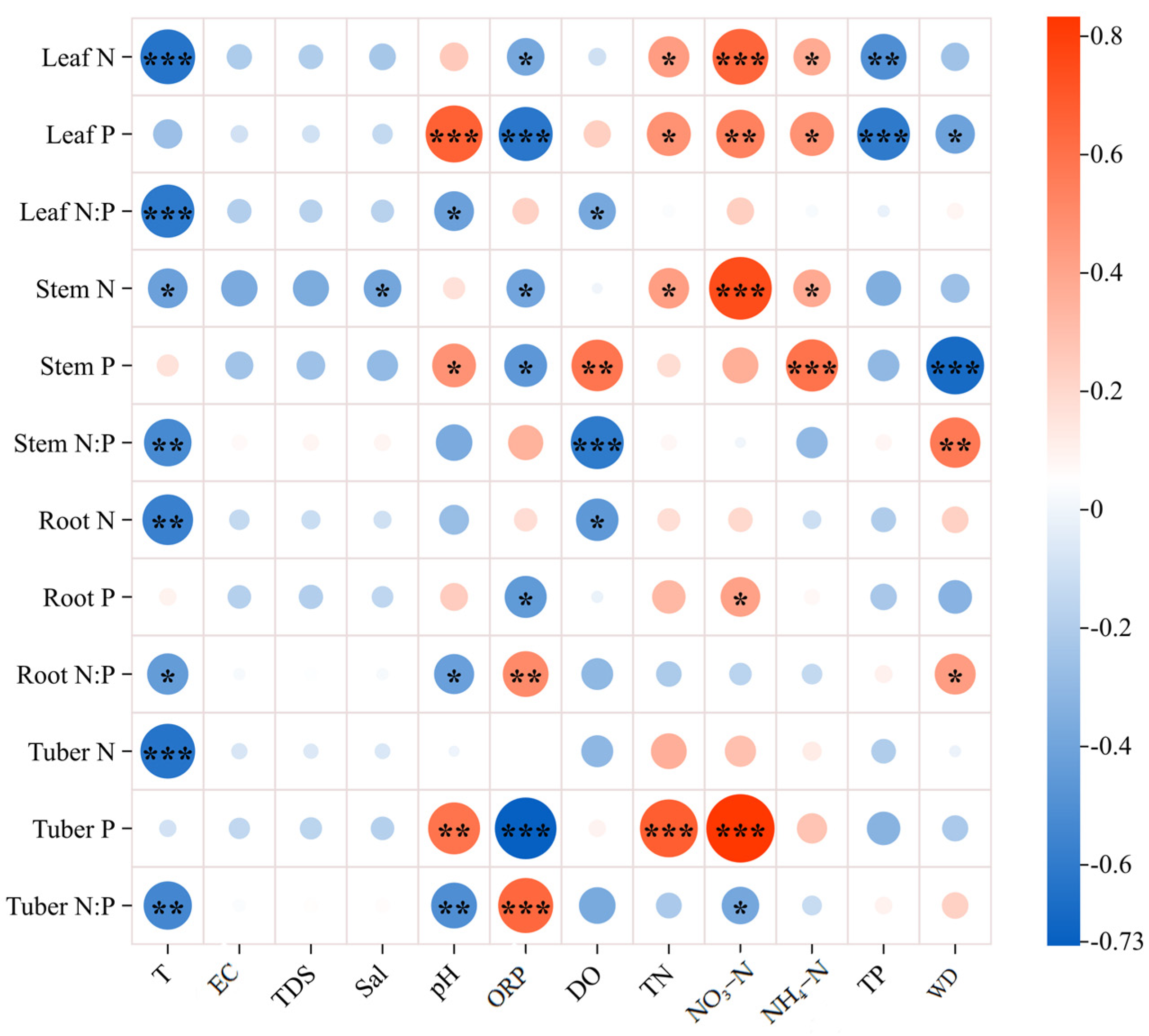
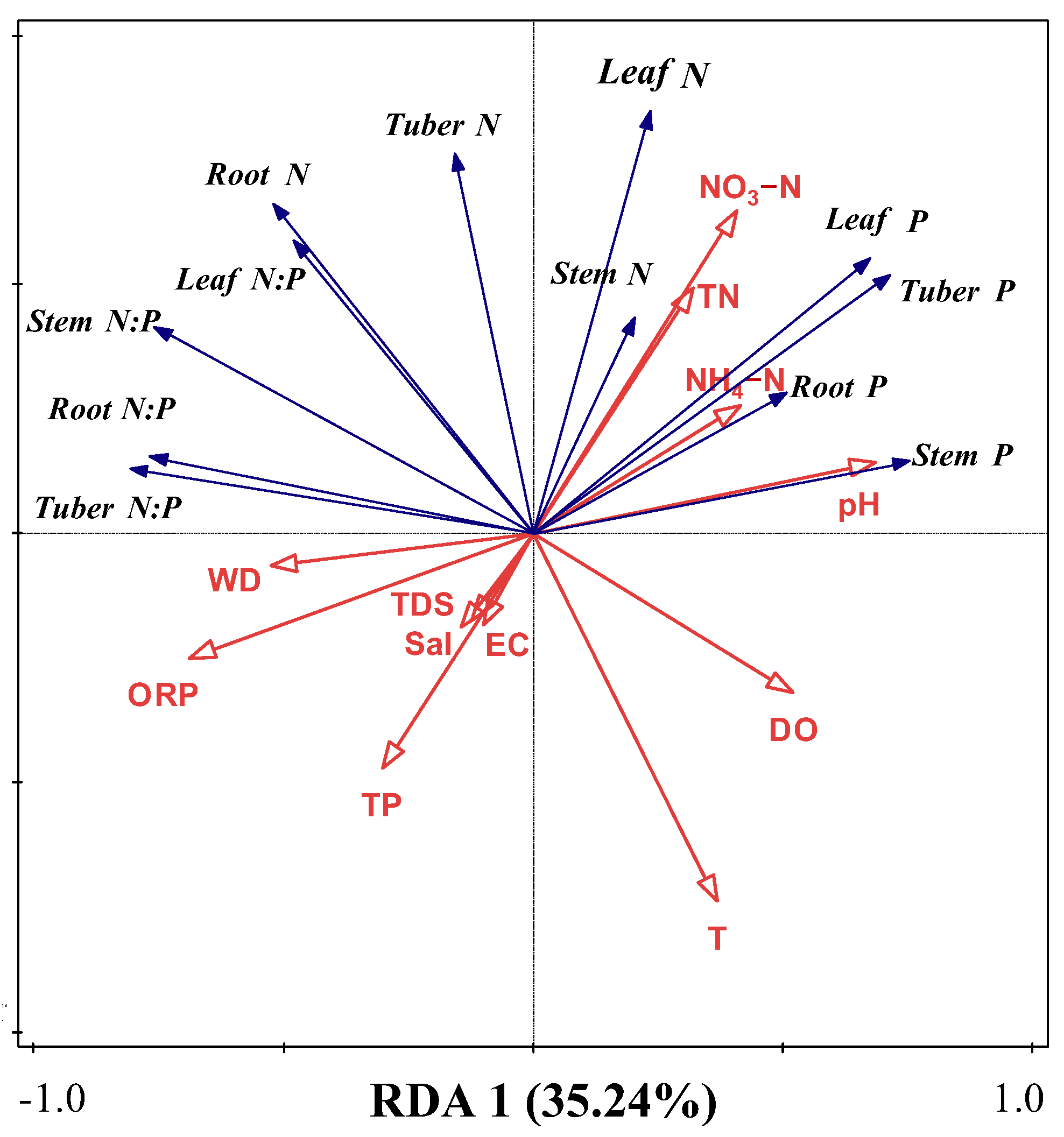
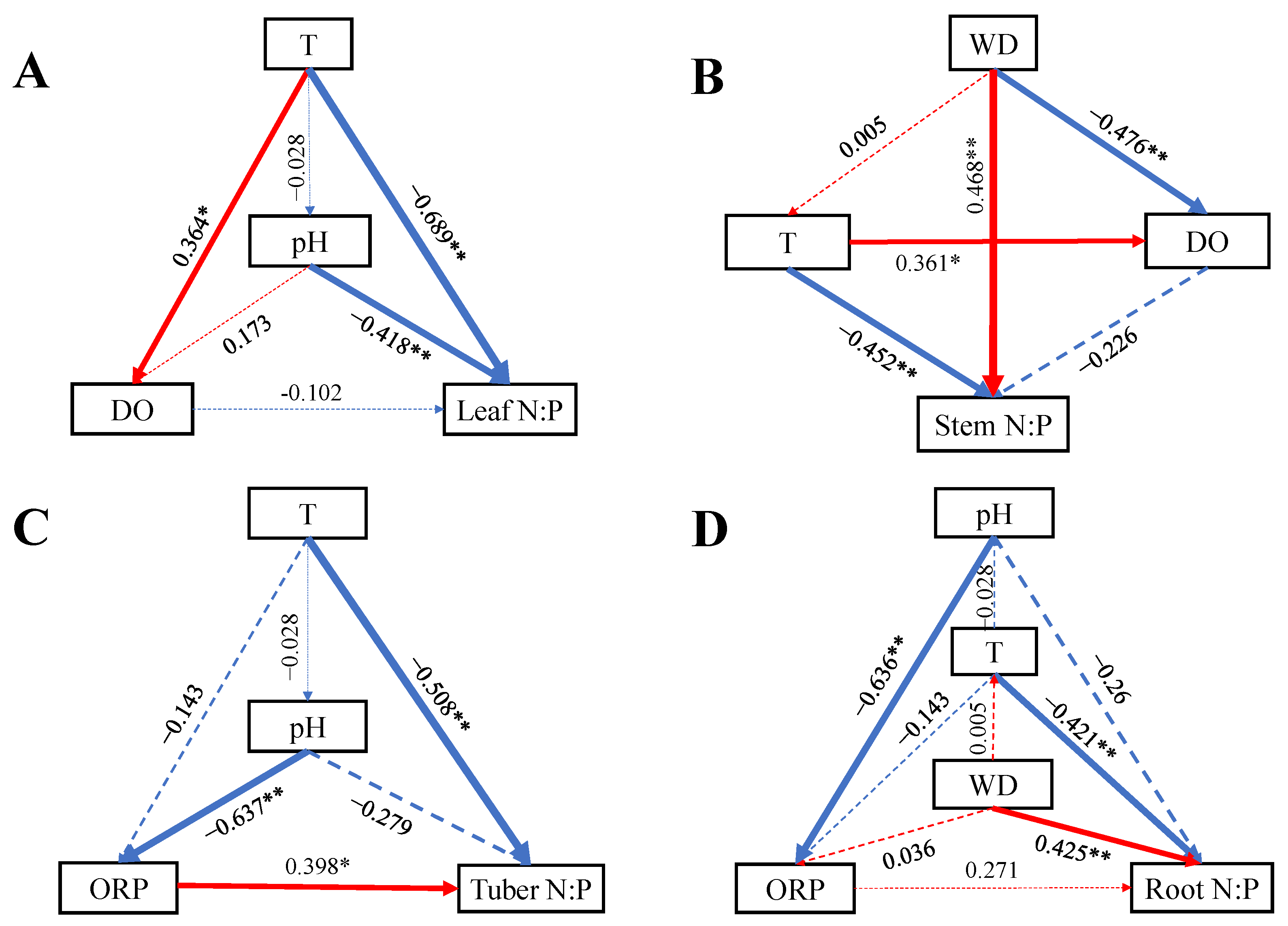
2.3. Effects of Water Variables on N, P, and N:P in Organs of B. planiculmis Plants
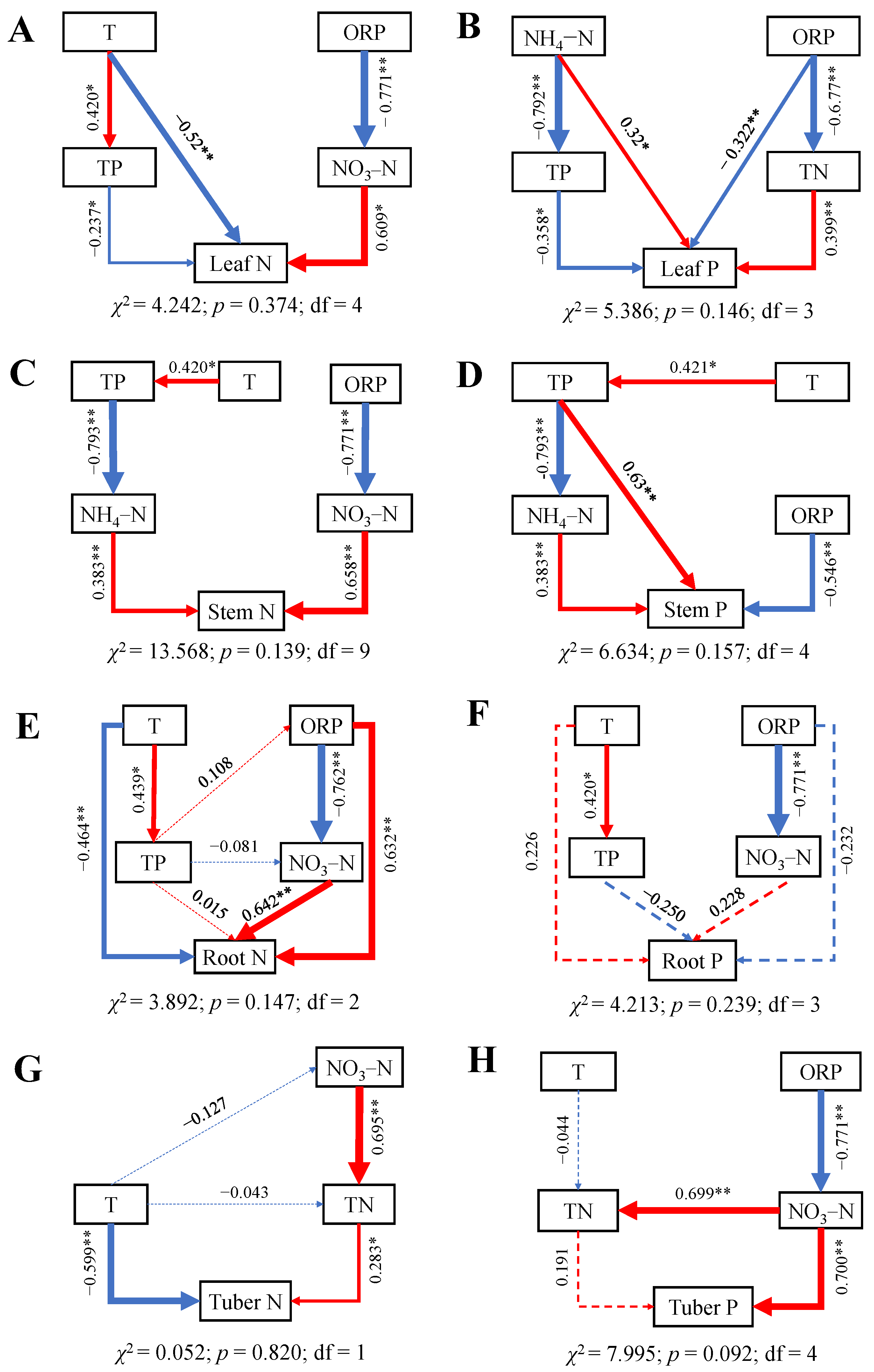
2.4. Influences of Water Variables on N and P Contents and N:P Ratio in Leaves, Stems, Tubers, and Roots
3. Discussion
4. Materials and Methods
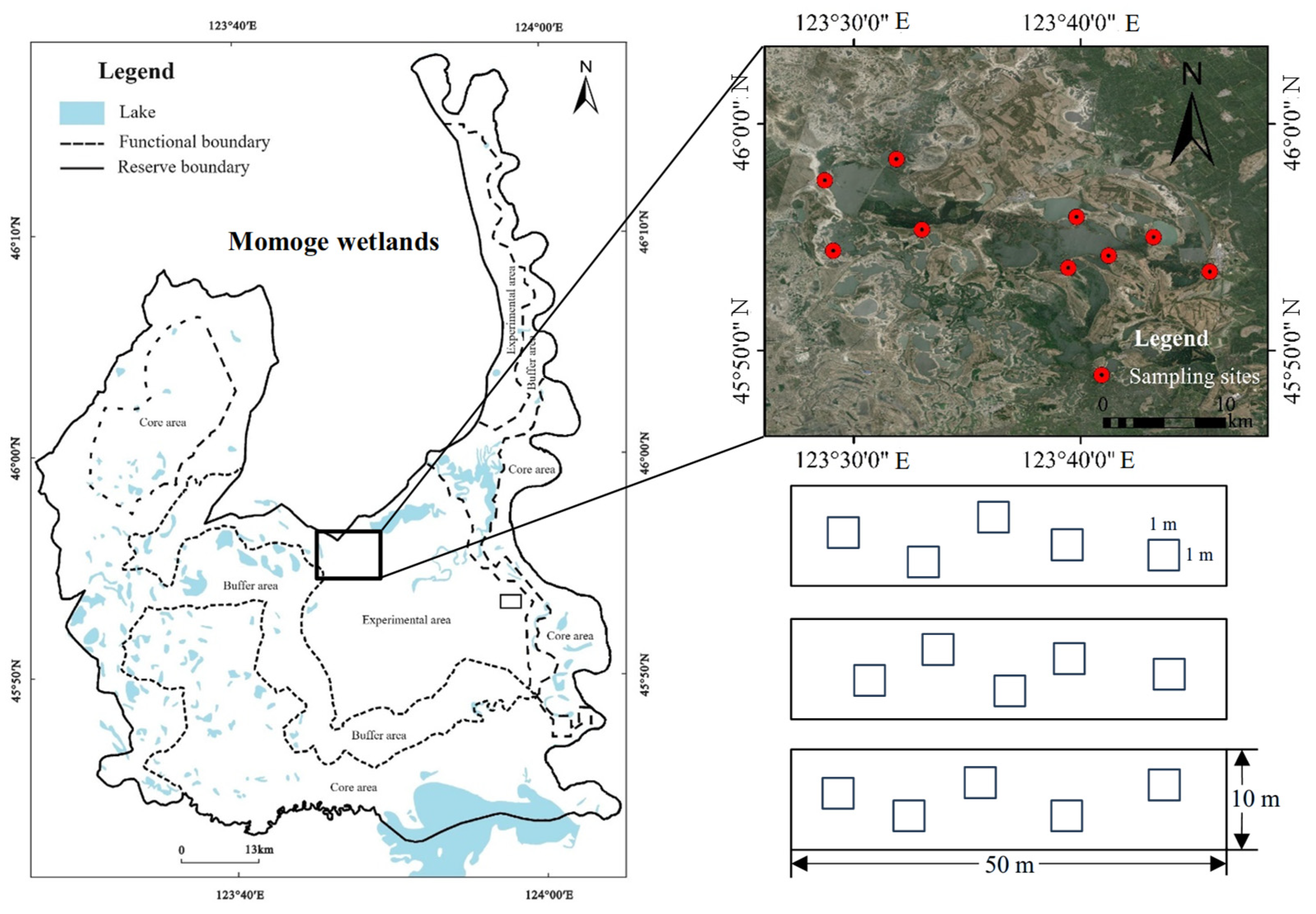
4.1. Study Area
4.2. Field Survey and Sampling
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, L.; Chen, D.; Huang, X.; Liu, Q.; Liang, J.; Hu, J.; Liu, Q. Variations of nitrogen and phosphorus between leaf, stem and root in shrubland biomes and responses to climate and soil factors across the Hengduan Mountains, China. Catena 2024, 241, 108008. [Google Scholar] [CrossRef]
- Mu, X.; Chen, Y. The physiological response of photosynthesis to nitrogen deficiency. Plant Physiol. Bioch. 2021, 158, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Xiao, W.; Mu, Q.; Li, D.; Chen, X.; Wu, H.; Li, L.; Peng, F. How does nitrate regulate plant senescence? Plant Physiol. Bioch. 2020, 157, 60–69. [Google Scholar] [CrossRef]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2013, 112, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Peng, Q.; Li, K.; Gong, Y.; Liu, Y.; Han, W. Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in Xinjiang, China. Catena 2021, 199, 105100. [Google Scholar] [CrossRef]
- Schreeg, L.A.; Santiago, L.S.; Wright, S.J.; Turner, B.L. Stem, root, and older leaf N:P ratios are more responsive indicators of soil nutrient availability than new foliage. Ecology 2014, 95, 2062–2068. [Google Scholar] [CrossRef] [PubMed]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Güsewell, S.; Freeman, C. Nutrient limitation and enzyme activities during litter decomposition of nine wetland species in relation to litter N:P ratios. Funct. Ecol. 2005, 19, 582–593. [Google Scholar] [CrossRef]
