
Neighbor Relatedness Contributes to Improvement in Grain Yields in Rice Cultivar Mixtures



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
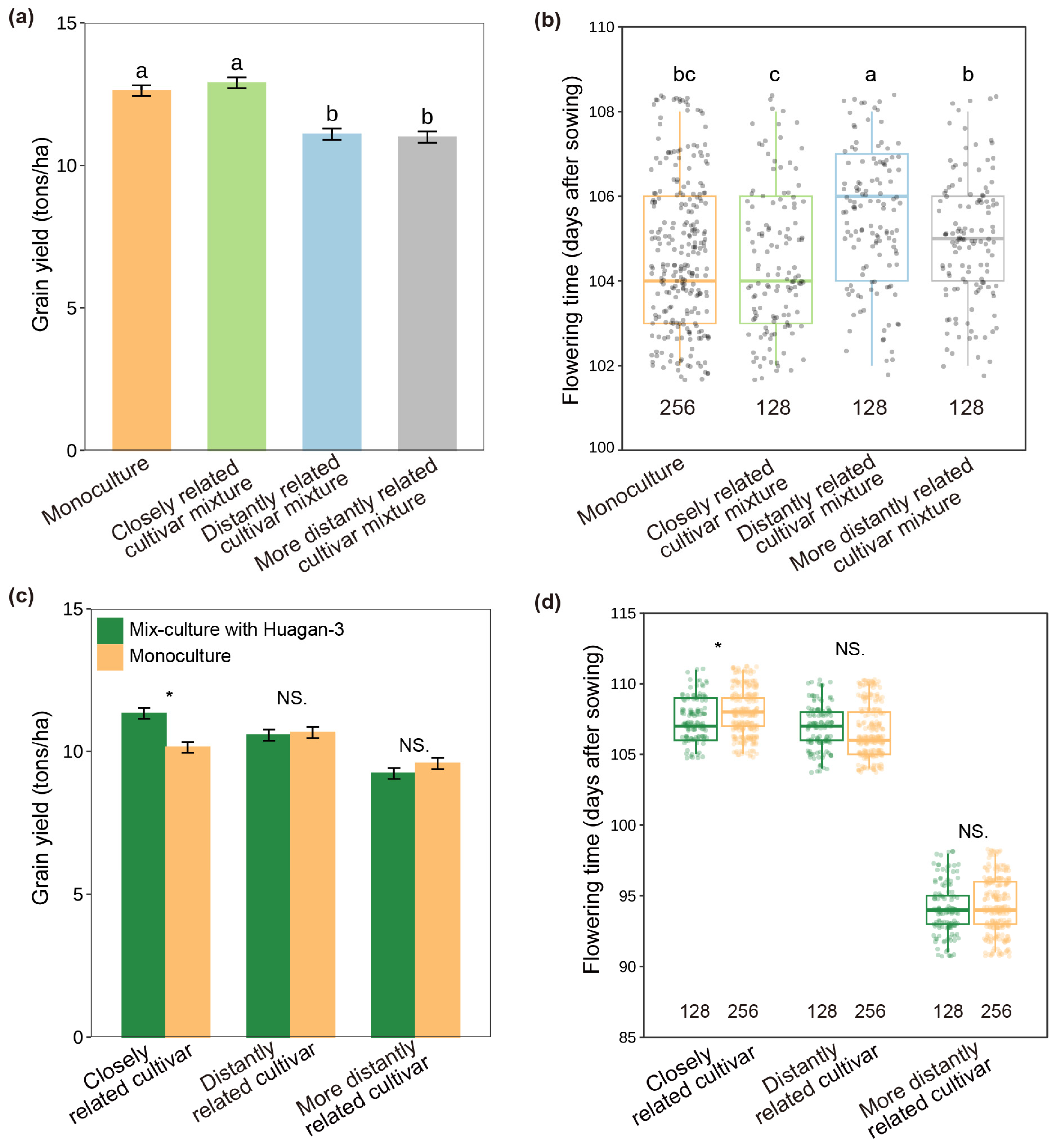
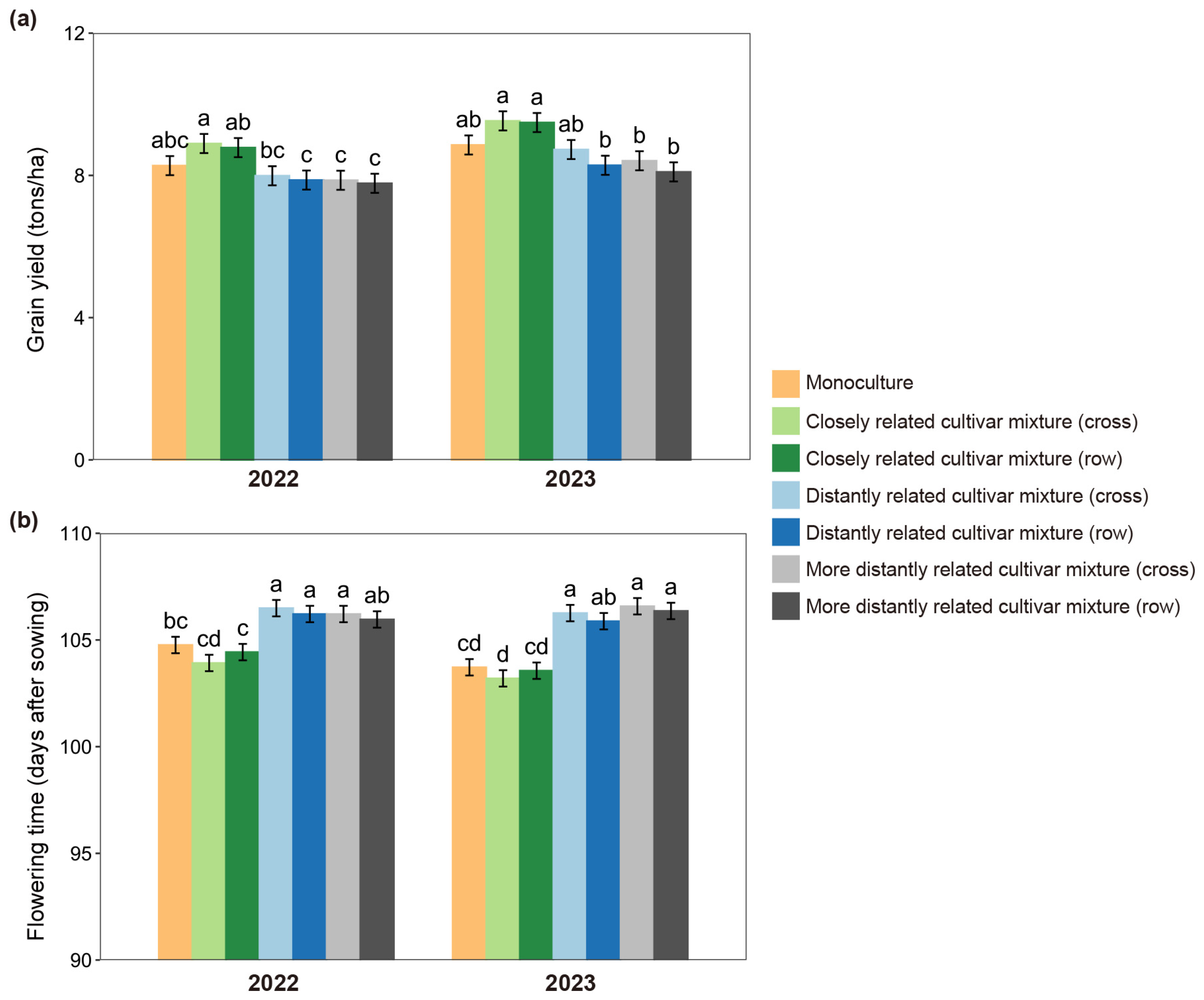
2.1. Effect of Neighbor Relatedness on Grain Yields
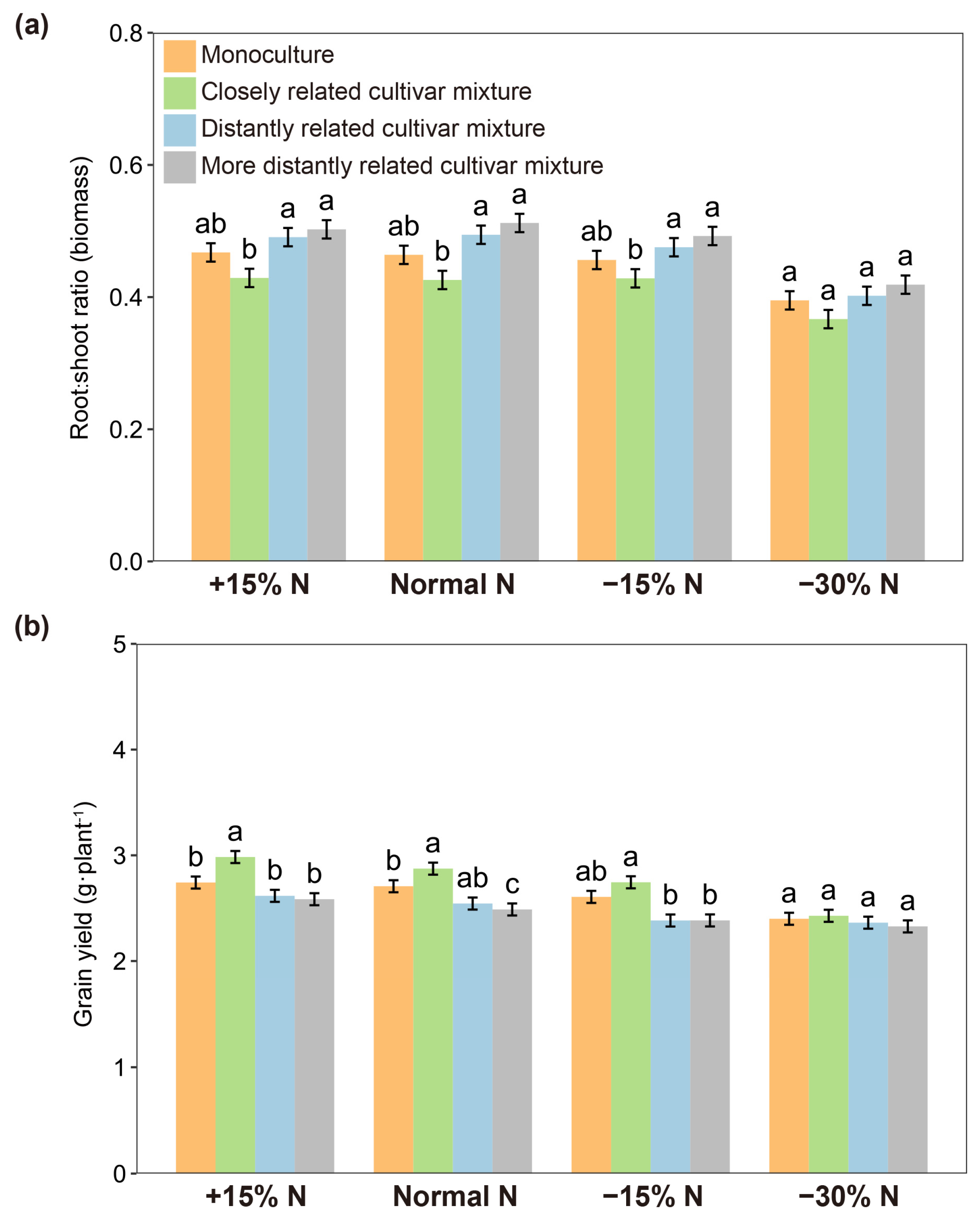
2.2. Soil Volume and Nitrogen Use Level Influence the Effect of Relatedness
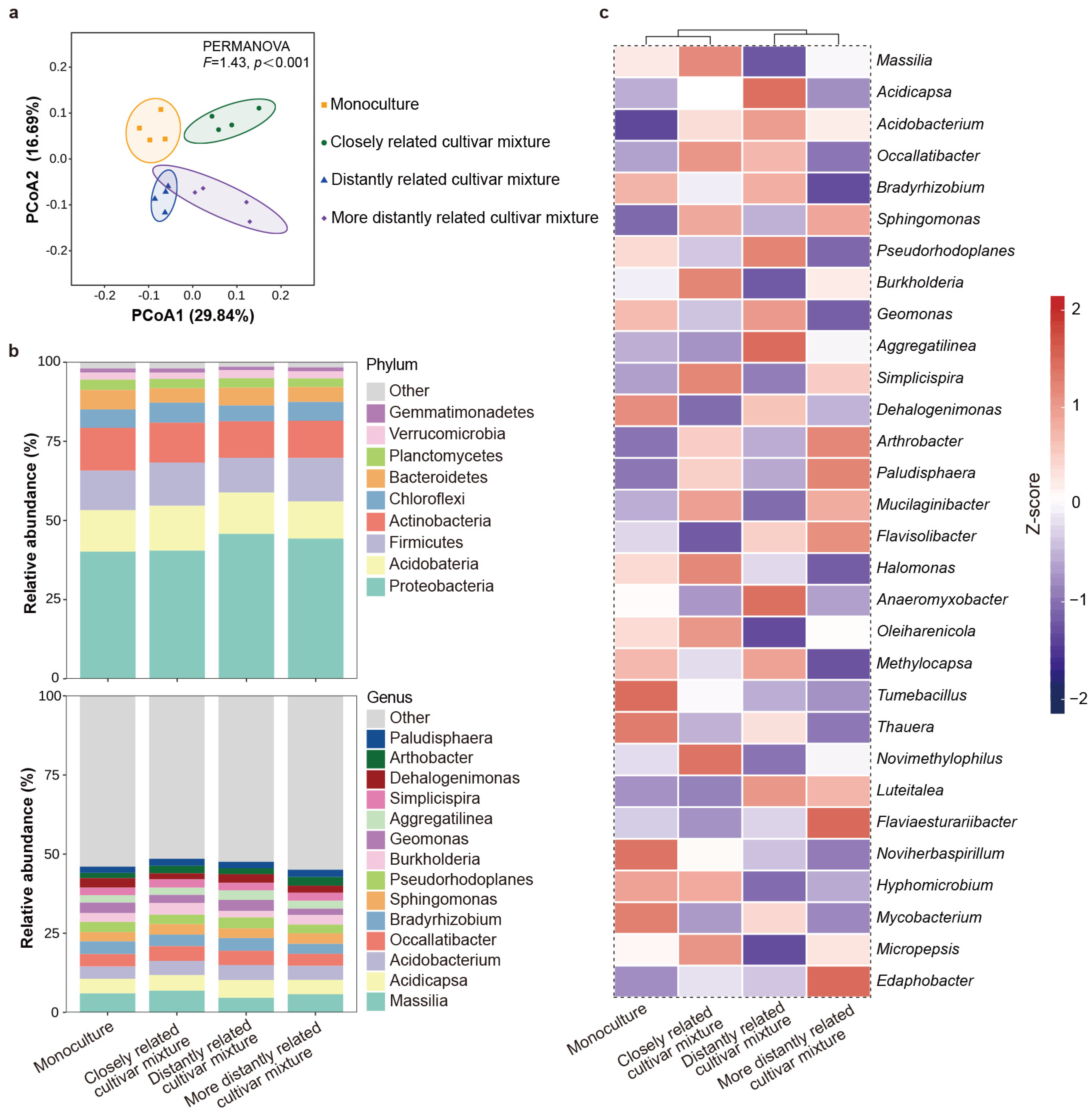
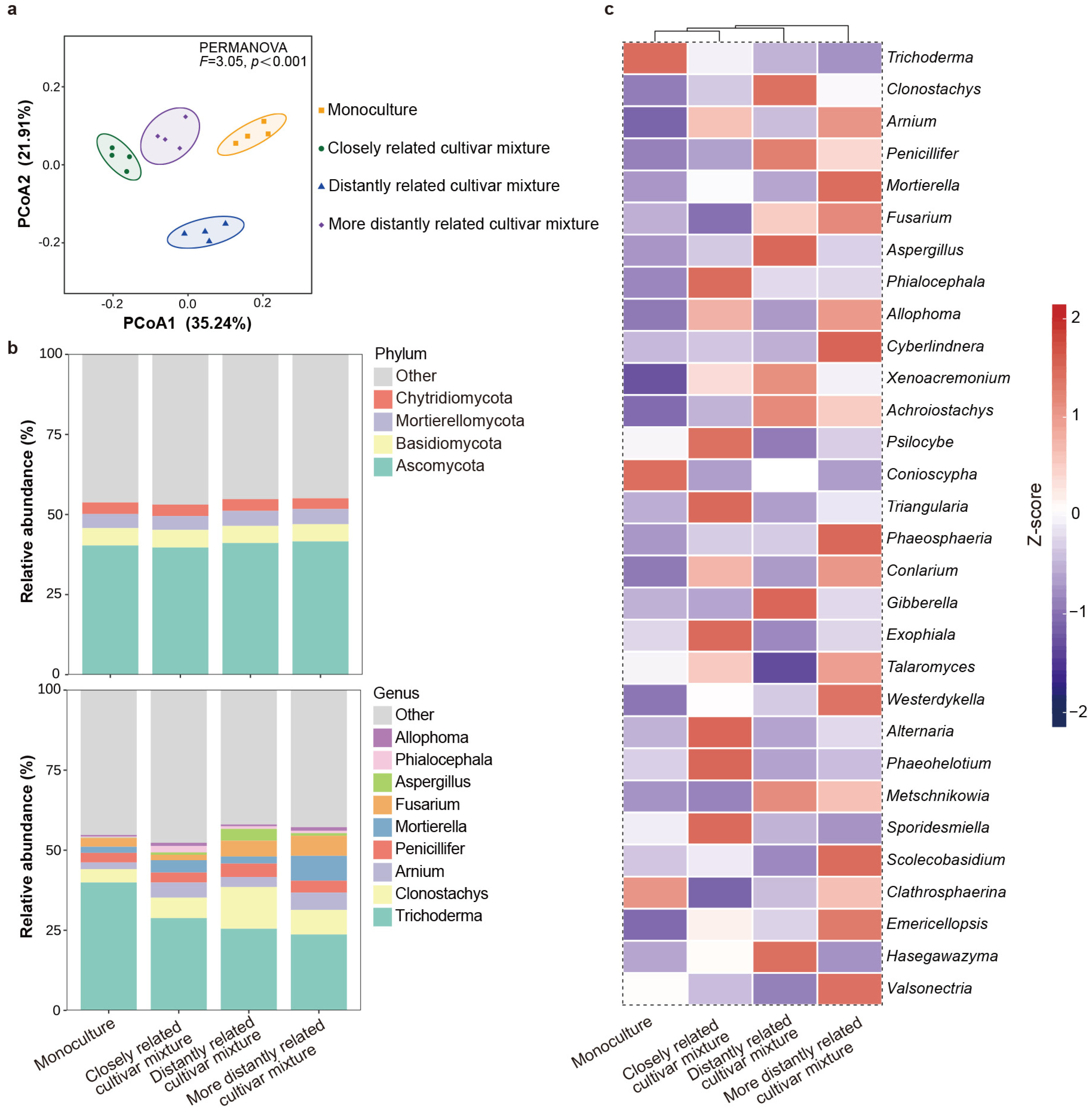
2.3. Effect of Neighbor Relatedness on Rhizosphere Soil Microbes
3. Discussion
4. Materials and Methods
4.1. Rice Cultivars and Their Relatedness
4.2. Field Trials
4.3. Greenhouse Experiments
4.4. Rhizosphere Soil Sampling
4.5. DNA Extraction, PCR Amplification, and Sequencing
4.6. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grettenberger, I.M.; Tooker, J.F. Inter-varietal interactions among plants in genotypically diverse mixtures tend to decrease herbivore performance. Oecologia 2016, 182, 189–202. [Google Scholar] [CrossRef]
- Barot, S.; Allard, V.; Cantarel, A.; Enjalbert, J.; Gauffreteau, A.; Goldringer, I.; Lata, J.C.; Le Roux, X.; Niboyet, A.; Porcher, E. Designing mixtures of varieties for multifunctional agriculture with the help of ecology. A review. Agron. Sustain. Dev. 2017, 37, 13. [Google Scholar] [CrossRef]
- Pelissier, R.; Ballini, E.; Temple, C.; Ducasse, A.; Colombo, M.; Frouin, J.; Qin, X.P.; Huang, H.C.; Jacques, D.; Florian, F. The genetic identity of neighboring plants in intraspecific mixtures modulates disease susceptibility of both wheat and rice. PLoS Biol. 2023, 21, e3002287. [Google Scholar]