- Hong, M.G.; Kim, J.G. Effects of initial density, nutrient, and water level regime on the seedling survival and growth of Typha orientalis Presl. J. Plant Biol. 2016, 59, 369–376. [Google Scholar] [CrossRef]
- An, Y.; Song, T.; Zhang, Y.; Tong, S.; Liu, B. Optimum water depth for restoration of Bolboschoenus planiculmis in wetlands in semi-arid regions. Hydrobiologia 2022, 849, 13–28. [Google Scholar] [CrossRef]
- Magee, T.K.; Kentula, M.E. Response of wetland plant species to hydrologic conditions. Wetl. Ecol. Manag. 2005, 13, 163–181. [Google Scholar] [CrossRef]
- Li, B.; Yang, G.; Wan, R.; Lai, X.; Wagner, P.D. Impacts of hydrological alteration on ecosystem services changes of a large river-connected lake (Poyang Lake), China. J. Environ. Manag. 2022, 310, 114750. [Google Scholar] [CrossRef] [PubMed]
- Mitsch, W.J.; Gosselink, J.G. Wetlands; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar]
- O’geen, A.T.; Budd, R.; Gan, J.; Maynard, J.J.; Parikh, S.J.; Dahlgren, R.A. Mitigating nonpoint source pollution in agriculture with constructed and restored wetlands. Adv. Agron. 2010, 108, 1–76. [Google Scholar]
- Blann, K.L.; Anderson, J.L.; Sands, G.R.; Vondracek, B. Effects of agricultural drainage on aquatic ecosystems: A review. Crit. Rev. Env. Sic. Technol. 2009, 39, 909–1001. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, L.; Xue, Z.; E, M.; Jiang, M.; Lu, X.; Yang, S.; Shen, X.; Liu, Z.; Sun, G.; et al. Impacts of agricultural and reclamation practices on wetlands in the Amur River Basin, Northeastern China. Wetlands 2018, 38, 383–389. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiao, L.; Qin, H.; Li, F. Effect of environmental stress on the nutrient stoichiometry of the clonal plant Phragmites australis in inland riparian wetlands of Northwest China. Front. Plant Sci. 2021, 12, 705319. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Gao, Y.; Tong, S. Emergence and growth performance of Bolboschoenus planiculmis varied in response to water level and soil planting depth: Implications for wetland restoration using tuber transplantation. Aquat. Bot. 2018, 148, 10–14. [Google Scholar] [CrossRef]
- Zou, Y.; Zhang, S.; Yu, X.; Fu, G.; Lu, X. Isotopic evidence for soil water sources and reciprocal movement in a semi-arid degraded wetland: Implications for wetland restoration. Fund. Res. 2023, 3, 861–867. [Google Scholar] [CrossRef]
- An, Y.; Gao, Y.; Zhang, Y.; Tong, S.; Liu, X. Early establishment of Suaeda salsa population as affected by soil moisture and salinity: Implications for pioneer species introduction in saline-sodic wetlands in Songnen Plain, China. Ecol. Indic. 2019, 107, 105654. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, X.; Jiang, M.; Lu, X. Vegetation change and its response to climate change between 2000 and 2016 in marshes of the Songnen Plain, Northeast China. Sustainability 2020, 12, 3569. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, X.; Jiang, M.; Tong, S.; Lu, X. Daytime and nighttime temperatures exert different effects on vegetation net primary productivity of marshes in the western Songnen Plain. Ecol. Indic. 2022, 137, 108789. [Google Scholar] [CrossRef]
- Song, T.; An, Y.; Tong, S.; Zhang, W.; Wang, X.; Wang, L.; Jiang, L. Soil water conditions together with plant nitrogen acquisition strategies control vegetation dynamics in semi-arid wetlands undergoing land management changes. Catena 2023, 227, 107115. [Google Scholar] [CrossRef]
- Liu, B.; Jiang, M.; Tong, S.; Zhang, W.; Zou, C.; Wang, B.; Lu, X. Effects of burial depth and water depth on seedling emergence and early growth of Scirpus planiculmis Fr. Schmidt. Ecol. Eng. 2016, 87, 30–33. [Google Scholar] [CrossRef]
- An, Y.; Gao, Y.; Tong, S.; Liu, B. Morphological and physiological traits related to the response and adaption of Bolboschoenus planiculmis seedlings grown under salt-alkaline stress conditions. Front. Plant Sci. 2021, 12, 567782. [Google Scholar] [CrossRef] [PubMed]
- Moloney, T.; Fenton, O.; Daly, K. Ranking connectivity risk for phosphorus loss along agricultural drainage ditches. Sci. Total Environ. 2020, 703, 134556. [Google Scholar] [CrossRef] [PubMed]
- Singh, A. Salinization and drainage problems of agricultural land. Irrig. Drain. 2020, 69, 844–853. [Google Scholar] [CrossRef]
- Zhang, J.; He, N.; Liu, C.; Xu, L.; Chen, Z.; Li, Y.; Wang, R.; Yu, G.; Sun, W.; Xiao, C.; et al. Variation and evolution of C:N ratio among different organs enable plants to adapt to N-limited environments. Glob. Change Biol. 2020, 26, 2534–2543. [Google Scholar] [CrossRef]
- Zhong, C.; Jian, S.; Huang, J.; Jin, Q.; Cao, X. Trade-off of within-leaf nitrogen allocation between photosynthetic nitrogen-use efficiency and water deficit stress acclimation in rice (Oryza sativa L.). Plant Physiol. Bioch. 2019, 135, 41–50. [Google Scholar] [CrossRef]
- Li, K.; Ren, H.; Zhao, W.; Zhao, X.; Gan, C. Factors affecting bulblet multiplication in bulbous plants. Sci. Hortic. 2023, 312, 111837. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y. Plant mixture balances terrestrial ecosystem C:N:P stoichiometry. Nat. Commun. 2021, 12, 4562. [Google Scholar] [CrossRef] [PubMed]