- Reiss, E.R.; Drinkwater, L.E. Cultivar mixtures: A meta-analysis of the effect of intraspecific diversity on crop yield. Ecol. Appl. 2018, 28, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Pan, Z.; Zhu, W.; Wu, E.; He, D.; Yuan, X.; Qin, Y.; Wang, Y.; Chen, R.; Thrall, P.H.; et al. Enhanced agricultural sustainability through within-species diversification. Nat. Sustain. 2019, 2, 46–52. [Google Scholar] [CrossRef]
- Wuest, S.E.; Peter, R.; Niklaus, P.A. Ecological and evolutionary approaches to improving crop variety mixtures. Nat. Ecol. Evol. 2021, 5, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Schofield, E.J.; Rowntree, J.K.; Paterson, E.; Brewer, M.J.; Price, E.A.C.; Brearley, F.Q.; Brooker, R.W. Cultivar differences and impact of plant–plant competition on temporal patterns of nitrogen and biomass accumulation. Front. Plant Sci. 2019, 10, 215. [Google Scholar] [CrossRef]
- Dahlin, I.; Kiær, L.P.; Bergkvist, G.; Weih, M.; Ninkovic, V. Plasticity of barley in response to plant neighbors in cultivar mixtures. Plant Soil 2020, 447, 537–551. [Google Scholar] [CrossRef]
- Cossani, C.M.; Sadras, V.O. Symmetric response to competition in binary mixtures of cultivars associates with genetic gain in wheat yield. Evol. Appl. 2021, 14, 2064–2078. [Google Scholar] [CrossRef]
- Fréville, H.; Montazeaud, G.; Forst, E.; David, J.; Papa, R.; Tenaillon, M.I. Shift in beneficial interactions during crop evolution. Evol. Appl. 2020, 15, 905–918. [Google Scholar] [CrossRef]
- Dudley, S.A.; File, A.L. Kin recognition in an annual plant. Biol. Lett. 2007, 3, 435–438. [Google Scholar] [CrossRef]
- Torices, R.; Gómez, J.M.; Pannell, J.R. Kin discrimination allows plants to modify investment towards pollinator attraction. Nat. Commun. 2018, 9, 2018. [Google Scholar] [CrossRef]
- Li, J.; Zhao, Q.X.; Li, W.L.; He, J.Z.; Xu, X.L. Distinct kin strategies of the legume soybean and the non-legume balsam by accomplishing different nitrogen acquisition and rhizosphere microbiome composition. Plant J. 2022, 110, 103–113. [Google Scholar] [CrossRef]
- Lewis, I.; Friedman, J. Kin discrimination causes plastic responses in floral and clonal allocation. Proc. R. Soc. B 2025, 292, 20242387. [Google Scholar] [CrossRef]
- Tomizuka, H.; Yamawo, A.; Tachiki, Y. Altruism or selfishness: Floral behavior based on genetic relatedness with neighboring plants. J. Evolution. Biol. 2025, 38, 492–503. [Google Scholar] [CrossRef]
- Dudley, S.A.; Murphy, G.P.; File, A.L. Kin recognition and competition in plants. Funct. Ecol. 2013, 27, 898–906. [Google Scholar] [CrossRef]
- Ehlers, B.K.; David, P.; Damgaard, C.F.; Lenormand, T. Competitor relatedness, indirect soil effects and plant coexistence. J. Ecol. 2016, 104, 1126–1135. [Google Scholar] [CrossRef]
- Xu, Y.; Cheng, H.F.; Kong, C.H.; Meiners, S.J. Intra-specific kin recognition contributes to inter-specific allelopathy: A case study of allelopathic rice interference with paddy weeds. Plant Cell Environ. 2021, 44, 3479–3491. [Google Scholar] [CrossRef] [PubMed]
- Mazal, L.; Fajardo, A.; Till-Bottraud, I.; Corenblit, D.; Fumanal, B. Kin selection, kin recognition and kin discrimination in plants revisited: A claim for considering environmental and genetic variability. Plant Cell Environ. 2023, 46, 2007–2016. [Google Scholar] [CrossRef]