- Pezeshki, S.R. Wetland plant responses to soil flooding. Environ. Exp. Bot. 2001, 46, 299–312. [Google Scholar] [CrossRef]
- Yamauchi, T.; Colmer, T.D.; Pedersen, O.; Nakazono, M. Regulation of root traits for internal aeration and tolerance to soil waterlogging-flooding stress. Plant Physiol. 2018, 176, 1118–1130. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Env. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef]
- Tyagi, J.; Ahmad, S.; Malik, M. Nitrogenous fertilizers: Impact on environment sustainability, mitigation strategies, and challenges. Int. J. Environ. Sci. Technol. 2022, 19, 11649–11672. [Google Scholar] [CrossRef]
- Yang, Z.; Midmore, D.J. Modelling plant resource allocation and growth partitioning in response to environmental heterogeneity. Ecol. Model. 2005, 181, 59–77. [Google Scholar] [CrossRef]
- Wang, H.; Inukai, Y.; Yamauchi, A. Root development and nutrient uptake. Crit. Rev. Plant Sci. 2006, 25, 279–301. [Google Scholar] [CrossRef]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef]
- Xing, W.; Han, Y.; Guo, Z.; Zhou, Y. Quantitative study on redistribution of nitrogen and phosphorus by wetland plants under different water quality conditions. Environ. Pollut. 2020, 261, 114086. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Shabala, L.; Barcelo, J.; Poschenrieder, C. Membrane transporters mediating root signalling and adaptive responses to oxygen deprivation and soil flooding. Plant Cell Environ. 2014, 37, 2216–2233. [Google Scholar] [CrossRef] [PubMed]
- Daniel, K.; Hartman, S. How plant roots respond to waterlogging. J. Exp. Bot. 2024, 75, 511–525. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, Z.; Shen, X.; Liu, D.; Pedersen, O. Flooding-adaptive root and shoot traits in rice. Funct. Plant Biol. 2024, 51, FP23226. [Google Scholar] [CrossRef]