- Xia, X.X.; Yan, S.B.; Wang, P.; Kong, C.H. A meta-analysis of response strategies and interfering factors of kin recognition in plants. Plants 2025, 14, 683. [Google Scholar] [CrossRef]
- Lepik, A.; Abakumova, M.; Zobel, K.; Semchenko, M. Kin recognition is density-dependent and uncommon among temperate grassland plants. Funct. Ecol. 2012, 26, 1214–1220. [Google Scholar] [CrossRef]
- Ding, L.; Zhao, H.H.; Li, H.Y.; Yang, X.F.; Kong, C.H. Kin recognition in an herbicide-resistant barnyardgrass (Echinochloa crus-galli L.) biotype. Plants 2023, 12, 1498. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.F.; Li, L.L.; Xu, Y.; Kong, C.H. Kin recognition in rice (Oryza sativa) lines. New Phytol. 2018, 220, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Pezzola, E.; Pandolfi, C.; Mancuso, S. Resource availability affects kin selection in two cultivars of Pisum sativum. Plant Growth Regul. 2020, 90, 321–329. [Google Scholar] [CrossRef]
- Kiers, E.T.; Denison, R.F. Inclusive fitness in agriculture. Phil. Trans. R. Soc. B 2014, 369, 20130367. [Google Scholar] [CrossRef]
- Murphy, G.P.; Swanton, C.J.; van Acker, R.C.; Dudley, S.A. Kin recognition, multilevel selection and altruism in crop sustainability. J. Ecol. 2017, 105, 930–934. [Google Scholar] [CrossRef]
- Fang, S.; Clark, R.T.; Zheng, Y.; Iyer-Pascuzzi, A.S.; Weitz, J.S.; Kochian, L.V. Genotypic recognition and spatial responses by rice roots. Proc. Natl. Acad. Sci. USA 2013, 110, 2670–2675. [Google Scholar] [CrossRef]