- Zeng, F.; Ali, S.; Zhang, H.; Ouyang, Y.; Qiu, B.; Wu, F.; Zhang, G. The influence of pH and organic matter content in paddy soil on heavy metal availability and their uptake by rice plants. Environ. Pollut. 2011, 159, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Jiang, D.; Chen, Y.; Wei, J.; Xia, F.; Xie, W.; Zhou, Y.; Li, X.; Deng, S. Spatial-temporal distribution, morphological transformation, and potential risk of dissolved inorganic nitrogen in the contaminated unconfined aquifer from a retired nitrogenous fertilizer plant. J. Environ. Res. Public Health 2022, 19, 8022. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wang, Y.; Dong, Y. A review of methods, influencing factors and mechanisms for phosphorus recovery from sewage and sludge from municipal wastewater treatment plants. J. Environ. Chem. Eng. 2023, 12, 111657. [Google Scholar] [CrossRef]
- Ou, Y.; Rousseau, A.N.; Wang, L.; Yan, B.; Gumiere, T.; Zhu, H. Identification of the alteration of riparian wetland on soil properties, enzyme activities and microbial communities following extreme flooding. Geoderma 2019, 337, 825–833. [Google Scholar] [CrossRef]
- Kaur, G.; Singh, G.; Motavalli, P.P.; Nelson, K.A.; Orlowski, J.M.; Golden, B.R. Impacts and management strategies for crop production in waterlogged or flooded soils: A review. Agron. J. 2020, 112, 1475–1501. [Google Scholar] [CrossRef]
- Wang, Y.; Lambers, H. Root-released organic anions in response to low phosphorus availability: Recent progress, challenges and future perspectives. Plant Soil 2020, 447, 135–156. [Google Scholar] [CrossRef]
- Lai, W.L.; Wang, S.Q.; Peng, C.L.; Chen, Z.H. Root features related to plant growth and nutrient removal of 35 wetland plants. Water Res. 2011, 45, 3941–3950. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, C.; Li, H.; Liu, J.; Jiang, T.; Yan, D.; Tong, J.; Dong, L. Review on ecological response of aquatic plants to balanced harvesting. Sustainability 2022, 14, 12451. [Google Scholar] [CrossRef]
- Reich, P.B.; Hobbie, S.E.; Lee, T.D. Plant growth enhancement by elevated CO2 eliminated by joint water and nitrogen limitation. Nat. Geosci. 2014, 7, 920–924. [Google Scholar] [CrossRef]
- Touchette, B.W.; Burkholder, J.M. Review of nitrogen and phosphorus metabolism in seagrasses. J. Exp. Mar. Biol. Ecol. 2000, 250, 133–167. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Wu, H.H.; Smith, M.D.; La Pierre, K.J.; Lü, X.T.; Zhang, H.Y.; Han, X.G.; Yu, Q. Nitrogen deposition promotes phosphorus uptake of plants in a semi-arid temperate grassland. Plant Soil 2016, 408, 475–484. [Google Scholar] [CrossRef]
- Guan, P. Dancing with hormones: A current perspective of nitrate signaling and regulation in Arabidopsis. Front. Plant Sci. 2017, 8, 1697. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf | Stem | Tuber | Root | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Nutrient | Mean ± SE | CV (%) | Mean ± SE | CV (%) | Mean ± SE | CV (%) | Mean ± SE | CV (%) | F | p |
| N (g kg−1) | 16.22 ± 4.62 a | 28.47 | 6.21 ± 1.74 b | 27.95 | 7.55 ± 1.71 b | 22.70 | 7.14 ± 1.13 b | 15.80 | 75.976 | <0.001 |
| P (g kg−1) | 1.45 ± 0.32 b | 22.04 | 1.02 ± 0.41 c | 40.07 | 2.78 ± 0.95 a | 34.26 | 1.08 ± 0.21 c | 19.51 | 53.155 | <0.001 |
| N:P | 11.2 ± 2.16 a | 19.28 | 6.71 ± 2.02 b | 30.04 | 2.89 ± 0.90 c | 31.26 | 7.07 ± 1.94 b | 27.48 | 90.322 | <0.001 |
| Variables | Simple Effects (Explains%) | F | p | Conditional Effects (Explains%) | F | p |
|---|---|---|---|---|---|---|