- Wu, C.C.; Diggle, P.K.; Friedman, W.E. Kin recognition within a seed and the effect of genetic relatedness of an endosperm to its compatriot embryo on maize seed development. Proc. Natl. Acad. Sci. USA 2013, 110, 2217–2222. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Q.Y.; Tian, Y.Q.; Xu, X.L.; Ouyang, H. Kin selection or resource partitioning for growing with siblings: Implications from measurements of nitrogen uptake. Plant Soil 2016, 398, 79–86. [Google Scholar] [CrossRef]
- Xu, Y.; Li, F.L.; Li, L.L.; Chen, X.; Meiners, S.J.; Kong, C.H. Discrimination of relatedness drives rice flowering and reproduction in cultivar mixtures. Plant Cell Environ. 2024, 47, 4572–4585. [Google Scholar] [CrossRef]
- Simonsen, A.K.; Chow, T.; Stinchcombe, J.R. Reduced plant competition among kin can be explained by Jensen’s inequality. Ecol. Evol. 2014, 4, 4454–4466. [Google Scholar] [CrossRef]
- Serra, N.S.; Shanmuganathan, R.; Becker, C. Allelopathy in rice: A story of momilactones, kin recognition, and weed management. J. Exp. Bot. 2021, 72, 4022–4037. [Google Scholar] [CrossRef]
- Anten, N.P.R.; Chen, B.J.W. Detect thy family: Mechanisms, ecology, and agricultural aspects of kin recognition in plants. Plant Cell Environ. 2021, 44, 1059–1071. [Google Scholar] [CrossRef] [PubMed]
- Bilas, R.D.; Bretman, A.; Bennett, T. Friends, neighbours and enemies: An overview of the communal and social biology of plants. Plant Cell Environ. 2021, 44, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Li, Z.; Li, F.L.; Xia, X.X.; Wang, P. Chemically mediated plant–plant interactions: Allelopathy and allelobiosis. Plants 2024, 13, 626. [Google Scholar] [CrossRef] [PubMed]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. Evol. Syst. 2024, 6, 207–215. [Google Scholar] [CrossRef]
- Homulle, Z.; George, T.S.; Karley, A.J. Root traits with team benefits: Understanding belowground interactions in intercropping systems. Plant Soil 2022, 471, 1–26. [Google Scholar] [CrossRef]
- Wang, C.Y.; Li, L.L.; Meiners, S.J.; Kong, C.H. Root placement patterns in allelopathic plant-plant interactions. New Phytol. 2023, 237, 563–575. [Google Scholar] [CrossRef]
- Belter, P.R.; Cahill, J.F., Jr. Disentangling root system responses to neighbours: Identification of novel root behavioural strategies. AoB Plants 2015, 7, plv059. [Google Scholar] [CrossRef]
- Biedrzycki, M.L.; Bais, H.P. Kin recognition in plants: Did we learn anything from roots? Front. Ecol. Evol. 2022, 9, 785019. [Google Scholar] [CrossRef]
- Monzeglio, U.; Stoll, P. Effects of spatial pattern and relatedness in an experimental plant community. Evol. Ecol. 2008, 22, 723–741. [Google Scholar] [CrossRef]
- Semchenko, M.; Saar, S.; Lepik, A. Intraspecific genetic diversity modulates plant-soil feedback and nutrient cycling. New Phytol. 2017, 216, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Sher, J.; Khan, N.; Tomlinson, K.W. Plant growth of Chenopodium quinoa (Willd) is better when growing with kin than with non-kin regardless of soil nutrient conditions. Plant Ecol. 2024, 225, 153–161. [Google Scholar] [CrossRef]
- Cahill, J.F., Jr.; McNickle, G.G.; Haag, J.J.; Lamb, E.G.; Nyanumba, S.M.; St. Clair, C.C. Plants integrate information about nutrients and neighbors. Science 2010, 328, 1657–1658. [Google Scholar] [CrossRef]