| T | 19.9 | 6.2 | 0.002 | 19.9 | 6.2 | 0.002 |
| ORP | 19.1 | 5.9 | 0.002 | 18.6 | 7.3 | 0.002 |
| NH4–N | 9.6 | 2.7 | 0.028 | 11.8 | 5.5 | 0.002 |
| NO3–N | 18.1 | 5.5 | 0.004 | 8.7 | 4.7 | 0.002 |
| WD | 11.6 | 3.3 | 0.010 | 5.0 | 2.9 | 0.022 |
| TN | 10.9 | 3.1 | 0.022 | 3.5 | 2.3 | 0.036 |
| pH | 17.8 | 5.4 | 0.004 | 0.9 | 0.7 | 0.592 |
| DO | 13.3 | 3.8 | 0.014 | 2.8 | 1.9 | 0.116 |
| TP | 9.7 | 2.7 | 0.032 | 3.3 | 2.0 | 0.066 |
| Sal | 2.1 | 0.5 | 0.714 | 1.0 | 0.7 | 0.594 |
| TDS | 1.8 | 0.5 | 0.804 | 3.0 | 2.2 | 0.076 |
| EC | 1.7 | 0.4 | 0.808 | 2.8 | 2.2 | 0.084 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, Y.; Wang, L.; Liu, B.; Wu, H.; Tong, S. Nitrogen and Phosphorus Stoichiometry of Bolboschoenus planiculmis Plants in Soda–Alkali Wetlands Undergoing Agricultural Drainage Water Input in a Semi-Arid Region. Plants 2025, 14, 787. https://doi.org/10.3390/plants14050787
An Y, Wang L, Liu B, Wu H, Tong S. Nitrogen and Phosphorus Stoichiometry of Bolboschoenus planiculmis Plants in Soda–Alkali Wetlands Undergoing Agricultural Drainage Water Input in a Semi-Arid Region. Plants. 2025; 14(5):787. https://doi.org/10.3390/plants14050787
Chicago/Turabian StyleAn, Yu, Le Wang, Bo Liu, Haitao Wu, and Shouzheng Tong. 2025. "Nitrogen and Phosphorus Stoichiometry of Bolboschoenus planiculmis Plants in Soda–Alkali Wetlands Undergoing Agricultural Drainage Water Input in a Semi-Arid Region" Plants 14, no. 5: 787. https://doi.org/10.3390/plants14050787
APA StyleAn, Y., Wang, L., Liu, B., Wu, H., & Tong, S. (2025). Nitrogen and Phosphorus Stoichiometry of Bolboschoenus planiculmis Plants in Soda–Alkali Wetlands Undergoing Agricultural Drainage Water Input in a Semi-Arid Region. Plants, 14(5), 787. https://doi.org/10.3390/plants14050787