- Ulbrich, T.C.; Rivas-Ubach, A.; Tiemann, L.K.; Friesen, M.L.; Evans, S.E. Plant root exudates and rhizosphere bacterial communities shift with neighbor context. Soil Biol. Biochem. 2022, 172, 108753. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.M.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- File, A.L.; Klironomos, J.; Maherali, H.; Dudley, S.A. Plant kin recognition enhances abundance of symbiotic microbial partner. PLoS ONE 2012, 7, e45648. [Google Scholar] [CrossRef]
- Bolin, L.G.; Lau, J.A. Linking genetic diversity and species diversity through plant–soil feedback. Ecology 2022, 103, e3692. [Google Scholar] [CrossRef]
- Suárez-Moreno, Z.R.; Caballero-Mellado, J.; Coutinho, B.G.; Mendonça-Previato, L.; James, E.K.; Venturi, V. Common features of environmental and potentially beneficial plant-associated Burkholderia. Microb. Ecol. 2012, 63, 249–266. [Google Scholar] [CrossRef]
- Mukherjee, P.; Mitra, A.; Roy, M. Halomonas rhizobacteria of Avicennia marina of Indian sundarbans promote rice growth under saline and heavy metal stresses through exopolysaccharide production. Front. Microbiol. 2019, 10, 1207. [Google Scholar] [CrossRef]
- Wang, D.N.; He, X.M.; Baer, M.; Lami, K.; Yu, B.; Tassinari, A.; Salvi, S.; Schaaf, G.; Hochholdinger, F.; Yu, P. Lateral root enriched Massilia associated with plant flowering in maize. Microbiome 2024, 12, 124. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Sánchez, A.; Plasencia, J.; Monribot-Villanueva, J.L.; Rodríguez-Haas, B.; Ruíz-May, E.; Guerrero-Analco, J.A.; Sánchez-Rangel, D. Virulence factors of the genus Fusarium with targets in plants. Microbiol. Res. 2023, 277, 127506. [Google Scholar] [CrossRef] [PubMed]
- Munguia-Rosas, M.A.; Ollerton, J.; Parra-Tabla, V.; Arturo De-Nova, J. Meta-analysis of phenotypic selection on flowering phenology suggests that early flowering plants are favoured. Ecol. Lett. 2011, 14, 511–521. [Google Scholar] [CrossRef]
- Lu, T.; Ke, M.J.; Lavoie, M.; Jin, Y.J.; Fan, X.J.; Zhang, Z.Y.; Fu, Z.W.; Sun, L.W.; Gillings, M.; Penuelas, J.; et al. Rhizosphere microorganisms can influence the timing of plant flowering. Microbiome 2018, 6, 231. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Chen, X.H.; Hu, F.; Zhang, S.Z. Breeding of commercially acceptable allelopathic rice cultivars in China. Pest Manag. Sci. 2011, 67, 1100–1106. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellín, C. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2021, 67, 1–48. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Han, Q.-H.; Xie, S.-S.; Kong, C.-H. Neighbor Relatedness Contributes to Improvement in Grain Yields in Rice Cultivar Mixtures. Plants 2025, 14, 2385. https://doi.org/10.3390/plants14152385
Xu Y, Han Q-H, Xie S-S, Kong C-H. Neighbor Relatedness Contributes to Improvement in Grain Yields in Rice Cultivar Mixtures. Plants. 2025; 14(15):2385. https://doi.org/10.3390/plants14152385
Chicago/Turabian StyleXu, You, Qin-Hang Han, Shuai-Shuai Xie, and Chui-Hua Kong. 2025. "Neighbor Relatedness Contributes to Improvement in Grain Yields in Rice Cultivar Mixtures" Plants 14, no. 15: 2385. https://doi.org/10.3390/plants14152385
APA StyleXu, Y., Han, Q.-H., Xie, S.-S., & Kong, C.-H. (2025). Neighbor Relatedness Contributes to Improvement in Grain Yields in Rice Cultivar Mixtures. Plants, 14(15), 2385. https://doi.org/10.3390/plants14152385